
气孔运动中保卫细胞壁作用的研究进展
2019-10-14 02:44魏建华戴绍军王宏芝
生物技术进展 2019年5期
周 影, 郑 林, 魏建华, 马 艳, 戴绍军, 王宏芝*
1.东北林业大学盐碱地生物资源环境研究中心, 东北油田盐碱植被恢复与重建教育部重点实验室, 哈尔滨 150000;2.北京市农林科学院北京农业生物技术研究中心, 北京 100097;3.江苏金陵科技学院, 南京 210038
气孔是植物叶表皮组织上的小孔,是由两个保卫细胞围绕而成的孔隙。气孔是植物与外界环境气体交换的门户,控制水分蒸腾和CO2吸入,在植物生长发育中发挥重要作用。气孔运动是植物重要生命活动之一,受环境和保卫细胞内部信号转导以及细胞膨压控制,通过细胞大小和形状的改变来实现。红光和蓝光、高湿和低浓度CO2等条件能够诱导保卫细胞膨压升高,细胞内溶质快速增加后吸水膨胀、体积增加、形状改变、气孔开放;反之,黑暗、高浓度CO2、脱落酸等能够使保卫细胞膨压下降、失水收缩、气孔关闭。在气孔开放时,保卫细胞的体积可增加40%,细胞内部压力可达5 MPa, 相当于大气压的50倍[1]。保卫细胞形状改变程度与细胞壁的机械性能密切相关,气孔正常运动要求保卫细胞的细胞壁不但能承受很高的物理张力,同时具有良好的柔韧性和弹性。
保卫细胞壁属于初生细胞壁,主要由纤维素、木葡聚糖(一种半纤维素)、果胶和少量结构蛋白组成。根据初生细胞壁模型[2]推测,保卫细胞中纤维素和木葡聚糖组成的网络结构以及填充其中的果胶多糖基质,是维持保卫细胞壁弹性与强度平衡的基础。随着细胞壁相关分子遗传学、生物化学研究的不断深入,以及分析模型的引入,人们已开始解析保卫细胞壁的各组成成分及空间分布特性对细胞壁机械性能和气孔运动功能的影响。本文将讨论这一研究领域在分子遗传学和生物化学研究中的最新成果,重点讨论保卫细胞壁不均一特性的最新进展,包括其在气孔开放运动中的重要作用、进化上的保守性和发育形成过程;并将分别论述保卫细胞壁中纤维素、木葡聚糖、果胶、木质素和细胞壁松弛因子等组分在维持气孔保卫细胞功能中的作用,同时展望了保卫细胞壁合成及修饰相关分子机制研究的发展方向,以期为保卫细胞壁的遗传改良提供思路。
1 保卫细胞壁结构组成特性
1.1 保卫细胞壁不均一特性在气孔运动中的作用
保卫细胞在发育过程中细胞壁沉积和修饰呈非对称性,形成不均一细胞壁。保卫细胞壁的位置特异性加厚,能够满足气孔开放和关闭对细胞壁不同部位强度、刚性和弹性的差异要求,是保卫细胞不均匀膨胀和气孔运动中形状改变的主要决定因素。肾型保卫细胞内壁(腹壁)加厚程度大于外壁(背壁),富含结晶纤维素,导致保卫细胞内壁的刚性大于外壁,而外壁的弹性大于内壁;哑铃型保卫细胞的球状末端壁较薄而中间棒状部分细胞壁较厚。传统的观点认为,保卫细胞外壁和内壁厚度和刚性的差异,使保卫细胞在膨压作用下外壁弯曲程度大于内壁,导致气孔开放[3]。但Carter等[4]利用原子力显微镜结合模型分析的最新研究表明,保卫细胞端壁特异加厚在气孔开放运动中起关键作用。保卫细胞内壁和外壁厚度和刚性的差异只存在于成熟的保卫细胞中。未成熟保卫细胞虽然没有这种差异,但同样能够响应环境和细胞内部信号而正常开放。进一步研究表明,保卫细胞端壁处的特异性加厚在不同物种中广泛存在,并且这种加厚在保卫细胞母细胞发育阶段即已发生,导致此处细胞壁刚性增强。在气孔开放运动过程中加厚的端壁限制了保卫细胞复合体的纵向伸长,而改为横向膨大,保证了气孔的有效开放。
1.2 保卫细胞不均一细胞壁的进化保守性
尽管从蕨类植物到被子植物的演化过程中,保卫细胞壁组分发生了显著变化[5],但细胞壁强度和弹性的不均一分布特性,特别是端壁的特异加厚特性未发生明显改变[4,6]。蕨类植物中,保卫细胞端壁富含木质素,以增加端壁细胞壁强度; 在被子植物中,端壁中结晶纤维素代替了木质素,此处细胞壁增厚[5];水生被子植物的基部,由于气孔一直保持开放状态,属于无功能状态的气孔,端壁的纤维结晶度较低[7];而陆生被子植物,除了结晶纤维素外,去甲酯化果胶的参入,进一步提高了端壁强度和刚性[8]。Shtein等[5]考察了6种不同种类植物的3种保卫细胞壁组分,包括蕨类植物的肾型保卫细胞、双子叶植物的肾型保卫细胞和单子叶植物的哑铃型保卫细胞,发现纤维结晶度存在3种不同类型。其中双子叶植物的肾型保卫细胞端壁的纤维结晶度最高,数字化生物机械模式分析显示此区域是气孔运动过程中承受张力最大的区域。保卫细胞端壁组分在不同物种之间的差异,推测是进化过程中不同环境选择压力的结果[5]。但这种差异并不影响保卫细胞壁强度和弹性的不均一分布特性以及气孔功能,二者在进化中均十分保守。
1.3 保卫细胞不均一细胞壁的发育形成过程
保卫细胞壁不均一加厚,自保卫细胞母细胞分裂前即已开始。以双子叶植物肾型保卫细胞发育为例,细胞壁经过一系列非对称沉积和修饰,最终发育成具有功能的保卫细胞(图1)。首先,在对称分裂发生前,保卫细胞母细胞端壁加厚,之后细胞以垂直于加厚端壁的方向发生分裂(图1A)[9]。新形成的细胞壁富含胼胝质和果胶,附近聚集微管束。随后胼胝质和果胶降解(图1B),两个保卫细胞内壁分离,形成孔状结构(图1C和1D)。胼胝质合成和降解[10]以及果胶降解[11]的精细时空特异性调控对正常孔状结构形成具有重要作用,尽管其中分子调控机制尚不清楚。成熟保卫细胞位于孔状结构处的内壁主要由结晶纤维素组成,细胞壁明显增厚,弹性下降,强度和刚性增加。同时,极性端壁及两个保卫细胞连接处细胞壁亦明显增厚,富含结晶纤维素和去甲酯化果胶,细胞壁强度和刚性明显提高(图1D)[4]。
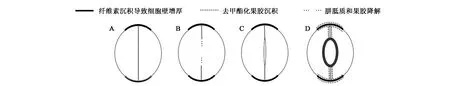
图1 保卫细胞壁分化过程中形成非对称性细胞壁[8]Fig.1 The asymmetric accumulation and modification of cell walls in stomatal differentiation[8].A:保卫细胞母细胞端壁出现纤维素加厚,细胞以垂直于加厚端壁方向发生对称分裂;B:新生细胞壁在特定部位发生胼胝质和果胶降解;C:两个保卫细胞内壁分离,形成孔状结构;D:成熟保卫细胞,内壁大量沉积结晶纤维素,细胞壁较外壁明显增厚;两个细胞连接处细胞壁及端壁亦明显增厚,富含去甲酯化果胶。
2 保卫细胞壁组分对气孔运动功能的影响
随着植物基因组计划的广泛开展和基因功能研究的深入,细胞壁组分影响保卫细胞壁机械性能和气孔运动的分子遗传学证据不断涌现。这些研究成果为保卫细胞壁的分子遗传改良提供了可能性。
2.1 纤维素和木葡聚糖对气孔运动功能的影响
Rui等[12,13]通过图像分析结合计算机械模型,对拟南芥纤维素和木葡聚糖突变体进行详细分析,从遗传学角度证明了纤维素和木葡聚糖与保卫细胞壁机械特性及气孔功能密切相关。拟南芥纤维素合成酶CesA3的突变体分析表明,保卫细胞壁纤维素含量与气孔开度密切相关。纤维素含量降低导致气孔响应外界条件信号的开关功能异常,气孔开度增加;突变体互补实验显示,当突变体纤维素含量恢复正常后,气孔开度亦恢复正常。木葡聚糖缺失突变体(xxt1xxt2),由于木葡聚糖含量降低,气孔开度减小,叶片表面温度升高。分析表明,木葡聚糖通过调节气孔纵向的收缩和扩张,调节气孔大小。纤维素和木葡聚糖参与气孔运动还体现在气孔运动过程中纤维素微纤丝发生的重新排列。气孔开放时纤维素微纤丝相对均匀地分布在细胞壁上,气孔关闭时,微纤丝成束存在。气孔运动过程中纤维素微纤丝的这种重新排列,即细胞壁结构改变需要木葡聚糖协助完成。木葡聚糖可以直接和纤维素结晶区交联,增加细胞壁的承重和抗张性[14]。尽管保卫细胞壁中纤维素和木葡聚糖缺失对气孔开度影响结果不同,但研究者认为纤维素和木葡聚糖缺失均会导致细胞壁强度和刚性增强[15]。
2.2 果胶对气孔运动功能的影响
保卫细胞壁中富含果胶,生物化学相关研究结果表明细胞壁果胶与气孔功能密切相关。早在1985年,Wu等[16]推测细胞壁中果胶基质多糖参与保卫细胞壁径向延伸。苔藓植物作为最早出现气孔的植物,气孔只在孢蒴膨大过程的短暂生长期能够开关,其后即失去开关能力。胶体金免疫标记技术研究表明,苔藓植物保卫细胞发育过程中细胞壁中果胶含量呈下降趋势,推测果胶含量下降与气孔失去运动能力有关[17]。Jones和Milne[18]使用聚阿拉伯糖内切酶处理植物表皮,解离聚鼠李糖半乳糖醛酸Ⅰ型果胶的阿拉伯糖侧链之后,使用壳梭包杆菌素诱导气孔开放或者脱落酸诱导气孔关闭,发现相应的气孔运动受到限制。进一步分析表明果胶中阿拉伯聚糖侧链决定保卫细胞壁弹性,进而影响气孔开关。
果胶甲酯化程度影响气孔功能。果胶多糖在高尔基体中合成,之后以高度甲酯化形式分泌到胞外。在细胞壁上沉积后,由果胶甲酯酶(pectin methylestrases,PME)选择性切除甲基基团。推测果胶去甲酯过程具有时空特异性。有关果胶甲酯化程度影响保卫细胞壁机械性能和气孔运动的最早的遗传学证据来源于Amsbury等[19]的研究成果。拟南芥果胶甲酯酶基因PME6突变后,细胞壁中去甲酯化果胶含量下降,气孔开放及关闭潜力下降,导致在高浓度CO2环境中,突变体气孔关闭不严,而在低浓度CO2环境中,气孔开放程度小于对照。推测保卫细胞中去甲酯化果胶含量减少导致细胞壁弹性下降。Huang等[20]利用拟南芥突变体证明果胶甲酯酶基因PME34参与保卫细胞热激响应。正常条件下,植物能够响应热激胁迫,改变气孔开度和蒸腾效率,以降低叶片表面温度和调节水分利用效率。PME34基因突变后,拟南芥响应热激能力丧失。此外,研究表明多聚半乳糖醛酸酶(一种果胶水解酶)参与气孔运动[11],而果胶甲酯酶可以通过提高果胶多聚半乳糖醛酸酶活性,进一步缓解胁迫对细胞壁造成的伤害[21]。推测果胶甲酯酶和多聚半乳糖醛酸酶在气孔运动过程中可以调节果胶分子的动态变化,其活性受植物生长发育和环境信号严格调控。
2.3 木质素在气孔运动中的作用
虽然木质素主要存在于次生细胞壁中,但保卫细胞壁中仍可检测到少量木质素。木质素是否与气孔功能有关,目前尚不清楚。蕨类植物保卫细胞端壁富含木质素,以增加细胞壁强度和刚性;被子植物只在保卫细胞内壁中能够检测到少量木质素[5]。拟南芥R2R3-MYB转录因子AtMYB61[22]及其杨树同源基因PtoMYB170[23]参与木质素合成调控。过量表达AtMYB61和PtoMYB170导致木质素沉积增强,次生细胞壁增厚。除了在维管组织中高水平表达外,这两个基因均在保卫细胞中高水平表达,过量表达导致气孔开度减小,而突变体气孔开度增加。拟南芥突变体中详细分析显示AtMYB61参与黑暗诱导的气孔关闭,而且这一过程独立于ABA和空气湿度诱导的气孔关闭途径。推测AtMYB61表达水平变化影响了保卫细胞壁特性,进而影响了气孔开度[22]。由于AtMYB61参与果胶沉积调控[24],所以AtMYB61突变体气孔开度改变是否与保卫细胞壁木质素沉积改变有关尚有待进一步研究。
2.4 细胞壁松弛因子对气孔运动功能的影响
气孔开放和关闭运动过程中,细胞壁纤维素微纤丝发生重新排列[13]。推测这一过程中纤维素微纤丝与交联的木葡聚糖间的氢键发生断裂,木葡聚糖分子断裂并重新连接,细胞壁松弛,在此基础上纤维素微纤丝发生滑动,促进保卫细胞在膨压作用下膨胀和收缩。已报告的细胞壁松弛因子主要包括膨胀素(expansin)蛋白、木葡聚糖内转糖苷酶/水解酶(xyloglucan endotransglucosylase (XET)/hydrolase,XTH)、糖基水解酶(glycosidehydrolase, GH)和羟基自由基(hydroxyl radical,·OH)[25]。目前有关细胞壁松弛因子参与气孔运动的研究有待深入,只有少数有关膨胀素蛋白、木葡聚糖内转糖苷酶/水解酶参与气孔运动的初步遗传学报道。
2.4.1膨胀素蛋白与气孔运动功能 膨胀素蛋白,也称扩展蛋白,是一类能够使细胞壁松弛的细胞壁活性蛋白[26]。膨胀素蛋白通过一种非酶机制,打断纤维素微纤丝和细胞壁基质多糖的非共价连接,导致植物细胞壁中纤维素微纤丝滑动,促进细胞壁在膨压作用下蠕动[27]。拟南芥细胞壁膨胀素蛋白EXPA1在保卫细胞特异表达,过量表达增加转基因株系在光诱导条件下的气孔开度;抑制该蛋白活性,降低了气孔对光刺激的敏感性[28]。推测细胞壁膨胀素蛋白通过改变保卫细胞壁的松弛性来调节气孔开放。
2.4.2木葡聚糖内转糖苷酶/水解酶与气孔运动功能 木葡聚糖内转糖苷酶/水解酶(xyloglucan endotransglucosylase/hydrolase, XTH)是植物细胞壁结构改变的关键酶,松弛细胞壁能够催化木葡聚糖分子水解与再连接,通过一种“分子嫁接”的机制介导细胞壁的结构改变和松弛[29]。研究者在西红柿中过量表达辣椒XTH3基因,干旱胁迫下转基因西红柿气孔关闭比例增加,干旱忍受能力增强。推测气孔关闭与XTH3基因过量表达引起的细胞壁结构改变有关[30]。
3 展望
气孔是植物与外界环境气体交换的门户,对全球碳和水分循环具有重要影响。气孔开放是植物的重要生命活动之一,与植物水利用效率和农业生产效率密切相关[31]。气孔在降低叶片温度和代谢流以及信号长距离运输转导中发挥重要作用[32~34]。有效调控气孔开关,提高植物光合效率和水资源利用效率,在全球气候变暖和水资源危机干旱持续发生的今天显得尤为重要。深入解析保卫细胞壁影响气孔运动的分子遗传机制,将为保卫细胞壁遗传改良和创制耐干旱植物提供新的策略。
尽管大量的遗传学研究已表明保卫细胞壁组成和空间分布特性影响气孔功能,但调控保卫细胞壁合成、修饰和特异沉积的上游转录调控因子和信号转导途径,以及这一过程对环境的响应知之甚少。转录调节在植物生长发育和响应环境信号过程中发挥重要作用。次生细胞壁合成调控研究表明,转录调控是次生细胞壁合成调控的核心,由众多转录因子共同参与,形成复杂的分级调控网络,协同调控细胞壁各组分的合成。因此,挖掘保卫细胞壁合成调控关键转录因子,将有助于阐明保卫细胞壁参与气孔运动调节和环境响应的分子机制及调控网络。由于气孔的开放和关闭,明显影响叶片表面温度,而叶片表面温度可以通过红外相机鉴定。利用这种方法,可以快速鉴定气孔突变体,挖掘影响气孔功能的转录因子。此外,随着植物转录组和基因组计划的广泛开展,根据基因特异表达模式,利用反向遗传学方法,同样可以挖掘潜在的候选基因。
有关保卫细胞壁参与气孔运动的另一个基本问题是:在气孔运动过程中,保卫细胞壁基质多糖(木葡聚糖和果胶多糖)的动态变化和重构是如何进行的?尽管我们对气孔运动过程中保卫细胞壁动态变化知之甚少,但现有的知识提示我们细胞壁果胶分子修饰相关酶类及细胞壁松弛因子可能参与气孔运动过程中保卫细胞壁的重构。可以利用反向遗传学方法,对保卫细胞中特异表达的果胶甲酯酶、果胶乙酰化酶、膨胀素蛋白、木葡聚糖内转糖苷酶/水解酶等基因进行功能鉴定。
猜你喜欢
乡村科技(2021年12期)2021-09-06
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
三农资讯半月报(2020年18期)2020-10-14
中国糖料(2019年1期)2019-02-13
数码世界(2018年1期)2018-12-23
中国酿造(2018年10期)2018-11-03
中国野生植物资源(2018年3期)2018-08-06
中成药(2018年6期)2018-07-11
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
