
8个高山杜鹃杂交F1代的叶片性状遗传分析
2020-08-03 02:26王继华彭绿春蔡艳飞李世峰解玮佳
江西农业学报 2020年7期
许 凤,宋 杰,王继华,彭绿春,张 露,蔡艳飞,李世峰*,解玮佳,2*
(1.云南省农业科学院 花卉研究所/国家观赏园艺工程技术研究中心/云南省花卉育种重点实验室,云南 昆明 650205;2.云南大学 生命科学学院/云南省高校植物病虫害防控工程研究中心,云南 昆明 650091)
高山杜鹃系指由杜鹃花科(Ericaceae)、杜鹃花属(Rhododendron)中杜鹃花亚属(SubgenusRhododendron)和常绿杜鹃亚属(SubgenusHymenanthes)的种质资源类型及由其育成的一些叶片革质、常绿、花冠硕大的植物[1]。高山杜鹃因其种类繁多、花大色艳和花姿优美等特性而具有较高的观赏和园林应用价值[2]。欧美国家对高山杜鹃的应用历史悠久,品种极为繁多[3]。而在我国,高山杜鹃大多仍处于野生状态,对其的园艺应用还处于初期起步阶段,故其品种基本上靠国外引进[2-3]。云南是世界杜鹃花的分布中心和分化中心之一,杜鹃花种类极为丰富。发掘优良野生种质,并与已有品种开展杂交育种,是培育具有市场竞争力和知识产权的高山杜鹃新品种的有效途径[4]。在此背景下,笔者所在项目组以高山杜鹃品种‘安娜’(R. ‘Anah Kruschke’)(矮小紧凑、紫花)为母本,以‘门铃’(R. ‘Jingle Bells’)(矮小紧凑、橙花)、‘红粉佳人’(R. ‘XXL’)(粉花、抗热)和‘莫泽之栗’(R. ‘Moser Maroon’)(红花)3个品种,以及露珠杜鹃(R.irroratum)(黄花)、马缨杜鹃(R.delavayi)(红花、抗旱)和宽杯杜鹃(R.sinofalconeri)(黄花、叶型奇特)3个物种为父本进行了杂交育种工作,希望获得更多性状优异的盆花杜鹃新品种。
我国的高山杜鹃育种研究工作起步较晚,现有研究主要集中在杂交亲和性[5-10]和可育性分析[6,11-16]上,对于杂交后代的性状遗传表现研究不多,现仅有刘晓青等对18个西鹃品种间23个配组杂交F1代的花色、斑点、花径及瓣型等观赏性状的遗传表现进行了分析[17],以及马绍宇等对高山杜鹃品种‘罗伯茨’(R. ‘Lord Roberts’)×大白杜鹃(R.decorum)杂交F1代的花期、花色、斑点、香味等主要观赏性状的遗传进行了分析[18]。但迄今对杜鹃育种早期筛选评价指标叶片性状的遗传研究甚少。为此,我们通过观测与分析以上配组杂交所获得的8个F1代的叶片性状,研究了其子代与亲本的叶片性状表现差异,总结了其遗传规律,可为今后的高山杜鹃杂交育种提供理论依据和借鉴。
1 材料与方法
1.1 试验时间、地点
本试验于2018年10~12月在昆明市晋宁区昆阳镇的云南省农业科学院花卉研究所春禾试验基地进行。
1.2 试验材料
本试验所涉及的高山杜鹃品种及种类的名称、特征及来源如表1所示。

表1 试验所用的高山杜鹃品种及种类的名称、特征及来源
1.3 试验方法
于2014年3~4月在春禾试验基地进行高山杜鹃品种(种)间的人工配组杂交授粉试验(详见表2)。于2014年10~12月获得杂交种子,并于翌年3月播种,培育出各杂交F1代的群体。于2018年10~12月对各杂交F1代群体的叶片性状进行调查。具体调查方法为:从各杂交F1代群体中随机选取植株30棵,再从各单株中随机选取当年生枝的中部叶片3片,用游标卡尺测定其叶片长、叶片宽和叶柄长度。
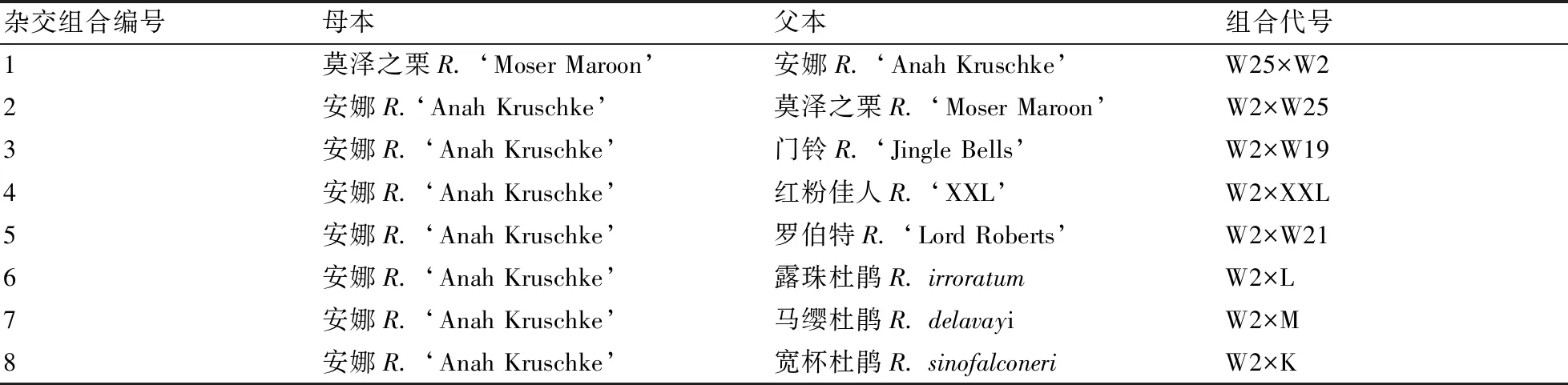
表2 高山杜鹃品种(种)间的人工配组杂交授粉试验
1.4 数据分析
应用Microsoft Excel 2010和IBM SPSS Statistics 19进行数据统计与分析。
2 结果与分析
2.1 各杂交组合杂交F1代的叶片性状遗传分析
2.1.1 W2×W25和W25×W2杂交F1代的叶片性状遗传分析 从W2×W25的杂交F1代叶片性状调查结果(表3)来看:叶片长总平均值占中亲值的104.2%,叶长大于高亲值的个数占43.3%,介于双亲值之间的个数占24.4%,小于低亲值的个数占32.2%,说明其杂交F1代的叶片长表现出了超亲优势;叶片长/宽大于高亲值的个数占52.2%,表现出较强的超亲优势;叶片宽和叶柄长的总平均值占中亲值的百分比分别为98.2%和90.3%,表现为明显的衰退现象。
从W25×W2的杂交F1代的叶片性状调查结果来看:其叶片长、叶片宽和叶柄长小于低亲值的个数占比分别为40.0%、58.9%和66.7%,表现为明显的衰退现象;而叶长/宽表现为超亲优势。
将W2×W25和W25×W2的杂交F1代的结果进行比较后发现,叶长/宽即叶型表现为强的超亲优势,而叶柄长则表现为衰退现象。
2.1.2 W2×W19杂交F1代的叶片性状遗传分析 如表3所示,在W2×W19杂交F1代中,叶片长、宽和叶片长/宽的平均值分别占中亲值的107.0%、105.6%、104.0%,而且其大于高亲值的个数分别占37.8%、33.3%、47.8%,说明杂交F1代在叶片长、宽和叶片长/宽这3个性状上均表现出了极强的超亲优势;但叶柄长的总平均值占中亲值的77.9%,小于低亲值的个数占比为62.2%,表现为衰退现象。
2.1.3 W2×XXL杂交F1代的叶片性状遗传分析 由表3可知:在W2×XXL杂交F1代中,叶片长/宽的平均值占中亲值的124.2%,大于高亲值的比例达到了76.7%,表明在叶型遗传上表现为超亲优势;其余3个性状的平均值占中亲值的比例都小于100%,而小于低亲值的个数占比都超过了50%,整体表现出衰退现象。
2.1.4 W2×W21杂交F1代的叶片性状遗传分析 从表3可以看出: W2×W21杂交F1代的叶片长的平均值略低于中亲值,介于双亲值间的后代个数占了48.9%,大于高亲值的后代占比为22.2%,说明杂交F1代的叶片长具有一定的衰退现象;叶片宽小于低亲值的个数较多,占总调查数量的57.8%,衰退现象比叶片长还要严重,使得叶片长/宽的总平均值占到了中亲值的126.5%,因而在叶型上反而出现了超亲现象;叶柄长小于低亲值的比例为52.2%,总平均值占中亲值的73.2%,所表现出来的衰退现象最严重。
2.1.5 W2×L杂交F1代的叶片性状遗传分析 W2×L杂交F1代的叶片长、宽的平均值分别占中亲值的94.9%、94.2%,均表现出衰退现象;叶片长/宽的平均值占中亲值的101.8%,接近于亲本;但叶柄长的总平均值占中亲值的65.8%,小于低亲值的数量占了72.2%,其衰退现象最为显著(表3)。
2.1.6 W2×M杂交F1代的叶片性状遗传分析 W2×M杂交F1代的叶片长、宽和叶柄长的平均值分别占中亲值的99.7%、98.6%和80.6%,小于低亲值的个数分别占34.4%、25.6%和63.3%,表现出了明显的衰退现象;而叶片长/宽的平均值占中亲值的101.4%,介于双亲值之间的个数占67.8%,表明其叶型遗传较为稳定(表3)。
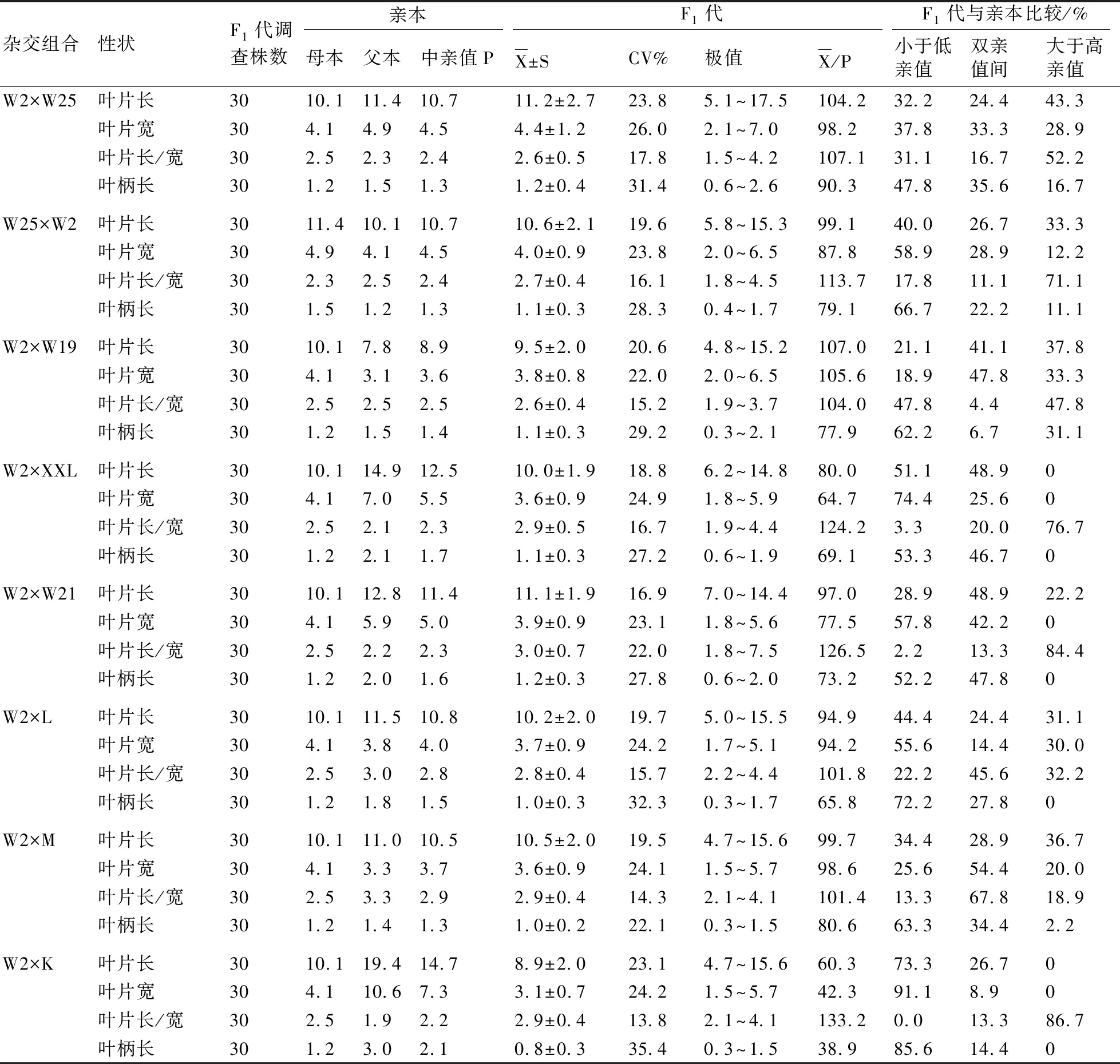
表3 各杂交组合及其杂交F1代的叶片性状比较
2.1.7 W2×K杂交F1代的叶片性状遗传分析 W2×K杂交F1代的叶片长、叶片宽和叶柄长的平均值分别占中亲值的60.3%、42.3%和38.9%,而小于低亲值的个数较多,占比分别为73.3%、91.1%和85.6%,其衰退趋势十分明显;但叶片长/宽的平均值占中亲值的133.2%,小于低亲值的个数为0,表明其杂交F1代的整体叶型表现为超亲现象(表3)。
2.2 不同杂交组合的叶片性状遗传比较
2.2.1 叶片长的遗传比较 如表4中所示,F1代叶片长与父本存在显著差异的组合较多,有W2×W19、W2×XXL、W2×W21、W2×L、W2×K共5个组合,占总杂交组合数的62.5%;而F1代叶片长与母本存在显著性差异的杂交组合有:W2×W25、W2×W21、W2×K,仅占总杂交组合数的37.5%,即与母本无显著差异的占了62.5%,说明F1代叶片长表现出了一定的遗传优势,而且此遗传优势倾向于母本。
2.2.2 叶片宽的遗传比较 F1代叶片宽与父本存在显著差异的组合较多(如表5所示),为W2×W25、W25×W2、W2×W19、W2×XXL、W2×W21和W2×K这6个组合,占所有杂交组合的75%;而与母本存在显著差异的组合为W2×XXL、W2×L、W2×M和W2×K,占全部组合的50%,说明叶片宽不具有明显的遗传优势。

表4 不同杂交组合的F1代与其双亲的叶片长差异性比较
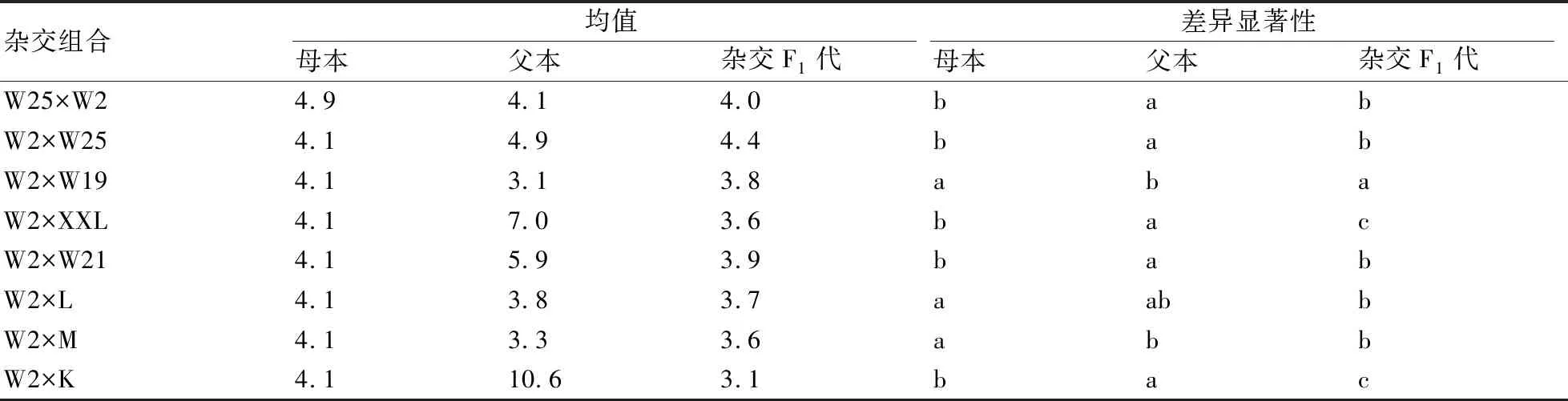
表5 不同杂交组合的F1代与其双亲的叶片宽差异性比较
2.2.3 叶长/宽的遗传比较 在不同杂交组合F1代的叶长/宽遗传中,超出母本的占了75.0%,超出父本的占62.5%,同时超出父、母本的占37.5%,因此叶长/宽表现出良好的超亲现象。其中,W2×W21、W2×M和W2×K的F1代的叶片长/宽均与双亲存在显著性差异(表6)。
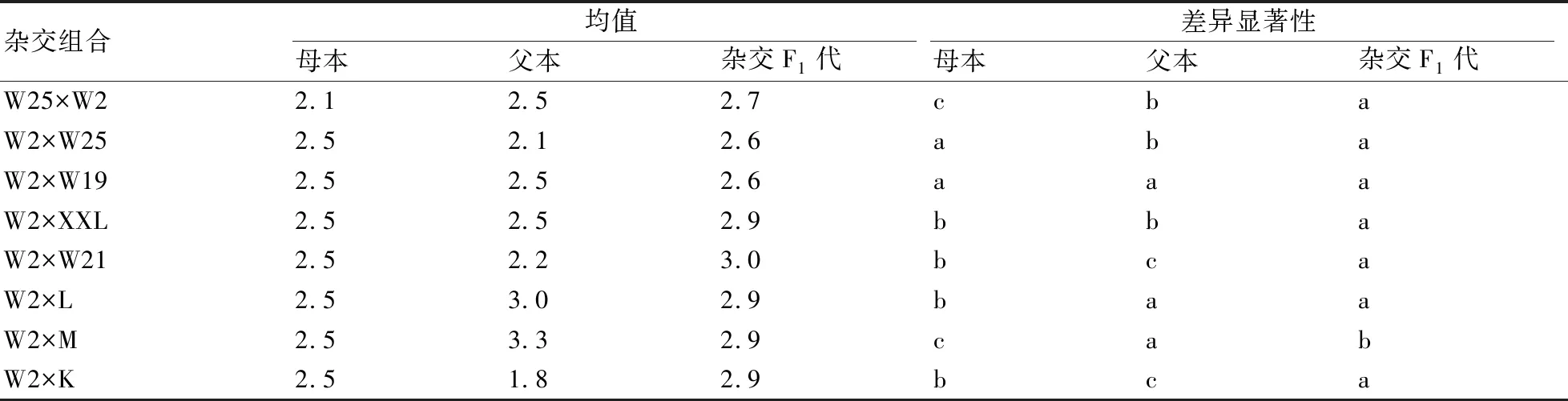
表6 不同杂交组合的F1代与其双亲的叶片长/宽差异性比较
2.2.4 叶柄长的遗传比较 在叶柄长的遗传中(表7),所有杂交组合F1代都与父本存在显著差异,而与母本存在显著差异的组合只有W2×L、W2×M和W2×K,仅占总组合数的37.5%,这说明叶柄长具有显著的遗传优势,而且此遗传来源于母本。
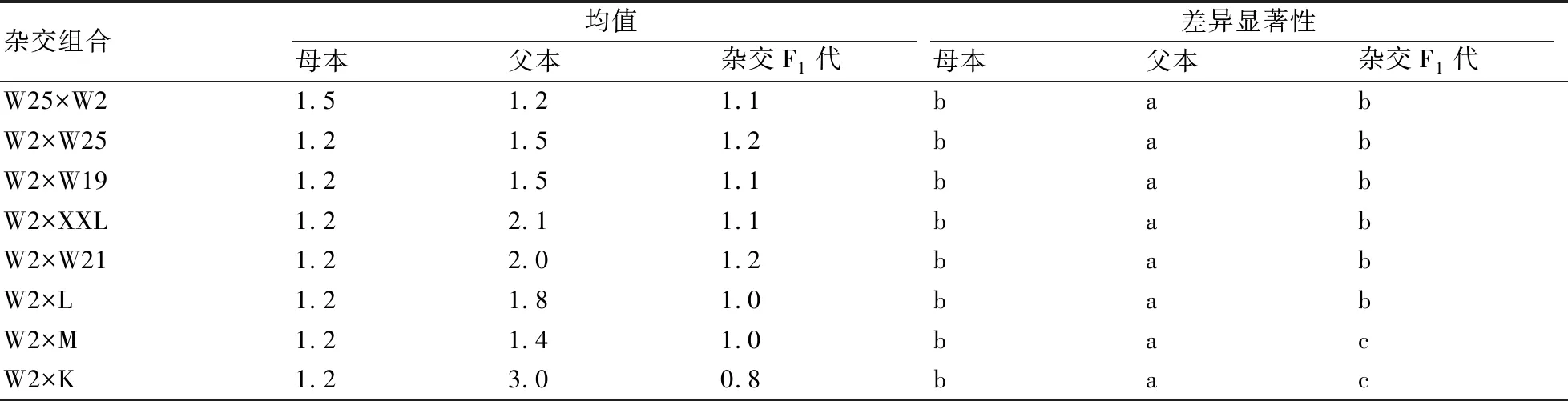
表7 不同杂交组合的F1代与其双亲的叶柄长差异性比较
3 讨论
杂交是转移优良性状或基因、获得植物新类型和选育有价值的新品种的有效方法之一,其在杜鹃花的育种中应用最为广泛[17]。抓到目标性状的遗传规律,找出变异元素,并将优质亲本进行合理的组合,从而提高育种效率,缩短育种年限,这对杂交育种来说具有重大意义。本试验对高山杜鹃不同杂交组合F1代的叶片性状遗传进行了分析,发现其叶片性状在遗传上表现出了一定的衰退趋势。类似的现象也出现在观赏荷花[20]和蝴蝶兰[21]上。分析其原因,可能是由于有性繁殖后的性状分离,致使杂种优势降低,故而导致后代性状平均值降低,最终导致衰退趋势的出现。但是,对于以缩小叶片为目标的育种来说,此衰退现象将非常有用;同时,对于一些观花、观果类植物育种而言,此遗传趋势也可以忽略。但是,在高山杜鹃F1代叶片性状衰退的同时,是否有其他观赏性状也发生了改变?这些都还有待于今后进一步的跟踪调查研究。
李娟娟等[19]在一串红F1代的观赏性状遗传研究中,将变异程度分为3个等级,即遗传变异度较小(0%~15%)、遗传变异度中等(15%~30%)、遗传变异度较大(>30%),认为变异度只要达到了中等即可认定该性状有较丰富的选择潜力。在本次试验中,高山杜鹃F1代的变异度都达到了中等,其中W2×K、W2×W25和W2×L的F1代叶柄长的变异度超过了30%,因此叶柄长这个性状存在较大的变异。但是,由于本次试验仅对各杂交组合F1代的较少植株(30株)进行了调查,在样本量上存在部分局限,因此今后的调查研究还需要加大样本量,以便能更准确地获得高山杜鹃叶片的变异情况,从而提高育种效率,得到目标性状。
4 结论
本试验结果表明:在高山杜鹃叶片性状的遗传中,叶片长的遗传最为稳定,叶柄长的变异度最大;从亲本的遗传性看,F1代更倾向于母本遗传;在各杂交组合中,‘安娜’ב门铃’杂交F1代表现出了很好的超亲现象,‘安娜’ב莫泽之栗’次之,其余6个组合表现为较大的衰退趋势,且‘安娜’ב宽杯杜鹃’杂交F1代的叶片各性状皆与双亲存在显著性差异。总体来说,要想利用杂种优势培育出综合性状优良的高山杜鹃后代还是有可能的。
猜你喜欢
作文·小学中高年级(2022年2期)2022-06-23
小天使·四年级语数英综合(2022年6期)2022-06-21
家教世界·创新阅读(2022年3期)2022-04-12
作文·小学低年级(2021年4期)2021-11-02
小学生学习指导(低年级)(2021年9期)2021-10-14
心声歌刊(2021年3期)2021-08-05
小聪仔(幼儿版)(2020年12期)2020-02-01
小学生学习指导(低年级)(2019年9期)2019-09-25
学生导报·东方少年(2019年27期)2019-01-14
小学生学习指导(低年级)(2018年9期)2018-09-26
