
高温胁迫对小黑麦光合作用影响
2020-04-30 08:40凌悦铭张亚黎王震任丽彤艾尼瓦尔李召锋孔广超
石河子大学学报(自然科学版) 2020年2期
凌悦铭,张亚黎,王震,任丽彤,艾尼瓦尔,李召锋,孔广超*
(1 石河子大学农学院/新疆生产建设兵团绿洲生态农业重点实验室,新疆 石河子 832003;2 石河子大学化学化工学院,新疆 石河子 832003)
21世纪末,全球气温极有可能会升高超过3 ℃[1],而气温每升高1 ℃,全球小麦产量预计将下降4.1%~6.4%[2]。小麦灌浆期最适宜温度为18~22 ℃,上限温度为26~28 ℃,日平均气温在30 ℃以上时,小麦就会提早结束灌浆,千粒重降低[3]。小麦在灌浆期遇到极端高温或者连续性高温天气,产量就会显著下降,而在灌浆前期遭受高温较后期高温对产量影响更大[4]。
作物产量主要源于光合作用。小麦旗叶具有较高的光合产物供应能力,是光合产物源的主要供应者。植物光合作用对高温最敏感[5],小麦在灌浆期如遇到长期高温胁迫,其叶片的Pn及叶绿素含量均会显著下降[6]。同时,高温胁迫还会加速光合器官的衰老,此外由于高温下叶片的蒸腾需求增加,还会影响植物水分状况进而影响光合作用,缩短光合作用的持续时间[7-8]。
小黑麦是由小麦和黑麦经人工属间杂交和染色体加倍培育的新作物,其不但结合了小麦的籽粒高产、优质和黑麦的抗病逆性强、适应性广的优点,而且具有生长优势明显、光合作用强等特点[9],是新型粮食与饲草作物。据联合国粮农组织统计,2017年全世界小黑麦播种面积为416.58万hm2,自2013年到2017年,中国小黑麦年种植面积从19万hm2发展到49万hm2,增加了约1.6倍。近年来对小黑麦在酸雨[10]、干旱[11-13]和盐[14-15]等胁迫下的影响等方面研究报道较多。小黑麦同小麦一样,同属于冷凉季作物。小黑麦在灌浆前期遭受高温胁迫,导致灌浆中后期籽粒中淀粉合成相关酶活性下降,籽粒中淀粉含量降低以及千粒重、容重下降[16]。作物干物质积累和产量形成主要来源于光合作用[17]。当前关于小黑麦在高温胁迫下光合作用的研究还没有见文献报导。本试验以4个六倍体小黑麦品种为材料,探讨高温胁迫对小黑麦光合作用影响,旨在为小黑麦耐热性评价以及耐热品种的培育提供理论依据。
1 材料与方法
1.1 材料
石河子大学麦类作物研究所选育的4个春性六倍体小黑麦品种(系),分别为新小黑麦3号、新小黑麦4号和新小黑麦5号及品系扩繁1号。
1.2 试验设计与田间管理
试验于在石河子大学农学院实验站(44°17′N,86°03′E)进行。采用随机区组试验设计,重复4次。小区面积4 m2,行长4 m,行距20 cm,播种量为562.5万粒/hm2。试验设人工增温和正常温度2个处理,田间管理同当地大田生产。在各品种(系)开花后5~14 d于小区中搭设透光PVC膜增温棚进行高温处理[16],未覆盖PVC膜增温棚的小区作为实验对照区。
1.3 测定项目与方法
在高温胁迫至第10天以及解除胁迫后第10天,在高温胁迫及对照区各重复中均选取长势均匀、生育进程一致的植株。采用美国LI-COR公司LI-6400光合测定系统测定旗叶中部净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs),并计算气孔限制值(Ls)[18]。每小区测定10株。测定过程选用红蓝光源,叶室中的PAR(photosynthesis active radiation)设为1200 μmol·m-2·s-1。测定时间为10:30—12:30。重复4次,取平均值。
1.4 数据统计分析
用Excel 2010进行数据统计,用SPSS 16.0完成方差分析和Duncan多重比较,作图用Origin 2019进行。
2 结果与分析
2.1 高温胁迫对小黑麦旗叶Pn和Tr影响
在高温胁迫条件下,4个六倍体小黑麦品种的Pn较对照下降了11.4%~39.8%(图1a),下降幅度达到了极显著水平。在参试的4个小黑麦品种中,高温胁迫导致Pn下降幅度由大到小依次是品系扩繁1号,下降幅度为39.8%,其次为新小黑麦4号,下降了17.2%,新小黑麦5号,下降了14.4%,新小黑麦3号下降了11.4%,其中品系扩繁1的Pn在高温胁迫下下降幅度显著大于其它3个品种,其余3个小黑麦品种在高温胁迫下的Pn下降幅度虽有所差异但其差异均不显著。
由图1b可见,高温胁迫下,新小黑麦5号的Tr较对照下降了1.1%,新小黑麦3号下降了3.1%,下降幅度均未达到显著水平;新小黑麦4号和扩繁1号在高温胁迫下旗叶的Tr较对照下降幅度分别为15.0%和26.2%,达到了极显著水平,其中以新小黑麦4号在高温胁迫下的Tr下降幅度最大。
2.2 高温胁迫对小黑麦旗叶Ci和Gs影响
高温胁迫下,参试的4个六倍体小黑麦品种旗叶的Ci较对照升高了2.0%~6.8%(图2a),但除新小黑麦4号在高温胁迫下Ci的升高幅度尚未达到显著外,其它3个品种旗叶的Ci升高幅度均较对照达到了显著或极显著水平,其中以扩繁1和新小黑麦5号的Ci升高幅度分别为6.8%、3.7%,均较对照达到了极显著水平。
由图2b可见,在高温胁迫下,4个六倍体小黑麦品种旗叶的Gs变化不同,其中扩繁1号和新小黑麦4号旗叶的Gs有下降趋势,分别下降了0.9%、2.4%,而新小黑麦3号和新小黑麦5号的叶片Gs则呈上升趋势,分别上升了1.5%、2.2%,但均未达到显著水平。可见,高温胁迫对小黑麦旗叶的Gs影响不大。
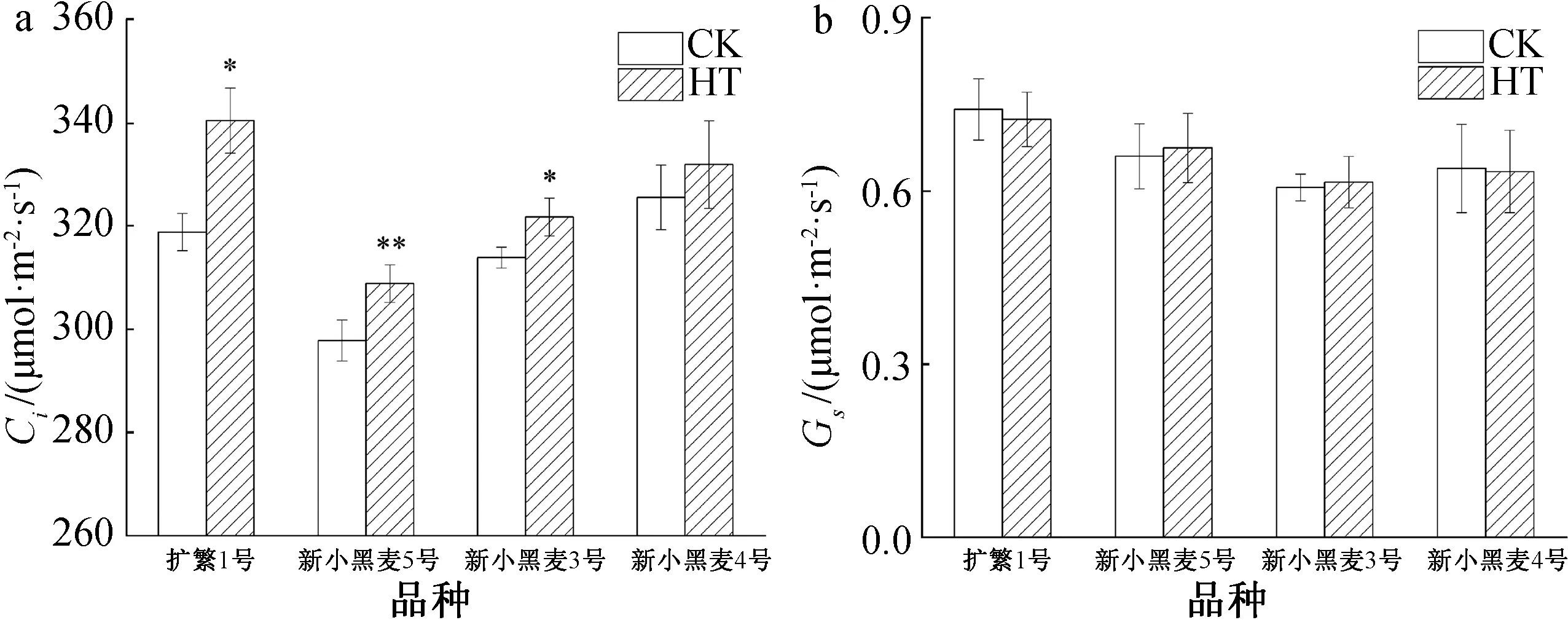
CK—对照;HT—高温胁迫;*表示在5%水平下显著;**表示在1%水平下显著。图2 高温胁迫下小黑麦Ci和Gs
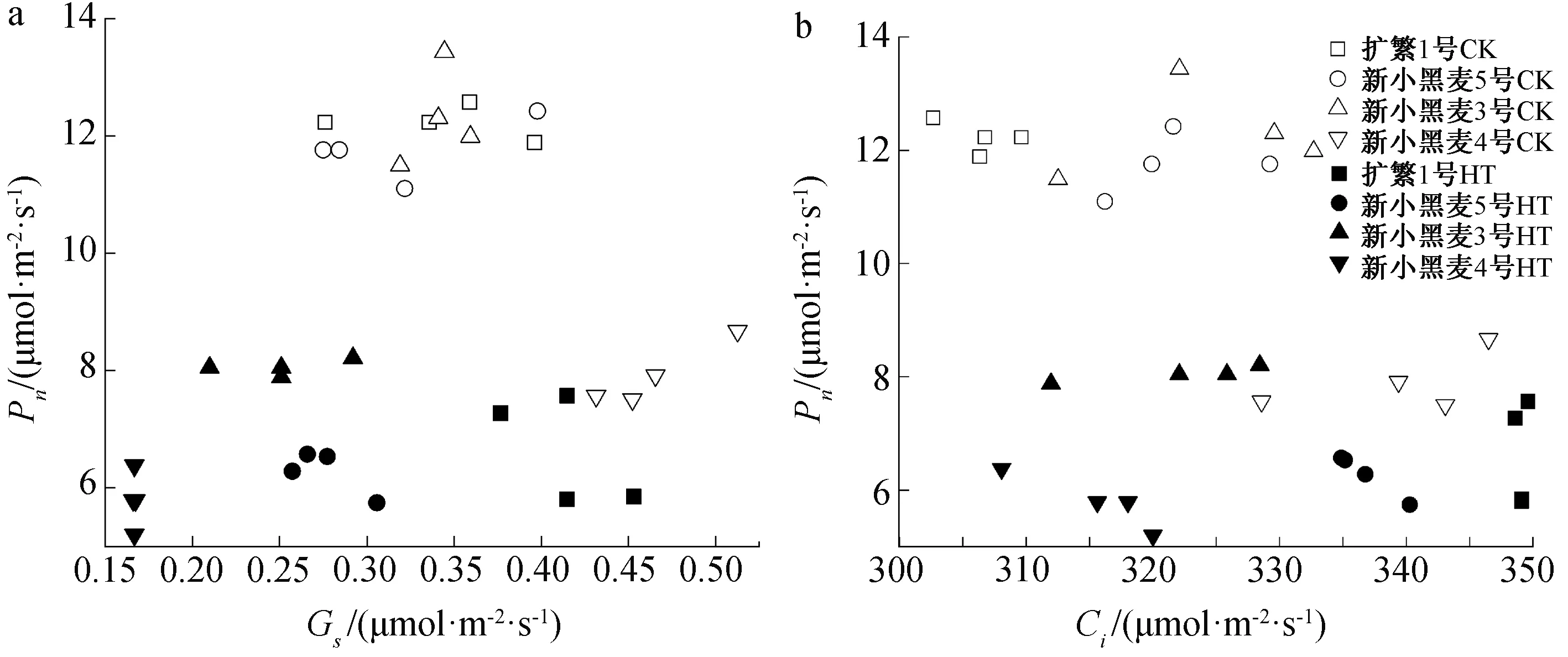
2.3 高温胁迫下小黑麦旗叶Pn与Gs、Ci的变化关系
从4个六倍体小黑麦品种叶片的Pn与Gs的变化关系看(图3a),二者之间没有明显的直接关系。但从Pn与Ci变化的关系看(图3b),无论在高温胁迫下还是对照条件下,总体表现出随小黑麦叶片中Ci的增大其叶片Pn呈现下降趋势,同时品种间Pn与Ci差异均较大,其中品种新小黑麦5号的Pn最高,而Ci最小。
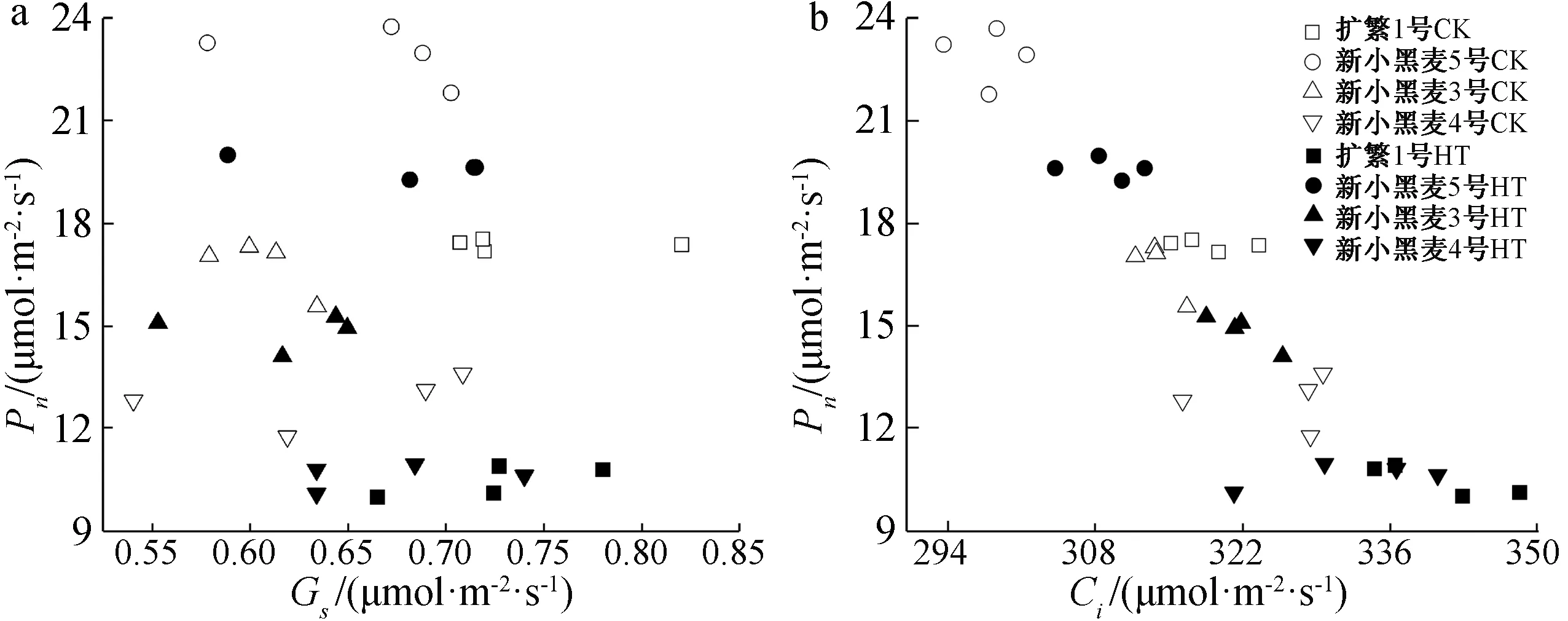
CK—对照;HT—高温胁迫;*表示在5%水平下显著;**表示在1%水平下显著。图3 小黑麦旗叶Pn与Gs、Ci的关系
在高温胁迫下,4个参试小黑麦品种的气孔限制值(Ls)均较对照有所下降(表1),但除扩繁1号在高温胁迫下的Ls下降幅度较对照达到显著水平外,其余3个品种的Ls下降幅度与对照均未达到显著水平。在高温胁迫解除后10 d时,扩繁1号和新小黑麦5号旗叶的Ls仍均较对照下降,而品种新小黑麦3号、新小黑麦4号的Ls较对照有所上升,但变化幅度均未达到显著水平。

表1 高温胁迫下小黑麦旗叶气孔限制值
注:CK为对照,HT为高温胁迫;同一列中的小写字母表示在5%水平下的差异显著,大写字母为1%水平下的差异显著。
2.4 高温胁迫后小黑麦旗叶光合参数变化
在高温胁迫结束后10 d时,前期高温胁迫下小黑麦叶片的Pn仍较对照低26.8%~46.6%,且其差异达到了极显著水平(见图4a)。在4个参试的六倍体小黑麦品种间,Pn下降幅度由大到小依次为新小黑麦5号、扩繁1号、新小黑麦3号、新小黑麦4号,其中新小黑麦3、新小黑麦4号旗叶Pn下降幅度显著低于扩繁1号和新小黑麦5号。
在高温胁迫处理结束10 d后,除扩繁1号的Tr较对照仍有所降低但差异不显著外,其他3个小黑麦品种因高温胁迫导致的Tr仍显著或极显著低于对照(图4b)。
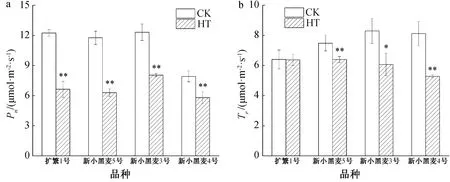
CK—对照;HT—高温胁迫;*表示在5%水平下显著;**表示在1%水平下显著。图4 高温胁迫结束后的小黑麦Pn和Tr
在高温胁迫结束后,新小黑麦3号旗叶的Ci基本恢复至对照水平(图5a),而扩繁1号、新小黑麦5号叶片Ci仍极显著高于对照,其升高幅度分别为4.7%、13.9%,但新小黑麦4号的Ci却极显著低于对照。
由图5b可知,在高温胁迫结束后,除扩繁1号旗叶的Gs较对照有所升高外,其他3个小黑麦品种的Gs均较对照有不同程度的下降,其中新小黑麦3号、新小黑麦4号的Gs较对照下降幅度分别为26.4%、64.2%,分别达到了显著或极显著水平。
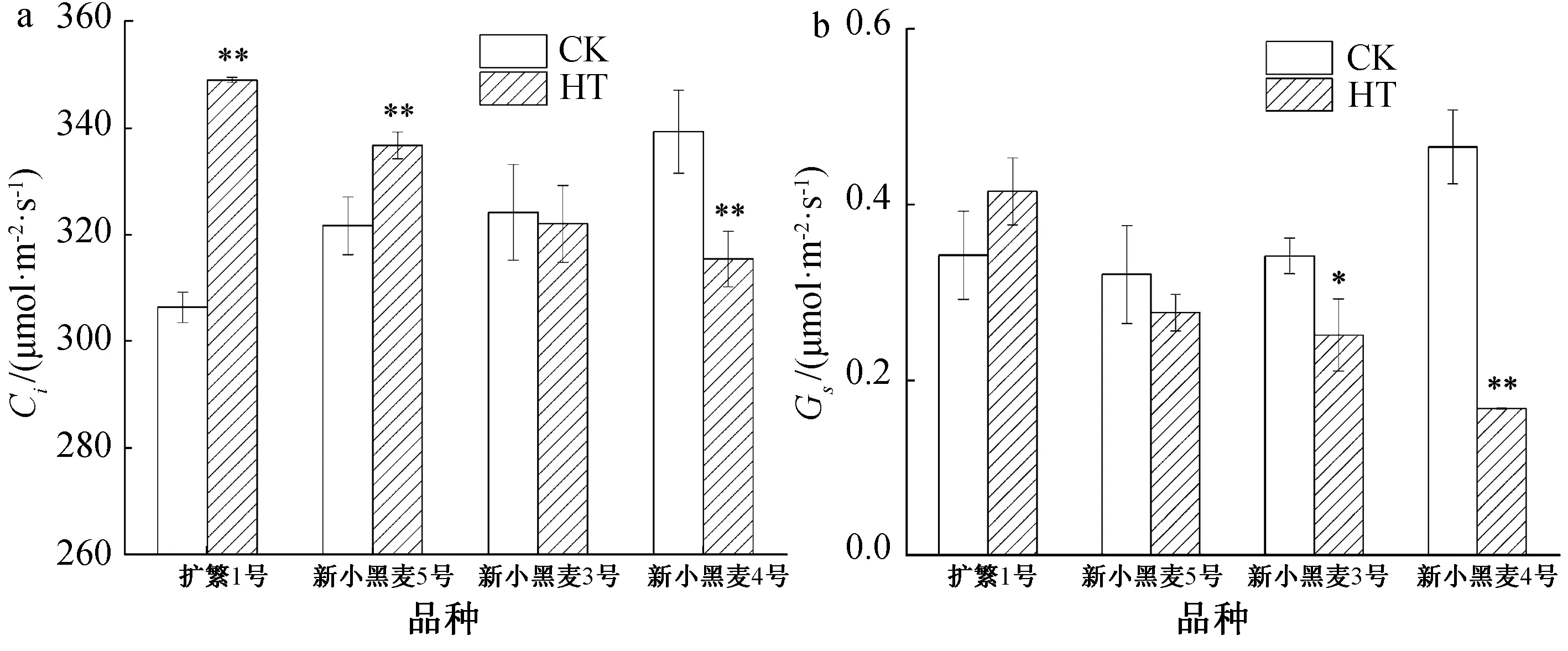
CK—对照;HT—高温胁迫;*表示在5%水平下显著;**表示在1%水平下显著。图5 高温胁迫后小黑麦Ci和Gs
图6a显示,高温胁迫结束后,除新小黑麦3号、新小黑麦4号的Pn随Gs的下降而下降外,其它2个小黑麦品种的Pn与Gs变化无明显关系。图6b显示,扩繁1号、新小黑麦5号的Pn仍随Ci的升高而下降,而新小黑麦4号的Pn随Ci的升高而升高,但新小黑麦3号的Pn随Ci的升高没有显著变化。
CK—对照;HT—高温胁迫;*表示在5%水平下显著;**表示在1%水平下显著。图6 高温胁迫解除后小黑麦Pn与Ci、Gs的关系
3 结论与讨论
光合作用是作物生长发育、干物质积累和产量形成的基础[17]。Pn是直接判断植株光合能力大小的稳定的遗传性状[19],可作为高光效育种的指标性状。提高叶片Pn,充分发挥叶片的光合潜能,对于提高作物产量具有重要意义[20]。引起植物净光合速率下降的原因主要分为气孔因素和非气孔因素,气孔因素指外界胁迫引起气孔关闭,CO2供应受阻;非气孔因素主要指叶肉细胞间隙气相空间和CO2扩散阻力增大,内部的酶活力下降[21-22]。气孔因素与非气孔因素判别的指标为Ci和Ls两方面[18]。本试验发现高温胁迫导致4个小黑麦品种(或品系)旗叶的Pn下降、Ci升高、而Gs、Ls多无显著变化,同时无论是在高温胁迫下还是对照中均存在随Ci增大Pn呈现下降趋势。可见高温胁迫下小黑麦旗叶Pn的下降主要由非气孔因素所致。这与非气孔因素是造成小麦灌浆期高温胁迫下光合速率降低的主要原因的结论一致[23]。也有研究发现,短期或轻度逆境胁迫下时,光合速率的下降是由气孔限制引起的[24-26],而随高温胁迫时间增加,或者重度高温胁压加大时,光合速率降低主要是受非气孔因素的影响[27]。短期或轻度高温胁迫引起的光合速率下降幅度较小且可恢复[28],连续极端高温可导致植物净光合速率急剧下降,且高温胁迫对光合器官造成的伤害则是不可逆的[29-30]。
本研究发现,在高温胁迫结束后10 d,新小黑麦3号、新小黑麦4号旗叶的Gs较对照显著降低,而Ls较对照差异不显著,这表明气孔导度降低可能是导致光合速率降低的主要原因。而从另外2个小黑麦品种扩繁1号、新小黑麦5号的Ci仍显著高于对照,Ls也低于对照来看,前期的高温胁迫对扩繁1号、新小黑麦5号旗叶叶片光合机构造成严重且不可恢复的破坏,从而导致高温胁迫解除后其旗叶Pn仍显著低于对照。综上所述,10 d高温胁迫致使对损伤修复能力较差的小黑麦叶片光合机构的活性受到了抑制,对叶片光合器官造成的伤害不可逆;损伤修复能力较强的小黑麦品种也因前期高温胁迫而导致CO2供应不足致使光合速率下降。
光系统Ⅰ(PSⅠ)、光系统II(PSII)、电子传递和光合CO2固定等任意一个结构受到高温胁迫损伤都足以抑制整个光合代谢过程[31]。高温处理下水稻叶片PSII潜在活性(Fv/Fo)和PSII光能转化效率(Fv/Fm)降低,且随着高温胁迫时间的持续和叶片功能的衰退,类囊体膜结构损伤越严重,光能转化效率越低[32]。适宜光强下短时亚高温处理不会产生明显光抑制,高温胁迫会对黄瓜幼苗PSII反应中心造成严重损伤,光合酶受高温胁迫诱导,其诱导效应与温度升高幅度和高温持续时间有关[27]。可见非气孔因素限制光合作用主要发生在植物受到较严重的胁迫时,其光合作用代谢过程被抑制和损伤[33],光合作用受到植株内部的酶活力和光合组分控制的影响[34],即叶肉细胞气体扩散抗阻增加、CO2溶解度降低、Rubisco(核酮糖-1,5-二磷酸羧化酶/加氧酶)对CO2亲和力降低、RuBp羧化酶(核酮糖-二磷酸羧化酶)再生能力下降或光合机构关键成分的稳定性在逆境下降低导致光合速率下降[35-36]。本研究也发现,在高温胁迫下小黑麦叶片叶绿素荧光参数F0显著上升、Fv/Fo、Fv/Fm多显著下降。这些均表明高温胁迫下小黑麦旗叶的PSII反应中心失活与光能转化率下降,出现了光抑制现象。
高温胁迫下,参试的4个小黑麦品种间在Pn下降,Tr下降,Ci升高,这些指标变化趋势虽然一致,但品种间影响程度不同,可见每个品种对高温的耐受性具有差异,其中新小黑麦3号耐热性较强,新小黑麦5号、新小黑麦4号次之,扩繁1号最弱;同时品种间对高温损伤修复的性能上也有差异,其中新小黑麦4号对损伤修复能力较强,新小黑麦3号、扩繁1号次之,新小黑麦5号最弱。新小黑麦3号、新小黑麦4号通过降低气孔导度和蒸腾速率从而减少水分散失可能是其对损伤修复能力较强的原因之一。这为选育耐高温小黑麦品种提供了理论支持。
猜你喜欢
航空材料学报(2022年4期)2022-08-04
贵州农业科学(2022年4期)2022-05-06
干旱地区农业研究(2022年1期)2022-01-28
股市动态分析(2021年25期)2021-12-30
汽车与安全(2019年11期)2019-06-01
科技创新与品牌(2019年12期)2019-02-06
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
股市动态分析(2014年27期)2014-07-29
