
mGluR6对大鼠胚胎神经干细胞生物学功能的影响*
2020-04-10 16:45王玉珏辛宁宁
陕西医学杂志 2020年3期
王玉珏,辛宁宁
1.延安大学医学院医学影像学专业(延安716000);2.咸阳师范学院校医院(咸阳712000)
神经系统疾病是危害人类健康重大疾病之一,神经损伤后的治疗和康复是医学界的重要问题[1-2]。干细胞的发现,改变了以往认为成年哺乳动物中枢神经系统的神经细胞不能再生的观念,这促使干细胞成为中枢神经系统疾病治疗新策略而备受关注。神经干细胞(NSCs)是一种具有多向分化潜能和自我更新能力的细胞,在特定的条件下可以分化形成神经元、星形胶质细胞和少突胶质细胞。许多因素例如神经递质、神经营养因子、化学引诱物等参与调控NSCs的分化与增殖。谷氨酸是NSCs中最关键的性神经递质,它通过与相应的受体分子作用,参与中枢神经系统正常的生理功能[3]。根据药理学和电生理学的特征,谷氨酸受体分为两种:即代谢型受体(Metabotropic glutamate receptors,mGluRs)和离子型受体(Ionotropic glitamate receptors,iGluRs)。iGluRs按其特异靶向激动剂的不同,又分为α-氨基-3-羟基-5-甲基-4-异恶唑丙酸(AMPA)、N-甲基-D-门冬氨酸(NMDA)及红藻氨酸(KA)三类受体;mGluRs属于G-蛋白偶联受体家族,按其氨基酸序列、激动剂选择性和信号传导机制的差异分为三组,分别为组I(mGluR1,mGluR5)、组II(mGluR2-3)和组III(mGluR4,mGluR6-8[4]。作为G蛋白耦联受体家族,mGluRs的生物学功能主要通过激活胞内第二信使通路从而引起生物学效应[5]。
自1993年研究者将mGluR6从大鼠视网膜的cDNA文库中分离出来[6],Masu等人发现 mGluR6主要存在于视网膜细胞中,并在视网膜双极细胞的突触传递中发挥重要作用[7]。Foreman等人通过Western blot方法在大鼠骨髓基质细胞中检测到mGluR6的表达,这也是研究人员首次在除视网膜外的其它细胞中检测到mGluR6的表达,他们的研究结果表明mGluR6通过抑制Ca2+内流,抑制一氧化氮(NO)的产生[8]。随后,人们在前列腺癌细胞和小神经胶质细胞中检测到mGluR6的表达[9]。值得一提的是,Vardi等人通过转基因技术在mGluR6启动子区域构建GFP报告基因,在角膜内皮、睾丸、肾髓质、肾小球、以及B淋巴细胞等非神经组织细胞中检测到mGluR6的表达[10]。该研究结果拓宽了人们对mGluR6调控细胞发育的认识,说明mGluR6并不局限于视网膜细胞中发挥作用,但其相应的调控机制至今未见报道。最近Zheng等在人缘表皮角化细胞系HaCaT中研究表明,敲除mGluR6导致细胞骨架重组,进一步研究发现mGluR6可能通过CaM KII/ERKMLC信号通路调节角质细胞的吞噬作用[11],截至目前,mGluR6是否参与NSCs生长及其调控机制还鲜见相关报道。在这个研究中,我们旨在探讨mGluR6对大鼠胚胎神经干细胞生物学功能的影响。
材料和方法
1 实验动物 Sprague-Dawley(SD)大鼠由西安交通大学医学部实验动物中心提供。本研究中使用的实验动物的饲养严格遵循中华人民共和国卫生部令第55号发布的《医学实验动物管理实施细则》和国家科学技术委员会第2号令发布的《实验动物管理条例》。孕鼠受孕后15d,对胚胎鼠进行分离,然后对大鼠胚胎NPCs进行培养。研究得到了延安大学医学伦理委员会的同意。
2 实验试剂 DMEM/F12(1∶1)培养基,N2、B27添加剂,bFGF购自Gibco公司(美国);EGF、肝素、MTT、RnaseA、胰蛋白酶、碘化丙啶(PI)购自Sigma公司(美国);AnnexinⅤ-FITC凋亡试剂盒购自美BD Bioscience公司(美国);RIPA裂解液购于碧云天公司;兔抗mGluR6、鼠抗β-Actin均购自Santa Cruz Biotechnology公司(美国);ECL化学发光底物试剂盒购自Pierce公司(美国)。
3 大鼠胚胎皮质NPCs的分离和培养 胚胎分离自孕15d大鼠,动物处理符合伦理要求和原则。从这些胚胎中分离脑组织并将其置于DMEM/F12中,去除所有脑膜后,分离大脑皮层,用DMEM/F12冲洗两次;将分离的大脑皮层切成小块,37℃胰酶消化5 min,用吸管吹打分散使组织分散为单细胞。用200目筛网过滤后,将细胞悬液移到离心管中,800转离心5min后,重悬于完全培养基(DMEM/F12基础培养基中加入20ng/ml EGF,10ng/ml bFGF,1× B27 supplement,1× N2supplement,100U/ml链霉素,100U/ml青霉素,0.4U/ml肝素)中,将细胞密度调整至1×105/ml后,以4ml/瓶接种于T50培养瓶中,37℃孵育,5~7d后传代。(本研究所用的NSCs均为传1代的细胞)。
4 载体构建和细胞转染 本实验所需mGluR6 siRNA序列由上海吉玛公司合成,序列信息如下:鼠mGluR6siRNA:sense 5’-CCCCAGUGAUGUUCA AUGATT-3’antisense 5’-UCAUUGAACAUCACUGGGGTT-3’隐形对照 siRNA NC-siRNA:sense 5’-UUCUCCGAACGUGUCACGUTT-3’antisense 5’-ACGUGACACGUUCGGAGAATT-3’过 表 达 载体pCMV2-GV146-GFP-mGluR6由上海生工生物工程有限公司构建。细胞转染前,6孔板中NSCs细胞数量为(0.8~2.4)×105,生长1d后加入过表达质粒DNA 4μg或者干DNA片段4μl以及TurboFectTM 6 μl进行转染,并通过qRT-PCR方法检测转染效率。
5 RNA提取及qRT-PCR 采用Omega试剂盒提取细胞总RNA,测量RNA浓度。根据生物信息学设计合成mGluR6和β-actin的qRT-PCR引物,序列如 下: mGluR6-F5 ’-GGACGCCAAGTAGCAAGGTT-3’ mGluR6-R5 ’-TCCGATGGTCTCTGTGGATCT -3’β-Actin-F5’-GGAGATTACTGCCCTGGCTCCTA-3’β-Actin-R5’-GACTCATCGTACTCCTGCTTGCTG-3’,使用TaKaRa公司逆转录试剂盒与荧光实时定量PCR试剂盒进行逆转录及qRTPCR,按照SYBR Premix Ex TaqTMⅡ进行反应,条件为:95℃,1min预变性,95℃变性10s,60℃退火,延伸40s,重复40个循环,采用2-ΔΔCt的方法计算 mGluR6的表达量。
6 应用流式细胞术分析mGluR6影响鼠NSCs的细胞周期 按照2×104个/孔将传1代的NSCs种入6孔板中,每孔单细胞悬液为2ml,培养48h后加药物干预。实验分组:正常对照组、阴性对照组、siRNA组,加入药物后继续培养。24h后,处理各组单细胞悬液,PI染色,流式细胞术分析mGluR6对鼠NSCs细胞周期的影响。
7 MTT实验检测mGluR7对人胚胎NSCs细胞活力的影响 按照2×104个/孔将传1代的NSCs种入96孔板中,每孔单细胞悬液为200μl,常规培养48h后,给予药物干预。实验分为三组:正常对照组、阴性对照组、siRNA组。每组设3个复孔,在转染后24h、48h和72h后上机检测鼠胚胎NSCs细胞增殖能力。
8 神经球直径测量 神经球直径测量实验步骤如下:将传1代的NSCs分散成单个细胞,按照20000个/孔种入96孔板中,置于37℃、5%CO2孵箱中培养48h后给予药物干预。实验分组:正常对照组、阴性对照组、siRNA组。分别于转染后24h,48h和72h后,通过Image-Pro Express(IPP)图像分析软件并取图片,进而研究mGluR6对各组神经球直径的影响。
9 通过流式细胞术分析mGluR6对鼠NSCs细胞凋亡的影响 细胞培养及分组同上,最后加入400μl的1×binding buffer,反复混匀重悬细胞,再加5μl Annexin V-FITC,于4°C避光染色15min。最后加入10μl PI染液,避光4°C染色5min,采用流式细胞仪检测,分析mGluR6对鼠NSCs细胞凋亡的影响。
10 统计学方法 采用SPSS 13.0统计学软件进行数据分析,数据均以均数±标准误表示,利用单因素方差分析(One-Way ANOVA)进行差异分析,P<0.05表示差异有统计学意义。
结 果
1 过表达或沉默mGluR6后其表达变化 为了研究mGluR6对大鼠胚胎NSCs生长对影响,我们构建了mGluR6的干扰RNA表达质粒(mGluR6siRNA)、过表达质粒(mGluR6)及对照质粒。我们对大鼠胚胎NSCs细胞分别转染了干扰RNA(mGluR6 siRNA)、过表达(mGluR6)和对照质粒。通过qRTPCR和Western Blot检测 mGluR6的过表达及沉默效率,结果表明大鼠胚胎NSCs细胞转染过表达质粒后,在mRNA和蛋白水平mGluR6的表达量均显著升高(图1A,B);转染 mGluR6siRNA后,在 mRNA和蛋白水平mGluR6的表达量均显著降低(图1A,B),证实了mGluR6的过表达载体和mGluR6siRNA是有效的,并将用于后续的细胞功能实验。
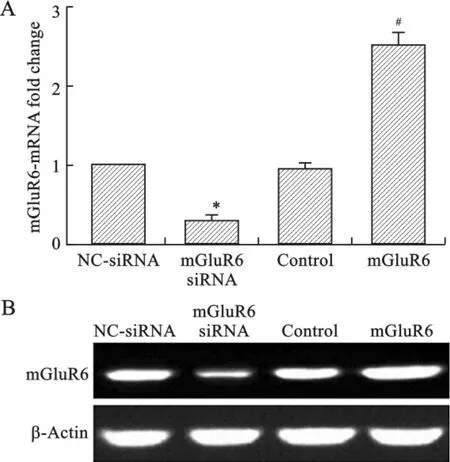
图1 在人NSCs中过表达或沉默mGluR6后其表达变化
2 mGluR6对大鼠NSCs增殖的影响 我们通过MTT实验检测过表达或沉默mGluR6对大鼠NSCs增殖的影响。结果表明,大鼠胚胎NSCs细胞转染过表达mGluR6质粒48h后,显著促进了细胞活力(图2A);转染mGluR6siRNA质粒48h后,显著抑制了细胞活力(图2B)。为了进一步检测mGluR6对大鼠神经球增殖的影响,我们测量了神经球直径,结果表明转染过表达质粒组的神经球直径明显比对照组大(图3A),而转染mGluR6siRNA质粒组的神经球明显小于对照组(图3B)。这表明mGluR6促进了大鼠NSCs增殖。
3 mGluR6对大鼠NSCs细胞周期的影响 为了研究mGluR6对大鼠NSCs细胞周期的影响,我们将mGluR6过表达和干扰siRNA分别转染至大鼠NSCs细胞中,通过流式细胞仪进行检测,结果表明转染mGluR6过表达质粒后,G1/G0期和细胞比率显著下降,S期细胞比率显著上升,促进了细胞周期G1到S期转换(图4、5);而转染 mGluR6siRNA 后,G1/G0期和细胞比率显著增加,S期细胞比率显著下降,抑制了细胞周期G1到S期转换(图6、7)。
4 mGluR6对大鼠NSCs细胞凋亡的影响 为了研究mGluR6对大鼠NSCs细胞凋亡的影响,我们将mGluR6过表达质粒和干扰siRNA分别转染至大鼠NSCs细胞中。通过流式细胞仪进行检测,和对照组相比,转染mGluR6过表达质粒的细胞早凋和晚凋比率均显著下降(图8、9);而转染mGluR6siRNA的大鼠NSCs早凋和晚凋比率均显著增加,特别是晚凋比率显著增强(图10、11)。
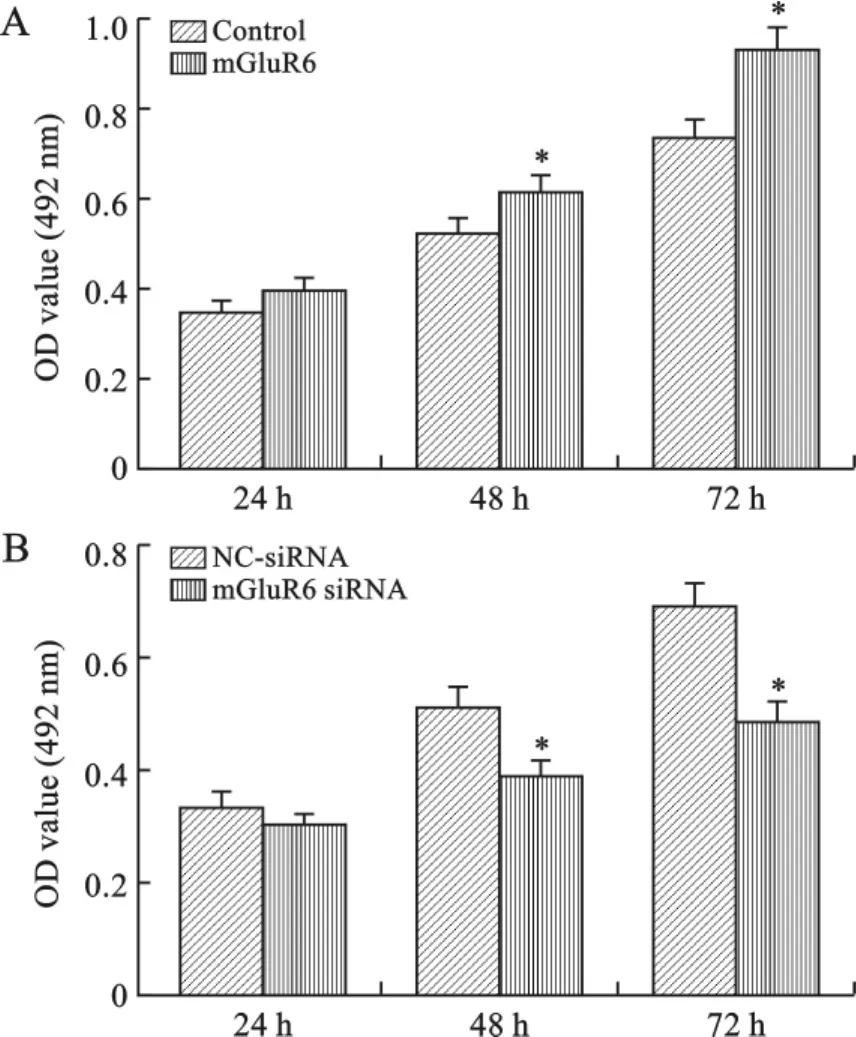
图2 mGluR6对大鼠NSCs增殖的影响(MTT比色)
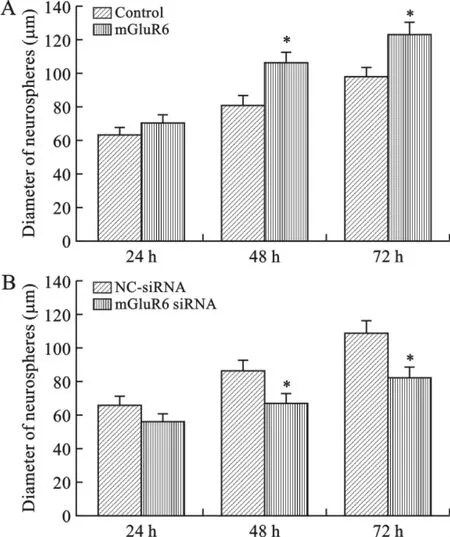
图3 mGluR6对大鼠神经球增殖的影响(神经球直径)

图4 流式细胞仪检测大鼠NSCs的Control组和mGluR6组周期图谱
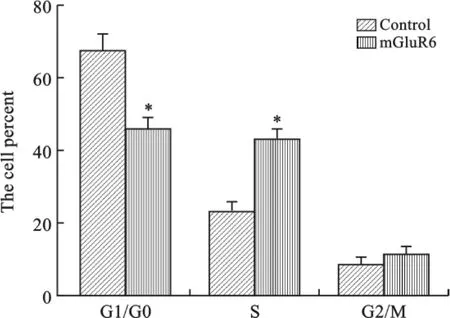
图5 转染mGluR6表达载体对大鼠NSCs周期影响(*P<0.01)
讨 论
在发育和成熟哺乳动物神经系统中,神经干细胞(Neural stem cells,NSCs)的研究为中枢神经系统(CNS)的损伤、治疗和修复带来了希望。目前,NSCs已经被广泛用来治疗Huntington病、缺血性中风、脱髓鞘病及脑等CNS疾病,NSCs的治疗手段已被广大同行认可,特别在改善神经损伤、脑缺血以及神经变性紊乱相关疾病后的神经功能[12-16]。研究NSCs并应用在CNS替代治疗中,基因调控及蛋白质功能改变对NPCs的增殖起着非常重要的作用。所以,探究NPCs增殖过程中的分子网络及调控机制在神经系统损伤修复及神经退行性疾病的治疗等方面具有重要价值。
随着研究的不断深入,发现谷氨酸可能通过mGluRs对脑损伤后的神经发生有一定的调控作用[17-18]。本 小 组 前 期 研 究 中 发 现,mGluR3[19]、mGluR5[20]和 mGluR7[21]均与 NSCs的生物学功能有着密切的关系。mGluR6是代谢性谷氨酸受体第III组的重要成员。自1993年被Nakajima等人报道mGluR6在大鼠视网膜中生物学功能以来[6],许多研究表明mGluR6在视杆和视锥细胞的突触传递过程中至关重要。最近,Devi等人发现mGluR6在人类黑色素细胞中表达,且能够促进黑色素的含量[22]。但是,mGluR6是否参与NSCs生长及其调控机制还不清楚,值得进一步深入研究。
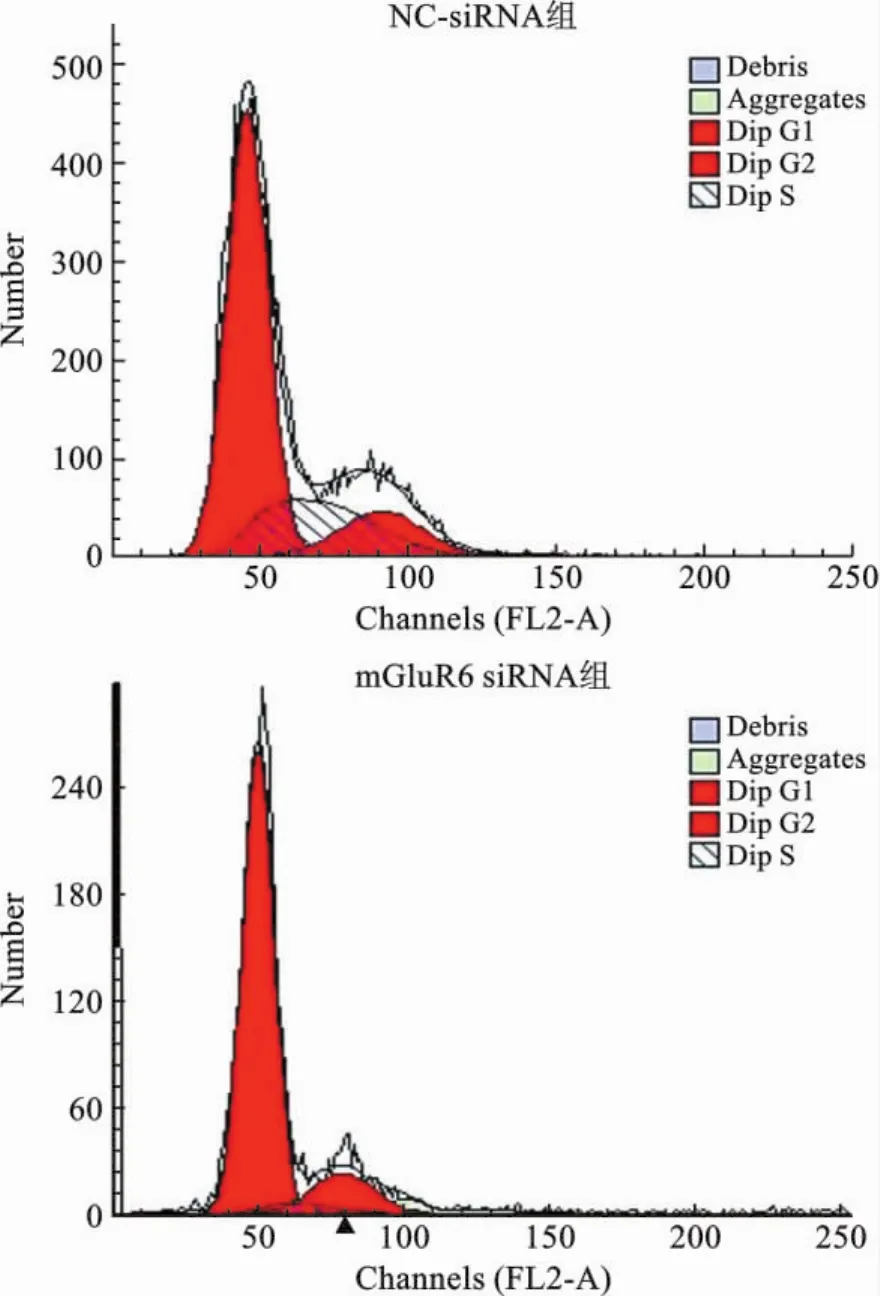
图6 流式细胞仪检测大鼠NSCs的NC-siRNA组和mGluR6siRNA组周期图谱
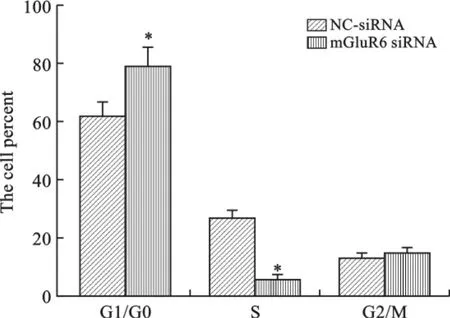
图7 转染mGluR6siRNA对大鼠NSCs周期的影响(*P<0.01,n=3)
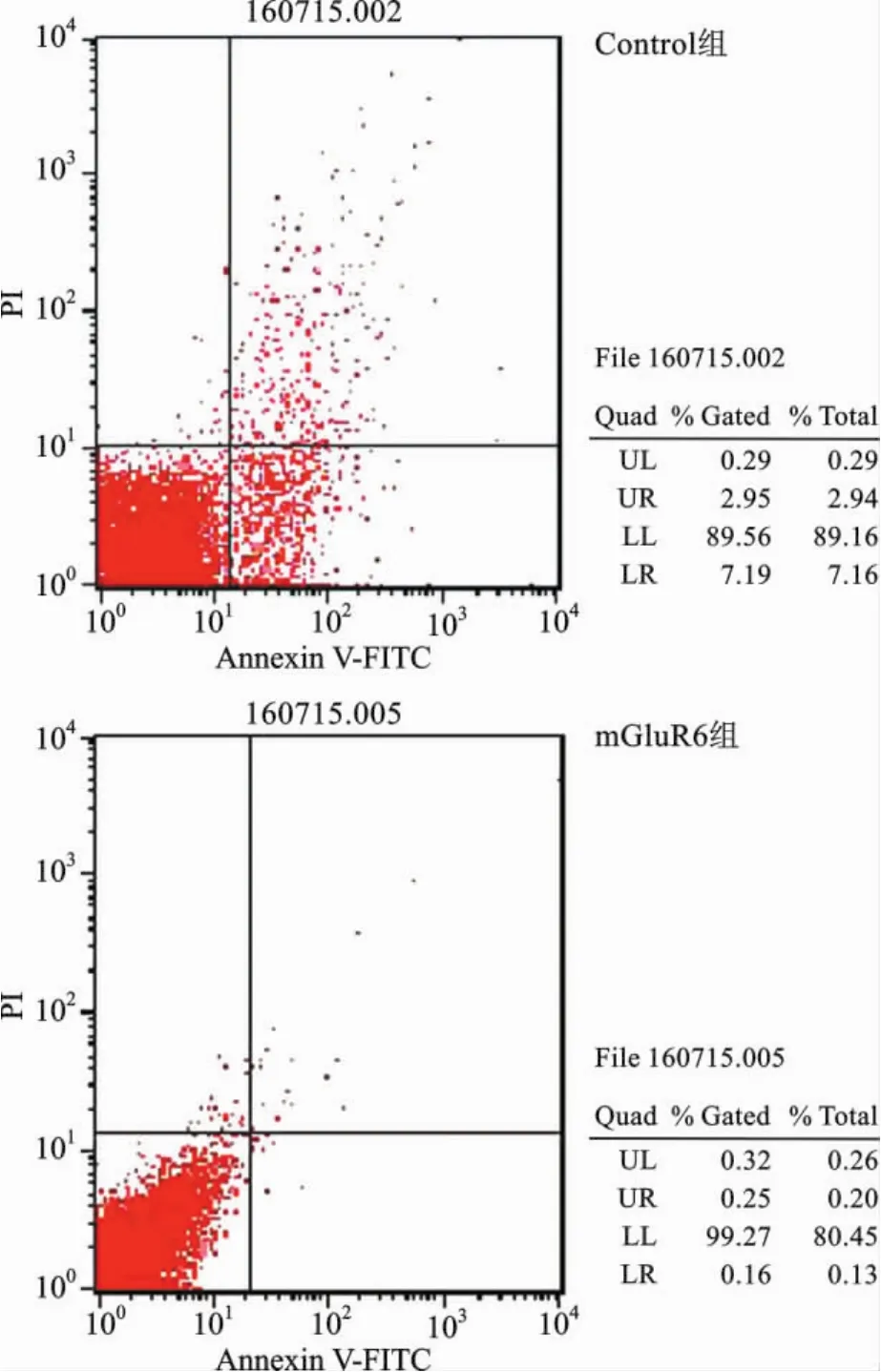
图8 流式细胞仪检测大鼠NSCs的Control组和mGluR6组凋亡图谱
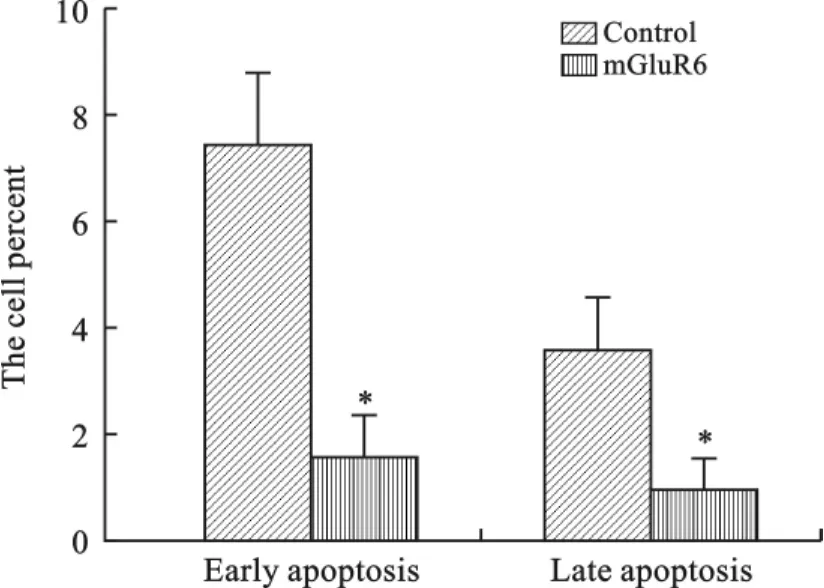
图9 转染mGluR6表达载体对大鼠NSCs凋亡的影响(*P<0.01,n=3)
Zhao等研究发现了mGluR5促进了人胚胎神经干细胞前体细胞的增殖及其机制[20]。为研究mGluR6对神经干细胞生物学功能的影响,本研究选择以大鼠胚胎神经干细胞(NSCs)为研究材料,通过构建mGluR6过表达和siRNA处理大鼠胚胎NSCs,分析其对细胞增殖的影响。研究发现,mGluR6过表达载体能增加体外培养的大鼠NSCs的细胞活力和神经球的大小。相反,mGluR6siRNA降低了NSCs的细胞活力和神经球的大小。这些研究结果表明,mGluR6可促进体外培养的大鼠胚胎NSCs的增殖。
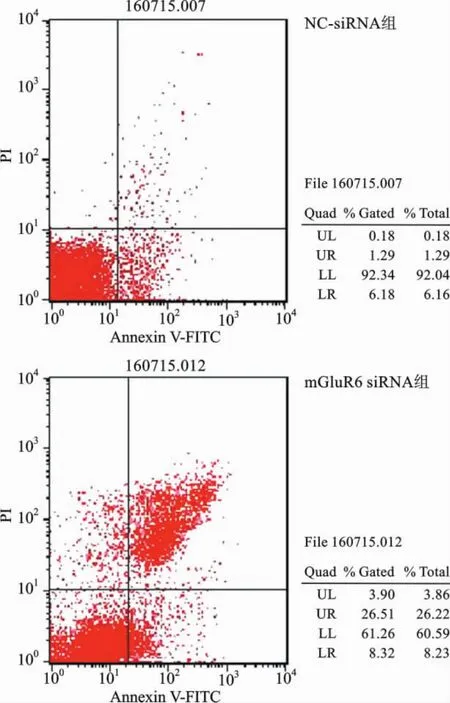
图10 流式细胞仪检测大鼠NSCs的NC-siRNA组和mGluR6siRNA组凋亡图谱
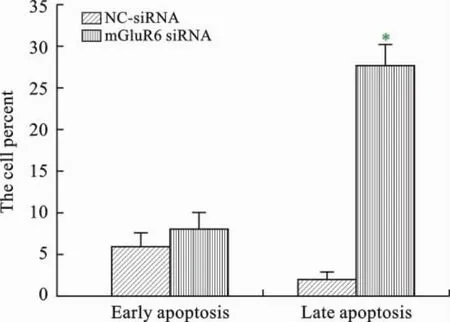
图11 转染mGluR6siRNA对大鼠NSCs凋亡的影响(*P<0.01,n=3)
细胞周期中G1向S期的过渡是调节细胞周期的重要节点,当G1/S节点通过后,细胞周期依次进行,然后逐渐回到G1期,这个阶段不容易受到其他调节因素[23]的影响。M期表明细胞处于增殖和分裂阶段,S期表明细胞开始新一轮的DNA复制和分裂。本研究发现,在 mGluR6siRNA处理NSCs后,G1/G0期细胞比率升高,S期细胞比率下降,mGluR6siRNA将人NSCs细胞阻滞在G1/G0期,细胞的DNA复制合成与细胞分裂显著减少。但与之对应的是,过表达mGluR6后,与对照组相比G1/G0期细胞比率显著下降,S期细胞比率显著上升,mGluR6过表达促进了细胞G1到S期转化。
综上所述,mGluR6同其它mGluRs在促神经发生方面主要可能通过以下功能来实现:①直接的促增殖作用;②促NSCs/神经祖细胞向神经元分化;③促存活、抗凋亡神经保护作用。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
科学与社会(2021年3期)2021-12-02
皮肤性病诊疗学杂志(2021年5期)2021-11-27
昆明医科大学学报(2021年8期)2021-08-13
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2021年1期)2021-01-27
中国生殖健康(2020年7期)2021-01-18
中国生殖健康(2020年5期)2021-01-18
三农资讯半月报(2020年11期)2020-06-21
