
云南省蓝舌病病毒“历史毒株”的遗传特征
2020-08-07 10:47
中国动物检疫 2020年8期
(云南省畜牧兽医科学院,云南省热带亚热带动物病毒病重点实验室,云南昆明 650224)
蓝舌病(bluetongue,BT)是一种由蓝舌病病毒(bluetongue virus,BTV)感染引起的烈性出血性动物疫病。BTV 主要通过库蠓属(Culicoidesspp.)雌性昆虫的吸血叮咬传播,能够感染牛、羊、骆驼和鹿等多种家养及野生反刍动物[1]。BTV感染绵羊后可引起较高的发病率和病死率[2-4]。而牛和山羊感染BTV 后往往无明显的临床症状,在BTV 传播过程中扮演了病毒储存库角色[3-4]。据统计,BT 对全球畜牧业生产每年造成约30 亿美元的经济损失,因此被世界动物卫生组织(OIE)列为须通报动物疫病[5-6]。
BTV 属于呼肠孤病毒科(Reoviridae)环状病毒属(Orbivirus),其基因组由10 条(genomic segment 1~10,Seg-1~10) 双 链RNA(doublestranded RNA,dsRNA)片段组成,可编码7 种结构蛋白(VP1~7)和5 种非结构蛋白(NS1~4 以及NS3a)[7]。Seg-2/VP2 是BTV 变异程度最高的成分。VP2 能够诱导被感染动物产生特异性中和抗体,对病毒血清型(serotype)具有决定性作用[7-8]。2008年至今,世界范围新发现了4 种BTV 新血清型(BTV-25~28)。因此,BTV 血清型由过去的24 种增加至28 种(BTV-1~28)[9-12]。
虽然VP3 序列在BTV 毒株间高度保守,但编码该蛋白的Seg-3 序列随BTV 毒株流行地域不同而出现差异,因此可以根据其序列差异,将世界范围内BTV 毒株划分为东方(Eastern)和西方(Western)两种主要的地域型(topotype)[13-14]。与VP3 类似,VP7 和NS3 蛋白的保守程度也较高,其编码节段Seg-7 和Seg-10 的序列变异也与BTV毒株流行地域密切相关,可根据Seg-7 和Seg-10的序列差异,将世界范围内BTV 毒株分为东方和西方两种主要的地域型以及数种地域亚型[13]。Seg-3、-7 和-10 序列具有强烈的地域特征,因此进行上述基因组节段序列分析,能够为追溯病毒起源提供依据[13-15]。
我国1979 年首次在云南省师宗县绵羊中记录了BT 的暴发[16]。之后,湖北省、安徽省、四川省和山西省相继暴发BT,并陆续分离到BTV-1、-2、-3、-4、-12、-15 和-16 型毒株,其中BTV-1和BTV-16 型在我国流行广泛,并且都能够导致绵羊出现较明显的临床症状[17-19]。2012 年以来,在国家公益性行业(农业)专项资助下,云南省畜牧兽医科学院在云南省、广西壮族自治区、广东省、湖北省、江苏省、山西省、新疆维吾尔自治区和内蒙古自治区等8 个省份设立哨兵动物,共分离到12 种血清型BTV 毒株(BTV-1、-2、-3、-4、-5、-7、-9、-12、-15、-16、-21 和-24)[20-23]。
多种血清型BTV 在我国广泛流行,不同血清型毒株间缺乏交叉保护,病毒通过突变和基因重配,不断产生变异毒株[17,20,22],这些都为当前的BT 防治工作带来了挑战。对我国不同历史时期的BTV 流行毒株进行序列比较分析,有助于掌握BTV 的演化特征以及我国 BTV 流行毒株的溯源分析。虽然本实验室已经完成了2012 年以来我国不同血清型BTV 分离毒株的全基因组测序,但对1995—1997 年云南省BTV 流行毒株的遗传特征尚不清楚。因此,对1995—1997 年分离自云南省分属7 种血清型的25 株BTV 毒株的Seg-2、-3、-7 和-10基因节段进行了扩增、测序和遗传特征分析,以期为我国BTV 的演化分析与溯源研究提供科学数据。
1 材料与方法
1.1 主要试剂
胎牛血清(fetal bovine serum,FBS)和minimum Eagle's medium(MEM) 培养基,购自Thermo Scientific 公司;一步法RT-PCR 试剂盒HiScript ⅡOne Step RT-PCR Kit(Dye Plus),购自南京诺唯赞生物科技有限公司;病毒基因组DNA/RNA 提取试剂盒和Universal DNA 胶回收纯化试剂盒,购自北京天根生化科技有限公司。
1.2 细胞、毒株和病毒培养
幼仓鼠肾细胞(baby hamster kidney cells,BHK-21),由本实验保存;25 株BTV 毒株,1995—1997 年分离自云南省师宗、芒市、景洪、峨山等地设立的哨兵牛,病毒分离信息见表1。将分离的BTV 毒株接种BHK-21 细胞,当细胞病变(cytopathic effect,CPE)达到90%时刮下细胞,4 ℃ 8 000 r/min 离心15 min,弃去沉淀,将病毒液分装,-80 ℃保存备用。
1.3 引物设计与合成
根据GenBank 中公布的BTV 毒株序列,采用Oligo 7.0 软件设计10 对引物,分别用于扩增7 种BTV 血清型毒株的Seg-2、-3、-7 和-10(表2)。引物委托上海捷瑞公司合成,以聚碳酸二乙酯(diethyl pyrocarbonate,DEPC)处理水稀释至终浓度为10 μmol/L 备用。
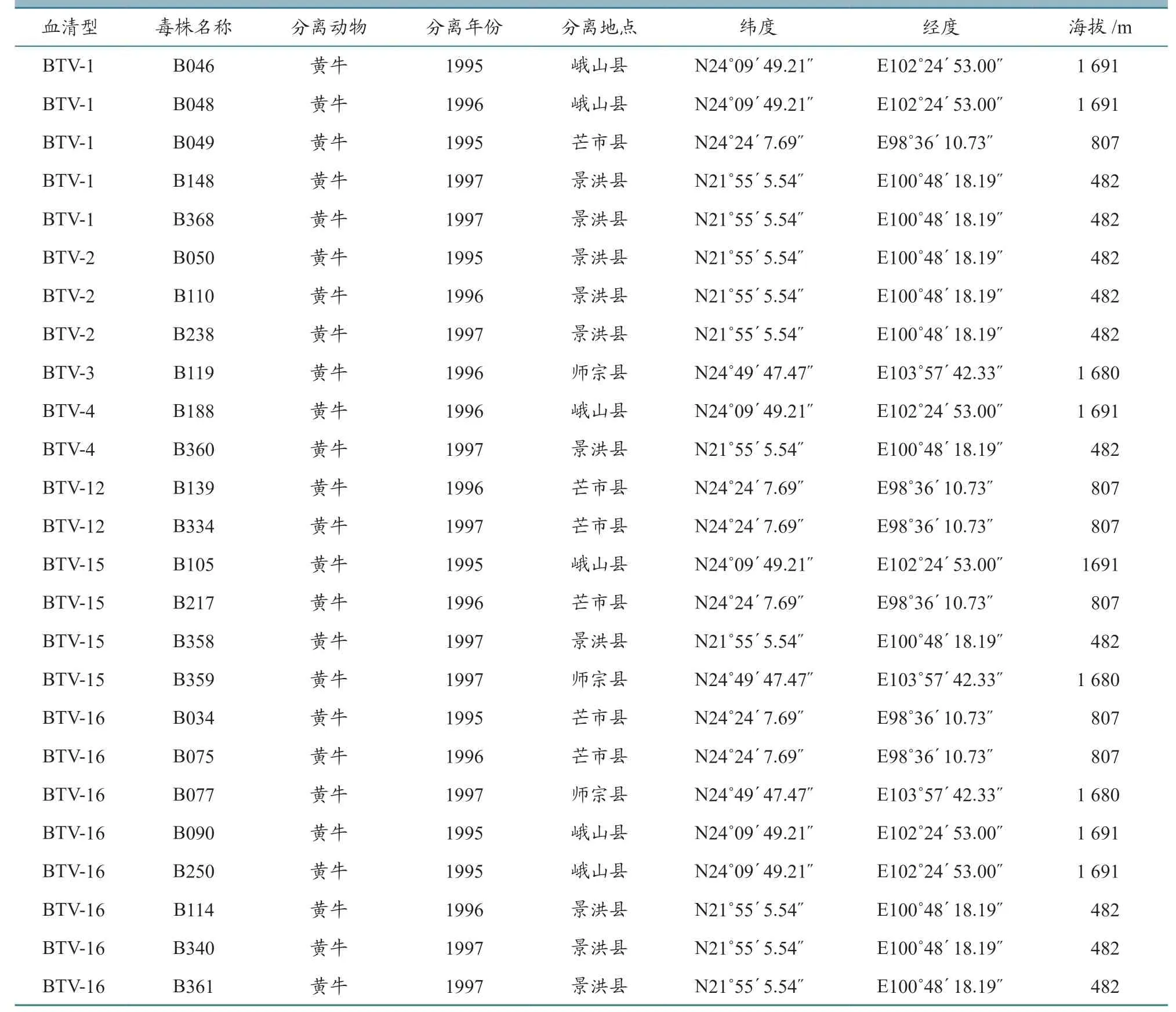
表1 1995—1997 年云南省BTV 分离毒株信息
1.4 RT-PCR 扩增与测序
采用病毒基因组DNA/RNA 提取试剂盒抽提病毒核酸,将核酸于94 ℃变性3 min 后,立即冰浴。分别取5 μL变性核酸为模板,使用表2中的引物对,采用一步法RT-PCR,对BTV 分离毒株的Seg-2、-3、-7 和-10 进行扩增。反应体系如下:2×One Step Mix(Dye Plus)25 μL,One Step Enzyme Mix 2.5 μL,上下游引物(10 μmol/L)各2 μL,核酸模板5 μL,以无RNA酶水补足至50 μL。反应条件如下:50 ℃ 30 min,94 ℃ 3 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1.5 min,35 个循环。利用Universal DNA 胶回收纯化试剂盒,对PCR 产物进行电泳胶回收,以PCR 扩增的上、下游引物作为测序引物,对纯化的DNA 进行双向测序。
1.5 序列分析与系统发生树构建
使用DNAstar 软件(Ver 6.0),对测序结果进行序列拼接组装;利用MEGA 6.0 软件[24],进行序列比对分析。采用最大似然法(maximum likelihood,ML)构建系统发生树:Seg-2 序列选择“GTR+G”模型,Seg-3 序列选择“TN93+G+I”模型,Seg-7 序列选择“GTR+G+I”模型,Seg-10序列选择“T92+G”模型,自举检验(bootstrap)取值1 000。序列相似度,以平均数(mean)表示。本研究中获取的序列以黑色菱形符号表示。系统发生树中其他国家分离的BTV 毒株序列以“GenBank序列号_BTV 血清型_分离国家_病毒分离时间”表示。
2 结果与分析
2.1 Seg-2、-3、-7 和-10 的RT-PCR 扩增
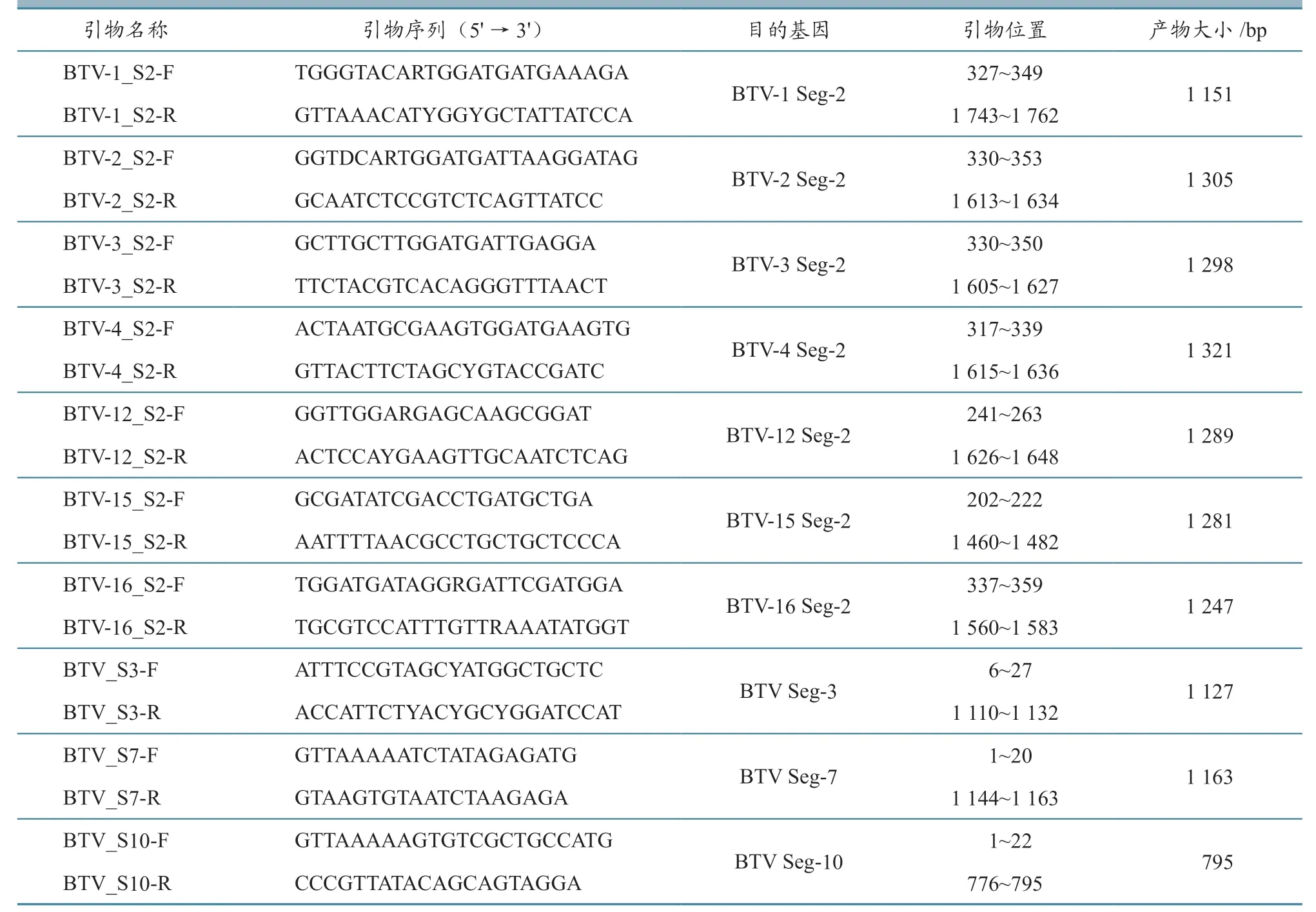
表2 云南省BTV 分离毒株Seg-2、-3、-7 和-10 的RT-PCR 扩增引物信息
设计10 对引物(表2),对分离的25 株BTV 毒株Seg-2、-3、-7 和-10 进行一步法RTPCR 扩增。结果显示,7 对针对不同血清型Seg-2的引物均能够特异性地扩增出大小约为1 200 bp,与预期大小相符的DNA 片段,而针对Seg-3、-7和-10 的3 对引物,也能够特异性扩增出大小分别为1 100、1 100 和800 bp,与预期大小相符的目的DNA 片段。
2.2 Seg-2 序列分析
序列比对分析显示,云南省1995—1997 年分离的25 株BTV 毒株分属BTV-1、-2、-3、-4、-12、-15 和-16 型等7 种血清型。世界范围内27种血清型(BTV-1~27)BTV 毒株的Seg-2 可分为12 种基因型(Nucleotype A—L)[13,15]。在系统发生树上,本研究中的25 株BTV Seg-2 与对应血清型BTV 参考毒株聚为一簇,分属A、B、H、I、G 和J等6 种基因型,与对应血清型BTV 国际标准参考毒株的Seg-2 序列相似度大于68.10%,VP2 序列相似度大于75.00%(图1~2),表明早期通过血清中和试验对BTV 血清型鉴定结果的正确性。云南省分离的同一种血清型BTV 的Seg-2/VP2 序列高度相似,Seg-2 核苷酸序列相似度在92.80%~100%之间,VP2 氨基酸序列相似度在96.90%~100%之间(图2),表明云南省流行的同一血清型BTV 的Seg-2 有着最近的共有祖先。
研究发现,同种血清型BTV 毒株Seg-2 之间,其序列也存在较大差异,并可根据这种差异,将同一血清型BTV 的Seg-2 分为东方地域型(Eastern topotype) 和西方地域型(Western topotype)[13,15]。构建的系统发生树显示,云南省分离的2 株BTV-12 型毒株(B139 和B334)的Seg-2 与南非BTV-12 型毒株聚为一簇,序列相似度大于98.10%,属于Western 地域型。云南省分离的BTV-1、-2、-3、-4、-15 和-16 型等毒株的Seg-2 与分离自日本、印度和澳大利亚的同血清型毒株聚为Eastern 地域型,其Seg-2 序列相似度在89.03%~95.46%之间,VP2 序列相似度在90.92%~98.30%之间;与同血清型Western 地域型毒株的Seg-2 序列相似度在68.00%~95.81%之间,VP2 序列相似度在75.28%~97.14%之间(图1~2)。
2.3 Seg-3 序列分析
Seg-3 序列比对分析显示,本研究的25株BTV 毒株间Seg-3 核苷酸序列相似度在79.35%~93.93%之间,VP3 氨基酸序列相似度在96.83%~99.95% 之间(图2)。构建的Seg-3 系统发生树显示,云南省分离的25 株BTV 毒株与来自中国台湾、澳大利亚和印度等地的毒株一同构成Eastern 地域型(图3)。本研究中的25 株BTV 与国外分离的Eastern 地域型、Western 地域型和新血清型(BTV-25~28)毒株间Seg-3 序列相似度在74.63%~89.29%之间,VP3 序列相似度在88.69%~99.12%之间(图2)。在Eastern地域型内部,4 株云南省分离的BTV-15 型毒株与1 株澳大利亚的BTV-15 型毒株聚为相对独立的一簇,构成远东地域亚型(Far Eastern topotype)(图3)[15],与Eastern 地域型其他毒株以及Western 地域型、新血清型(BTV-25~28)毒株间的Seg-3/VP3 序列相似度在75.18%~79.60%/88.20%~96.83%之间(图2)。这一方面表明云南省与澳大利亚的BTV-15 型毒株Seg-3 有着共同的起源,另一方面也表明该基因节段在进化上的相对独立性。
2.4 Seg-7 序列分析
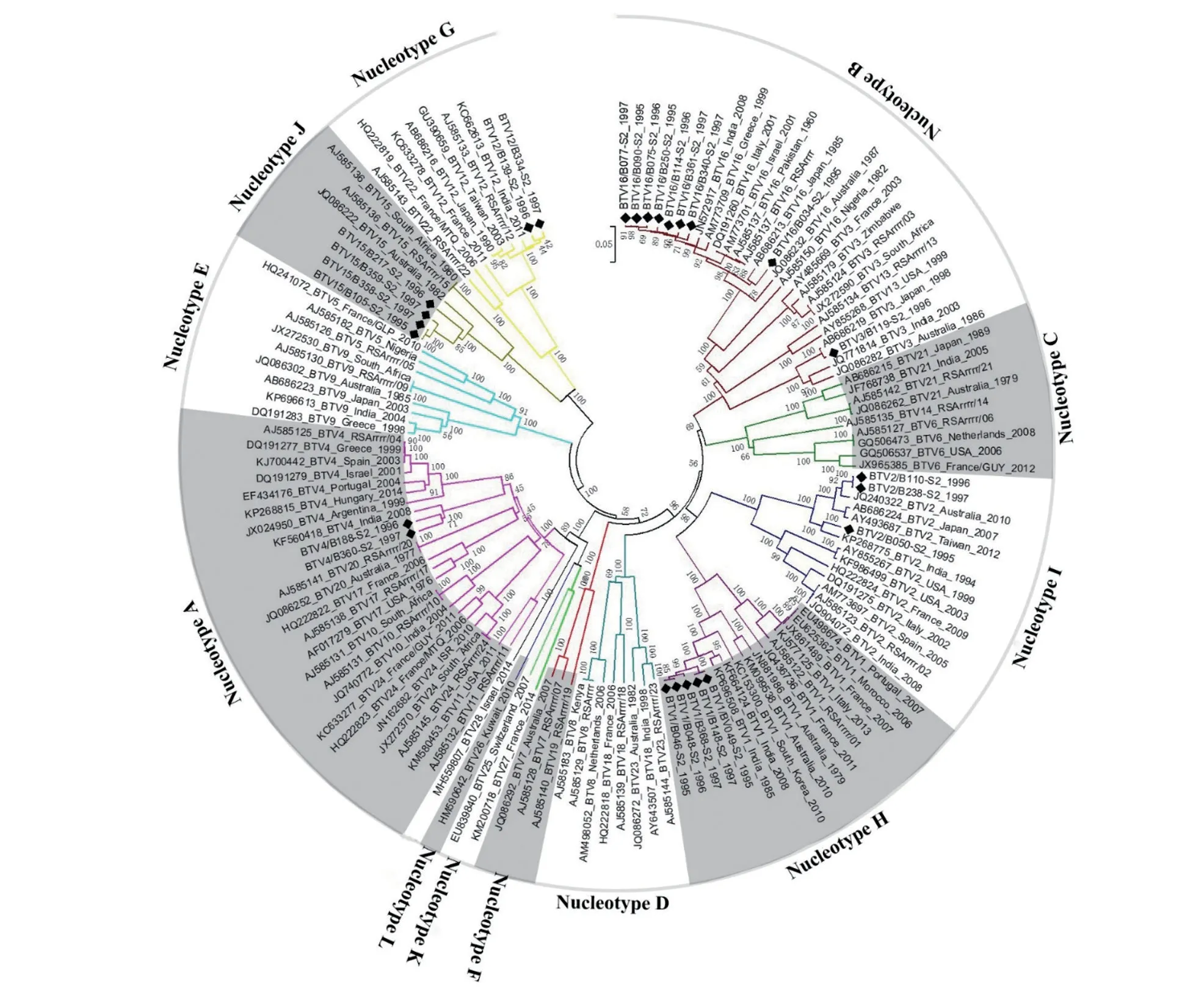
图1 1995—1997 年云南省BTV 分离毒株Seg-2 系统发生树
云南省不同血清型BTV 毒株间Seg-7 核苷酸序列相似度在73.85%~94.36%之间,VP7 氨基酸序列相似度在85.96%~99.35%之间(图2)。Maan等[13]的研究显示,BTV 的Seg-7 可分为3 种东方地域亚型(Eastern topotype 1~3)和7 种西方地域亚型(Western topotype 1~7)。云南省分离的21 株BTV 毒株的Seg-7 分属Eastern topotype 1~3地域亚型,分别与来自中国台湾、日本、印度和澳大利亚等地毒株的Seg-7 享有最近的亲缘关系(图4)。值得注意的是,云南省4 株BTV 的Seg-7 属于Western 地域型:云南省BTV-2 型毒株(B238)和BTV-12 型毒株(B139 和B334) 的Seg-7 属于Western topotype 1 地域亚型,与南非毒株具有最近的亲缘关系[13],序列相似度大于98.60%;BTV-16 型(B034)毒株为Western topotype 3 地域亚型,与荷兰毒株(GQ506478)和南非毒株(GQ506512)具有最近的亲缘关系,序列相似度均为95.60%(图4)。上述4 株BTV 毒株的Seg-7 序列相似度在78.90%~100%之间,VP7 序列相似度在94.60%~100%之间,而与云南省分离的Eastern 地域型毒株的Seg-7/VP7 序列相似度分别在78.33%~79.07%/94.46%~95.74%之间(图2)。
2.5 Seg-10 序列分析
云南省分离的25 株BTV 毒株的Seg-10 核苷酸序列相似度在86.61%~94.68%之间,NS3 氨基酸序列相似度在95.00%~99.08%之间(图2)。BTV 毒株Seg-10 系统发生树显示,该基因节段序列被划分为2 个东方地域亚型(Eastern topotype 1~2) 和3 个西方地域亚型(Western topotype 1~3)(图5)[13]。云南省分离的22 株BTV 毒株的Seg-10 均属于Eastern topotype 1 地域亚型,分别与来自中国台湾、日本、印度和澳大利亚等地毒株的Seg-10 享有最近的亲缘关系(图5)。而3 株云南省分离的BTV-15 型毒株(B105、B358和B359)单独构成Eastern topotype 2 地域亚型,与其他地域亚型毒株间Seg-10/NS3 序列相似度则在76.93%~85.81%/83.41%~97.26% 之间(图2、图5)。本研究中的25 株BTV 与Eastern topotype 1 地域亚型内其他毒株、Eastern topotype 2 地域亚型和Western 地域型亚型毒株间的Seg-10 序列相似度在77.02%~91.37%之间,NS3 序列相似度在83.23%~97.55%之间(图2、图5)。

图2 1995—1997 年云南省BTV 分离毒株核苷酸/氨基酸序列相似度
3 讨论
云南省复杂的地理环境和气候条件造就了该地区动物、植物和病毒的天然多样性。云南省地处我国边境地区,与越南、老挝和缅甸等多国接壤,家畜及畜产品贸易频繁,动物疫病跨境传播风险较高。多种血清型BTV 在云南省广泛流行,说明该地区在我国BT 防控中具有举足轻重的地位。过去认为BTV 主要流行于南纬35°至北纬40°之间。但是,随着气候变暖、库蠓活动范围扩大和潜在传播虫媒种类增多,BT 有逐渐向寒冷的高海拔和高纬度地区扩散的趋势[2,25-27],对我国北方地区的牛羊养殖业构成了威胁。掌握云南省1995—1997 年BTV 流行毒株的遗传背景,能够为分析我国BTV演化特征,进行病毒扩散与溯源分析提供宝贵的序列信息。
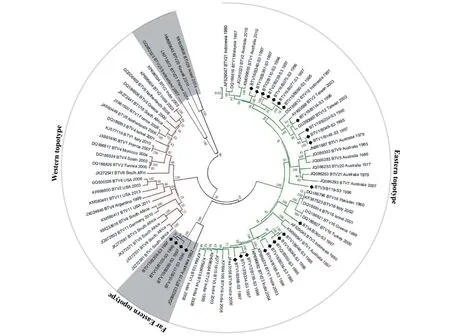
图3 1995—1997 年云南省BTV 分离毒株Seg-3 系统发生树
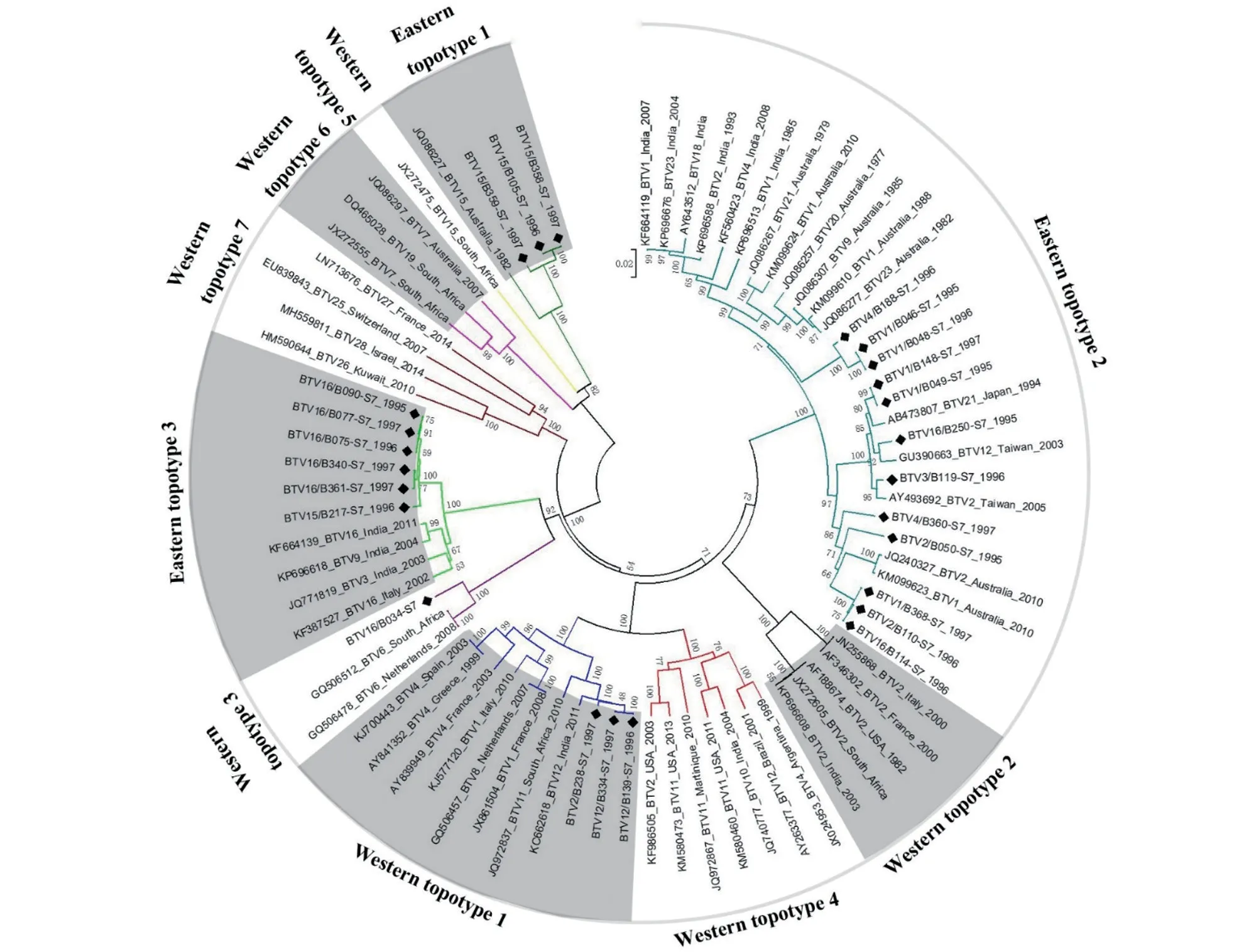
图4 1995—1997 年云南省BTV 分离毒株Seg-7 系统发生树
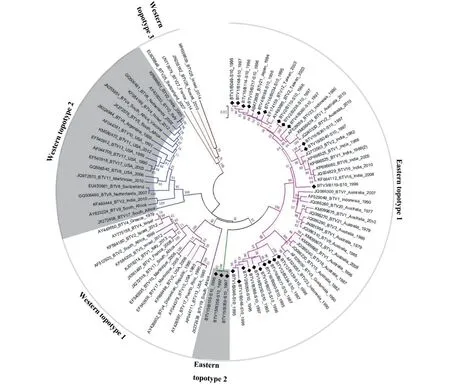
图5 1995—1997 年云南省BTV 分离毒株Seg-10 系统发生树
1995—1997 年,本实验室在云南省分离出7种血清型BTV(BTV-1、-2、-3、-4、-12、-15 和-16),2012 年至今,在云南省哨兵动物中分离出11 种血清型BTV(BTV-1、-2、-3、-4、-5、-9、-12、-15、-16、-21 和-24)[17-23],表明云南省流行的BTV 血清型具有多样性。BTV-5、-9、-21 和-24等4 种血清型BTV 在云南省的首次发现,提示我国可能还有其他血清型BTV 存在,因此对我国流行BTV 多样性的监测仍是一个长期的过程。病毒所处的生态环境不同,施加的选择压力不同,病毒的基因进化就会呈现出特定的地域特征。本研究发现,云南省流行的BTV 毒株序列也呈现出特有的地域特征:云南省流行的BTV-15 型毒株的Seg-3 形成了独立于Eastern 地域型和Western 地域型的进化分支(Far Eastern topotype),BTV-1和BTV-16 型毒株的Seg-2 也形成了独立的进化支系,BTV-15 型毒株的Seg-10 形成独立的Eastern topotype 2 地域亚型,揭示了云南省流行的BTV 在进化上的独特性。
Western 地域型BTV 毒株侵入云南省后,在传播过程中与我国本地流行毒株发生了基因重配,与同属环状病毒属的流行性出血病病毒(epizootic hemorrhagic disease virus,EHDV)某些血清型间Seg-7 较难发生重配不同[28-29],BTV 毒株的Seg-7片段能够在自然环境中发生相对自由的重配,提示BTV 进化可能主要由基因重配驱动,使病毒更适合在当地的生态系统中传播和生存[20]。
本研究通过分析1995—1997 年分离自云南省分属7 种血清型的25 株BTV 毒株Seg-2、-3、-7和-10 的遗传特征,发现云南省存在多种血清型BTV 流行,既有本地长期流行的BTV-1 型和BTV-16 型毒株,也有Western 地域型BTV 毒株侵入云南省后发生重配的毒株,提示我国BT 防控形势十分严峻。本研究为分析我国BTV 毒株的突变和基因重配提供了第一手资料,为追溯我国BTV 毒株来源提供了依据。
猜你喜欢
首都食品与医药(2022年21期)2022-12-06
动物医学进展(2022年9期)2022-11-26
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
医药前沿(2021年25期)2021-10-16
皮肤病与性病(2021年3期)2021-07-30
养猪(2020年6期)2021-01-27
中国兽药杂志(2019年4期)2019-05-15
江苏农业科学(2014年6期)2014-08-12
