
细鳞鲑对谷氨酸行为趋向的反应阈值
2019-09-03 07:57蔡红英陈春山张旭光郭弘艺
海洋渔业 2019年4期
蔡红英,魏 凯,陈春山,张旭光,郭弘艺,李 超,时 晓
(1.上海海洋大学水产与生命学院,上海 201306;2.北京市水生野生动植物救护中心,北京 102100;3.北京市鲟鱼鲑鳟鱼创新团队,北京 102100;4.中国水产科学研究院东海水产研究所,上海 200090)
细鳞鲑(Brachymystax lenok)是我国一种名贵的陆封型鲑科鱼类[1]。目前,由于多种人为原因,细鳞鲑的分布区域逐渐缩小,野生资源量急剧下降。为了保护和恢复细鳞鲑野生资源,研究人员开展了大量对其自然资源养护[2-4]、人工繁殖和养殖实验研究[5-6]。在鱼类人工养殖中,适口性是配合饲料能被鱼类迅速摄食和吸收的重要前提[7]。随着动物蛋白成本的提高,在鱼类配合饲料中增加植物蛋白成为一个必然的趋势[8]。然而,配合饲料中植物蛋白的比例增加可能会引起饲料的适口性下降,因此,需要在饲料中添加促进摄食的引诱物质来提高养殖鱼类的摄食动机以提高摄食效率[8]。鱼类摄食过程主要有3个阶段:感知食物、搜索食物和摄入食物[9]。氨基酸可以诱发鱼类摄食相关的嗅觉行为[10],被认为是最有效的鱼类诱食化合物之一[11]。有研究表明,单体谷氨酸和脯氨酸都可以有效地触发红大麻哈鱼(Oncorhynchus nerka)进行嗅觉印迹[12],说明鲑鱼对氨基酸的嗅觉趋向性可能是一种先天性的能力,也印证了氨基酸作为诱食剂的机制。细鳞鲑对多种氨基酸的行为反应研究表明,其对谷氨酸有较强的行为趋向性[3]。这种趋向性行为可以看作鱼类摄食过程中感知并趋向食物的阶段。但是细鳞鲑的行为正趋向的程度与氨基酸的浓度相关性仍不清楚。本文以细鳞鲑为研究对象,通过自制的Y形迷宫水槽研究了细鳞鲑对不同浓度谷氨酸的选择偏好性,以期获得细鳞鲑嗅觉行为敏感的最佳氨基酸浓度,为细鳞鲑诱食剂的开发利用提供实验参考。
1 材料与方法
1.1 实验材料
实验用鱼为取自吉林省延边细鳞鲑良种场当年孵化的细鳞鲑幼鱼,共计60尾,平均体质量为(250.0±2.4)g,平均体长为(24.0±1.8)cm。试验鱼置于实验室的帆布循环水槽(1m×Ф1.6 m)中暂养,水温由冷水机控制为(13±1)℃,正常自然光照,暂养期间每日投喂一次人工颗粒饵料,适量换水以保持水体清洁。实验时随机捞取健康的鱼进行实验。
1.2 实验方法
实验在自制的Y型迷宫水槽(76 cm×36 cm×20 cm)中进行,水深10 cm,水槽的一端设置长30 cm的档板将水槽一端分成2个相同的Y形选择臂槽,选择臂槽通过导管与刺激液水槽相连,通过调节阀控制刺激液以相同的流速进入选择臂槽(图1)。在水槽的另一端设置了出水口,并通过调节阀控制水流速度,以调节刺激液在选择臂的扩散速度。根据有色溶液扩散类比,20 mL溶液扩至选择臂末端的时间约为3 min,由此确定了视频记录时间为3min,以避免2个选择臂内刺激液混和。实验时,随机捞取1尾试验鱼到实验水槽,并由活动网隔离在选择臂前方适应15 min后移去活动网,同时打开氨基酸刺激液和对照组水溶液(与暂养用水同)的进水阀,并开始记录试验鱼的行为反应,视频文件保存在电脑中进行离线分析。谷氨酸刺激液分别配制浓度为1×10-2、1×10-3、1×10-4、1×10-5、1×10-6mol·L-1的5组母溶液。理论上,谷氨酸母溶液扩散至选择臂末端的最大实际浓度分别为3.7×10-5、3.7×10-6、3.7×10-7、3.7×10-8、3.7×10-9mol·L-1,浓度数量级为 1×10-5~1×10-9。实验时,不同浓度谷氨酸母溶液和对照组水溶液随机加入不同的刺激液槽内,以减少位置信息对实验结果的影响。每组实验分别测试了10尾细鳞鲑幼鱼对不同浓度谷氨酸的嗅觉行为反应。每次实验结束,试验鱼置于暂养水槽的隔离网笼内,并冲洗实验水槽,更换新水进行下一次实验。
1.3 数据分析
实验中,细鳞鲑幼鱼对不同浓度谷氨酸的趋向反应可表现3个阶段:首先是对味道信息的感知,主要表现出游向选择臂的动机,比如头部转向选择臂或胸鳍扇动速度增加;进而选择进入或远离选择臂槽;最后在选择臂槽停留或搜寻。依据试验鱼的行为反应,对试验鱼的初次行为反应进行定性评分[13]:实验开始后,试验鱼改变原先身体方向,游向选择臂或头部转向特定的选择臂,可看到胸鳍扇动速度增加,但并没有进入选择臂时,可视为1分;试验鱼头部或一半身体进入选择臂内时,则视为2分;试验鱼整个身体全部进入选择区视为3分。在这种打分评价试验鱼选择性时,只对试验鱼首次的选择结果进行评分,而没有被选择的组则为0分。通过比较试验鱼对不同刺激液的选择总分值,可以快速、定性地确定试验鱼对不同浓度谷氨酸的趋向偏好差异。但是这种评价并不能反映试验鱼在选择臂的停留时间和反复进入次数等,而且,试验鱼也会改变选择,而趋向相反的选择臂[14]。因此,引入偏好度(preference degree)作为试验鱼趋向程度大小的指标[3]。试验鱼对不同浓度谷氨酸刺激液的偏好度定义为P=(n×T)/t。式中,P为偏好度;n为进入选择臂总次数;T为在选择臂的平均停留时间;t为第一次进入选择臂的反应时间或延迟时间。这样,试验鱼进入选择臂的次数越多,停留时间越长或第一次进入反应越短,则说明其对刺激液的偏好程度越大。为了更直观比较试验鱼对不同浓度谷氨酸的趋向性,将每组实验中试验鱼对水溶液的偏好度定义为1,根据公式R=PGlu/Pwater进行标准化转化,计算试验鱼对不同浓度谷氨酸的相对偏好度(R),通过比较相对偏好度大小来确定最佳趋向反应的谷氨酸浓度,并根据试验鱼对该浓度的第一次平均反应时间确定其嗅觉反应阈值。所有数据均表示为平均值 ±标准误(Mean±SEM),由 Sigmaplot(v 12.5)进行绘图,并进行显著性分析(T检验)。
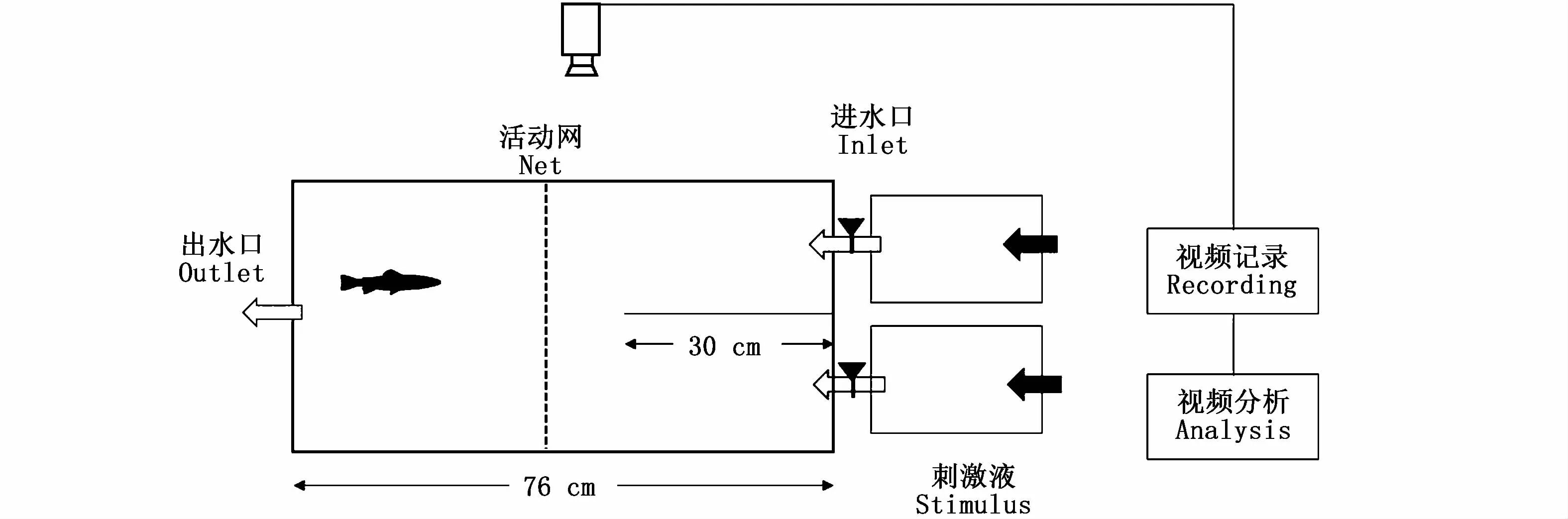
图1 Y型迷宫水槽示意图Fig.1 Schematic diagram of the Y maze flow system
2 结果与分析
2.1 行为表现方式
实验分别测试了细鳞鲑幼鱼对5组不同浓度的谷氨酸刺激液的行为反应。试验鱼对不同浓度谷氨酸有不同的反应。移去挡板后,试验鱼通常会表现缓慢的游泳状态或原地摆动尾巴。给予高浓度谷氨酸刺激液时(1×10-5mol·L-1),试验鱼(n=4)有时会表现出快速逃离行为,游到远离选择臂的角落,一直至实验记录结束;或表现出急剧的冲撞水槽壁的行为(n=2);个别试验鱼(n=2)会尝试再次游向谷氨酸选择臂,但同样会快速的逃离,这可能与氨基酸刺激液在水中的不均匀分散有关;或直接进入对照组选择臂(n=3)。当给予低浓度的谷氨酸时(1×10-8mol·L-1),试验鱼(n=5)通常选择进入选择臂停留或小范围的转圈游动,有时离开选择臂绕一圈后再次返回进入有谷氨酸的选择臂中或进入对照组侧(图2)。
2.2 嗅觉反应阈值
细鳞鲑幼鱼对5组不同浓度氨基酸刺激液的行为反应得分总值如图3。在浓度1×10-5mol·L-1谷氨酸刺激液与水的试验组(以下简称为谷氨酸和水)中,试验鱼对谷氨酸刺激液反应的平均得分为(0.90±0.27),低于对水溶液的反应平均得分(1.20±0.48)(P<0.05),其中 4尾鱼对水溶液的反应得分为3分,没有试验鱼对氨基酸的反应得分为3分;在1×10-8mol·L-1谷氨酸和水中,试验鱼对谷氨酸的反应平均得分(1.56±0.48),高于对水溶液的平均得分(0.67±0.27)(P<0.05),其中有 4尾鱼对氨基酸刺激的反应得分为3分;在1×10-6mol·L-1谷氨酸和水及1×10-7mol·L-1谷氨酸和水中,试验鱼对水反应的平均得分分别为(1.22±0.41)和(1.33±0.42),大于对谷氨酸的反应平均得分(0.67±0.35)和(0.89±0.43),但二者没有显著性差异(P>0.05);在1×10-9mol·L-1谷氨酸和水中,试验鱼对谷氨酸的平均得分为(1.11±0.33),大于对水反应的平均得分(0.89±0.43),二者没有显著性差异(P>0.05)。在1×10-6、1×10-7、1×10-8mol·L-1谷氨酸的试验组中,试验鱼对谷氨酸和水溶液的反应得分都有3分的情况。

图2 细鳞鲑幼鱼对谷氨酸刺激的行为反应模式Fig.2 Behavioral response to am ino acid stimulation in Brachymystax lenok juveniles
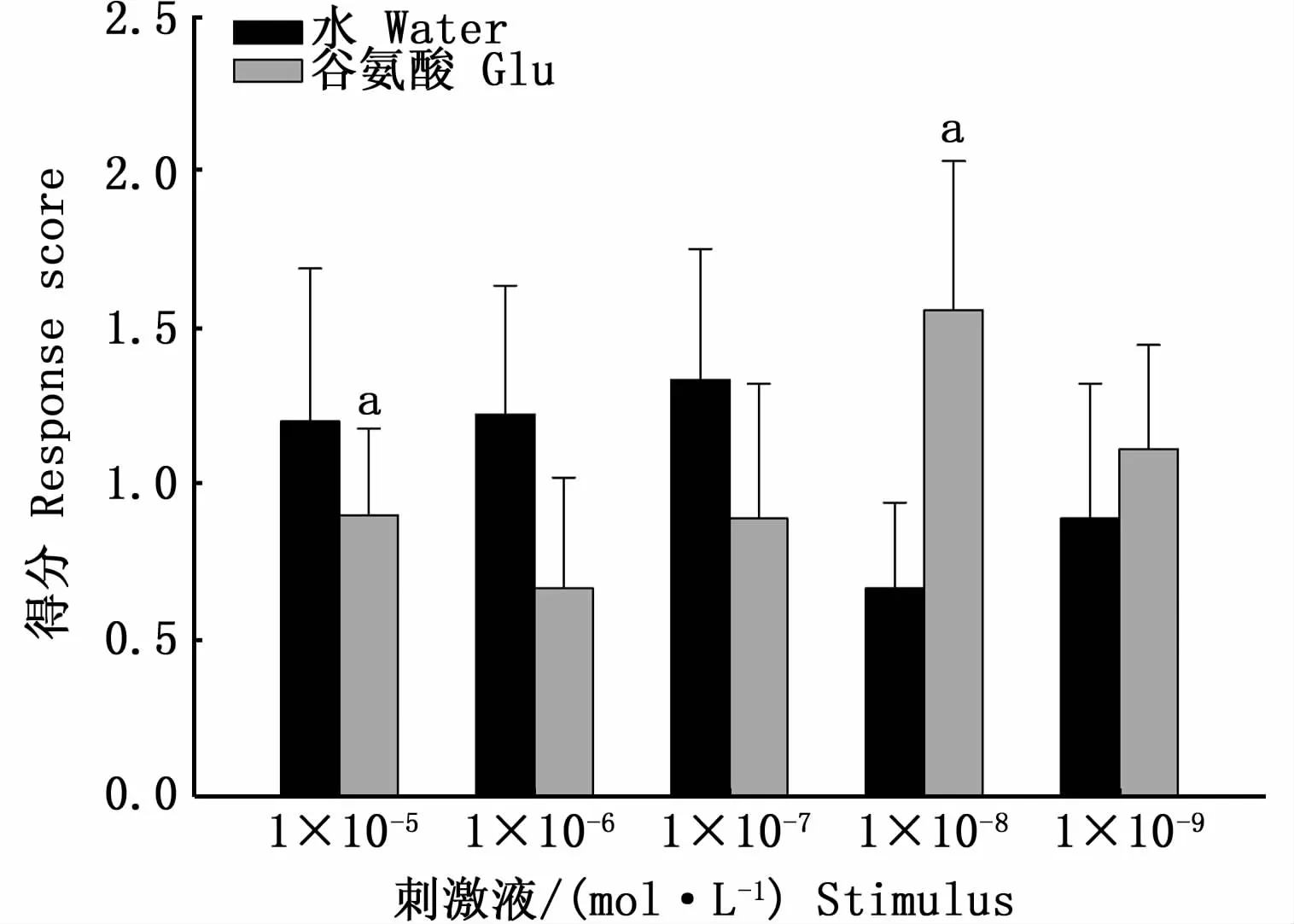
图3 细鳞鲑幼鱼对刺激液的行为反应得分Fig.3 Response scores of B.lenok juveniles to stim ulus
细鳞鲑幼鱼对5组不同浓度氨基酸刺激液的偏好度如图4所示。在1×10-5mol·L-1谷氨酸和水中,试验鱼对1×10-5mol·L-1谷氨酸的偏好度为(0.61±0.23),小于对水的偏好度(1.54±0.93),但二者没有显著性差异(P>0.05);在1×10-6mol·L-1谷氨酸和水中,试验鱼对 1×10-6mol·L-1谷氨酸的偏好度为(0.57±0.18),小于对水的偏好度(1.32±0.52),二者没有显著性差异(P>0.05);在1×10-7mol·L-1谷氨酸和水中,试验鱼对1×10-7mol·L-1谷氨酸的偏好度为(0.38±0.19),小于对水的偏好度(1.01±0.45),二者没有显著性差异(P>0.05);在1×10-8mol·L-1谷氨酸和水中,试验鱼对1×10-8mol·L-1谷氨酸的偏好度为(2.93±0.89),大于对水的偏好度(0.77±0.36),二者有显著性差异(P<0.05);在1×10-9mol·L-1谷氨酸和水中,试验鱼对 1×10-9mol·L-1谷氨酸的偏好度为(1.38±0.81),略大于对水的偏好度(1.20±0.39),二者没有显著性差异(P>0.05)。
根据标准化转换公式,试验鱼对3.7×10-5~3.7×10-9mol·L-1谷氨酸刺激的相对偏好度分别为 0.396、0.436、0.381、3.801、1.142。根据试验鱼对谷氨酸刺激液的行为反应得分和相对偏好度,试验鱼对实际浓度为3.7×10-8mol·L-1谷氨酸刺激液的行为趋向最强。试验鱼对谷氨酸刺激液的反应时间为 18—57 s,平均值为(34.5±9.4)s,根据溶液扩散,在34.5 s时的选择臂槽内的谷氨酸实际浓度约为0.7×10-8mol·L-1,即细鳞鲑对谷氨酸行为趋向的反应阈值为0.7×10-8mol·L-1。

图4 细鳞鲑幼鱼对氨基酸的偏好度Fig.4 Preference degree to am ino acids in B.lenok juveniles
3 讨论
目前,鱼类配合饲料中诱食剂的研究一直是一个备受关注的研究课题。外源性氨基酸类物质不仅可以满足鱼类的营养需求,而且可以诱发鱼类摄食动机,促进鱼类的摄食进程和饲料利用率[11]。如鳗鲡科鱼类(Anguillidae)配合饲料行业标准(SC/T 1004-2004)就明确规定了蛋氨酸的添加指标[15]。但非必需氨基酸由于动物可以自身合成,在诱食剂的研究中往往被忽略,比如谷氨酸。在氨基酸类诱食剂研究中,常根据鱼类喜食的天然饵料中主要成份来锁定可能的氨基酸种类[16]。细鳞鲑是典型肉食性鱼类,在自然条件下,幼鱼期喜食水蚤、摇纹幼虫等小型生物以及各种落入水中的昆虫等,成年期也能摄食自己体长1/2的鱼类[1]。对其天然饵料中的氨基酸成分分析发现,谷氨酸、丙氨酸、精氨酸、甘氨酸是主要的氨基酸[17]。细鳞鲑肌肉中的谷氨酸(1.75%,干重)、天冬氨酸(1.59%)、甘氨酸(0.85%)和丙氨酸(0.75%)也是鱼体含量较高的几类氨基酸[18]。对细鳞鲑栖息河流的主要氨基酸种类调查发现,按含量顺序分别为谷氨酸、天冬氨酸、丝氨酸、甘氨酸、组氨酸、丙氨酸等,其中谷氨酸占总量的20%以上[19]。谷氨酸不仅可以对马苏大麻哈鱼(Oncorhynchusmasou)形成嗅觉印迹[13],而且可以激活未开口的大西洋鲑(Salmo salar)仔鱼的猛冲、咬合等一系列摄食行为[10]。结合细鳞鲑对氨基酸趋向偏好性的研究结果,表明谷氨酸也可能成为潜在的细鳞鲑诱食添加剂。
本文通过Y形迷宫实验研究了细鳞鲑幼鱼对谷氨酸的行为趋向,同时通过选择偏好度[3]和行为特征评分[13]确定其最敏感的氨基酸浓度。实验结果表明,细鳞鲑幼鱼对1×10-5、1×10-6、1×10-7mol·L-1谷氨酸刺激液的相对偏好度小于1,表明其更偏向水溶液,对较高浓度的谷氨酸刺激具有回避或负趋向性行为;而对1×10-8、1×10-9mol·L-1谷氨酸母溶液刺激的相对偏好度大于1,表明其更偏向谷氨酸溶液,对较低浓度的谷氨酸刺激具有趋向性行为。行为评分的分值也同样体现类似的趋向,对1×10-8、1×10-9mol·L-1谷氨酸母溶液刺激的行为分值也高于水溶液。在施氏鲟(Acipenser schrenckii)对几种氨基酸的诱食活性研究中有类似的结果,1×10-3、1×10-4mol·L-1的苏氨酸、精氨酸母溶液刺激对施氏鲟的摄食行为有明显的抑制性,而母溶液浓度降至1×10-5mol·L-1时,则产生明显的诱食活性[16]。这也表明高浓度氨基酸浓度对鱼类产生抑制行为,而低浓度的氨基酸对鱼类具有诱食性。因此,确定鱼类对氨基酸的行为反应阈值浓度是研究诱食剂的重要前提之一。在本实验中,细鳞鲑幼鱼对实际浓度小于3.7×10-8mol·L-1的谷氨酸刺激表现出正向的行为趋向反应。魏凯等[3]研究了细鳞鲑幼鱼对几种氨基酸的嗅觉行为反应,谷氨酸实际浓度为2.5×10-6mol·L-1时引起试验鱼明显的偏好选择。在本实验中,试验鱼对实际浓度 3.7×10-6mol·L-1的谷氨酸并没有明显的趋向性,且相对偏好度依然小于1。这种看似矛盾的结果,原因其一可能是实验中对照组选择的差异造成,魏凯等[3]采用甘氨酸作为对照组,以确定试验鱼对不同氨基酸的选择差异,而本实验中以水溶液作为对照组;其次,本实验所用的试验鱼个体较大,可能不同生长阶段的鱼对谷氨酸的反应阈值存在差异。SHPARKOVISKY等[20]研究也发现,随着年龄的增长,大西洋鲑对亮氨酸的正趋向性反应增加,对缬氨酸和丙氨酸的正趋向性反应减少,而精氨酸和组氨酸引起的负趋向性反应随着鱼的年龄增长也变得更加明显。但鱼类这种趋向行为的变化是否与其嗅觉敏感性相关,仍有待进一步的实验研究。
尽管很多研究证实单体的氨基酸可以诱发鱼类的摄食动机,但也有研究表明单体氨基酸的诱食效果并不好。王常安等[21]在氨基酸对哲罗鲑(Hucho taimen)的诱食研究中发现,饲料中添加0.5%的谷氨酸、丙氨酸、甘氨酸、脯氨酸对哲罗鲑的诱食效果不明显。这可能是由于添加的氨基酸的剂量不合适,如1×10-3mol·L-1谷氨酸对施氏鲟摄食有明显抑制性,而1×10-4、1×10-5mol·L-1谷氨酸对其摄食行为的抑制和引诱效果都不显著。另一方面也说明,相同的氨基酸对不同鱼类的作用存在差异。如脯氨酸和丙氨酸可以诱发褐鳟(Salmo trutta)幼鱼的摄食行为[10],却是虹鳟(S.garidneri)的抑食物质[22]。因此,在诱食剂研究中,需要首先通过行为学和电生理方法,筛选出对鱼类具有明显引诱作用的促摄食物质,并确定其反应阈值。本文虽然通过行为学的方法确定了细鳞鲑对谷氨酸的趋向行为的反应阈值,但还需要经过长期的摄食行为和生长实验来精确验证。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
特种经济动植物(2022年2期)2022-02-15
中国饲料(2021年17期)2021-11-02
小资CHIC!ELEGANCE(2021年28期)2021-08-18
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
西夏学(2020年2期)2020-01-24
当代陕西(2019年8期)2019-05-09
分析化学(2019年3期)2019-03-30
复旦学报(医学版)(2016年6期)2016-12-20
