
龙眼DlbZIP21基因克隆与表达分析
2020-08-28 08:57王金英张树伟潘介春彭宏祥范志毅赵德征李浩然
西南农业学报 2020年7期
王金英,张树伟,丁 峰,2*,潘介春*,彭宏祥,范志毅,赵德征,黄 幸,李 琳,王 颖,李浩然
(1.广西大学农学院,广西 南宁 530004;2.广西农业科学院/广西作物遗传改良生物技术重点开放实验室,广西 南宁 530007;3.广西农业科学院园艺研究所,广西 南宁 530007;4.广西宏华生物实业股份有限公司,广西 柳州 545000)
【研究意义】龙眼(DimocarpuslonganLour.)属无患子科(Sapindaceae)龙眼属植物,是一种重要的亚热带常绿果树,因其果实营养价值高及口感美味而具有很高的商业价值。在龙眼生长发育过程中,花芽分化受多种不良环境因素的影响,致使成花率降低,进而影响龙眼的产量和经济效益。此外,生产上种植的中熟龙眼品种较多,早、晚熟品种较少,品种结构不合理,龙眼收获期较集中,致使龙眼果实售价偏低,也直接影响果农的经济效益。碱性亮氨酸拉链(bZIP)转录因子可直接参与植物的成花调控,如bZIP转录因子FLOWERINGLOCUSD(FD)可与成花关键基因FLOWERINGLOCUST(FT)互作调控下游基因APETALA1(AP1)和LEAFY(LFY)的表达从而促进植物成花[1]。石硖龙眼一年只能开花结果一次,而四季蜜龙眼具有一年多次开花结果特性,但目前关于bZIP转录因子参与龙眼成花的调控机制尚不明确。因此,分析bZIP转录因子基因DlbZIP21在石硖和四季蜜两个龙眼品种不同组织和年周期表达水平的差异性,对揭示不同龙眼品种成花差异的分子调控机制及龙眼的熟期育种和产期调节具有重要意义。【前人研究进展】DlbZIP21基因为bZIP转录因子基因家族中的一员,具有bZIP转录因子的部分性质。bZIP转录因子在真核生物中广泛分布且相对保守[2],由一个碱性区域和一个bZIP结构构成,约包含20个氨基酸残基,碱性区域相对保守,能通过固定的核定位信号结构N-(X)7-R/K与DNA顺式原件特异性结合[3]。目前,已在多种植物中发现bZIP转录因子[2,4],如拟南芥、高梁、蓖麻、玉米和大豆。根据bZIP碱性结构域及其他保守结构域特点,可将拟南芥中的bZIP转录因子家族成员分为A、B、C、D、E、F、G、H、I和S共10个亚家族[3],不同亚族分别行使不同的功能[5],目前许多植物bZIP亚族的分类仍以此分类方法为依据。bZIP转录因子可参与植物组织分化、细胞延伸、能量代谢和蛋白调控等生物学进程。bZIP53蛋白能与启动子中的G-box元件特异性结合,同时与bZIP10或bZIP25相互作用,显著增强与DNA的结合活性,增加靶基因的转录水平[6]。OsbZIP39能通过蛋白水解转化为可溶性蛋白,从而转移到细胞核中,OsbZIP39DC能激活水稻原生质体结合蛋白1(BiP1)的启动子,并参与内质网应激反应[7],缺乏SC-uORF 59-先导区的转基因tbz17烟草植株叶片增厚,细胞增大,蔗糖含量升高[8]。Silveira等[9]研究发现,AtbZIP9转基因植物能使叶片出现发育缺陷,诱导其不正常发育。细胞分裂素(CTK)和茉莉酸(JA)能提高ATbZIP2基因的转录水平,增强其表达量[10]。此外,bZIP 转录因子能响应逆境胁迫,在胁迫条件下植物内源脱落酸合成量增加,从而激活bZIP家族基因表达[2]。在渗透胁迫处理下,MdbZIP48转基因拟南芥株系种子的发芽率、子叶绿化率和根长均优于野生型拟南芥[11]。Lee等[10]研究发现,盐处理能提高ATbZIP53基因的转录水平和表达量;AtbZIP2和AtbZIP11基因对非生物胁迫和激素也有不同响应。【本研究切入点】bZIP转录因子参与拟南芥等植物的成花调控,推测DlbZIP21基因可能参与龙眼成花调控,但目前关于bZIP转录因子对龙眼成花影响的研究未见报道。【拟解决发关键问题】对从石硖和四季蜜龙眼品种中克隆得到的bZIP转录因子基因DlbZIP21进行生物信息学分析,通过荧光定量PCR检测其在两个龙眼品种不同组织和年周期中的特异性表达情况,揭示不同龙眼品种成花差异的分子调控机制,为龙眼的熟期育种和产期调节提供理论依据。
1 材料与方法
1.1 试验材料
于2019年3月分别在广西大学龙眼试验园和果树标本园采集一年只开花一次的石硖龙眼和一年四季开花的四季蜜龙眼的根、茎、叶片、叶柄和花芽,在2019年5月分别采取转色期果皮、果肉和种子等不同组织材料,于2019年1-12月每月分别采集一次石硖和四季蜜龙眼叶片作为年周期材料。各材料洗净后放入采样袋中,液氮速冻后带回实验室,-80 ℃保存备用。
采用TIANGEN多糖多酚植物总RNA提取试剂盒提取龙眼总RNA,使用PrimeScriptTMRT reagent Kit with gDNA Eraser逆转录试剂盒进行cDNA链合成,采用PrimeSTAR®HS DNA Polymerase和ExTaq®进行基因克隆PCR检测,采用TaKaRa MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0试剂盒进行胶回收,采用TB GreenTMPremix ExTaqTMII试剂盒进行荧光定量PCR检测。
1.2 DlbZIP21基因克隆
采用1.0 %琼脂糖凝胶电泳检测提取的龙眼各组织总RNA纯度,利用逆转录试剂盒反转录合成cDN链合成。根据实验室经过3代转录组测序并结合龙眼基因组数据获得的bZIP基因序列,使用Primer Premier 5.0 设计DlbZIP21特异性引物(表1),第一阶段用高保真酶进行扩增,PCR反应体系25.00 μl:PrimeSTAR Max Premix 12.50 μl,ddH2O 9.50 μl,cDNA模板及上、下游引物各1.00 μl。扩增程序:94 ℃ 预变性3 min;98 ℃ 10 s,55 ℃ 15 s,72 ℃ 90 s,进行35个循环;72 ℃ 延伸10 min。以第一轮PCR扩增产物为模板,利用ExTaq酶进行第二轮PCR扩增,扩增体系25.00 μl:ExTaq0.15 μl,10×ExTaqBuffer 2.50 μl,dNTP Mixture 2.00 μl,模板及上、下游引物各1.00 μl,ddH2O 17.35 μl。扩增程序:98 ℃预变性4 min;98 ℃ 10 s,55 ℃ 30 s,72 ℃ 2 min,进行35个循环;72 ℃延伸10 min。PCR扩增产物检测纯化后克隆到pMD18-T载体进行测序。
1.3 生物信息学分析
利用NCBI在线分析DlbZIP21的开放阅读框(ORF),同时获得不同物种的bZIP氨基酸序列,应用BIOXM 2.6推测DlbZIP21氨基酸序列,使用ExPASy ProtParam在线工具分析DlbZIP21蛋白的理化性质,其跨膜螺旋区使用TMHMM进行预测,利用SignalP 4.1 Server在线进行信号肽预测,使用NCBI-CDD对DlbZIP21蛋白保守结构域进行预测分析,应用PSORT II Prediction在线预测亚细胞定位,使用NetPhos 3.1 Server分析DlbZIP21蛋白的磷酸化位点,利用在线分析工具SOPMA对DlbZIP21蛋白进行二级结构预测,用DNAMAN进行多序列比对,利用MEGA 6.0以邻近相接法(Neighbour-joining)步长值1000构建系统发育进化树。
1.4 DlbZIP21基因表达分析
采用LightCycler 480 System荧光定量PCR系统,在96孔板中进行荧光定量PCR反应。选用龙眼Actin作内参基因,根据测序所得的DlbZIP21基因序列设计定量引物(表1),分别以石硖和四季蜜龙眼的根、茎、叶片、叶柄、花芽、果皮、果肉、种子及全年12个月的叶片cDNA为模板,采用两步法进行荧光定量PCR扩增,反应体系20.00 μl:TB Green Premix ExTaqII 10.00 μl,上、下游引物各0.50 μl,cDNA模板1.00 μl,ddH2O 8.00 μl。扩增程序:95 ℃预变性30 s;95 ℃ 5 s,60 ℃ 1 min,进行40个循环;95 ℃融解5 s,60 ℃ 1 min,95 ℃ 0 s,50 ℃降温30 s。采用2-ΔΔCT法计算基因相对表达量[12],采用Excel 2010进行基因表达水平分析及制图。
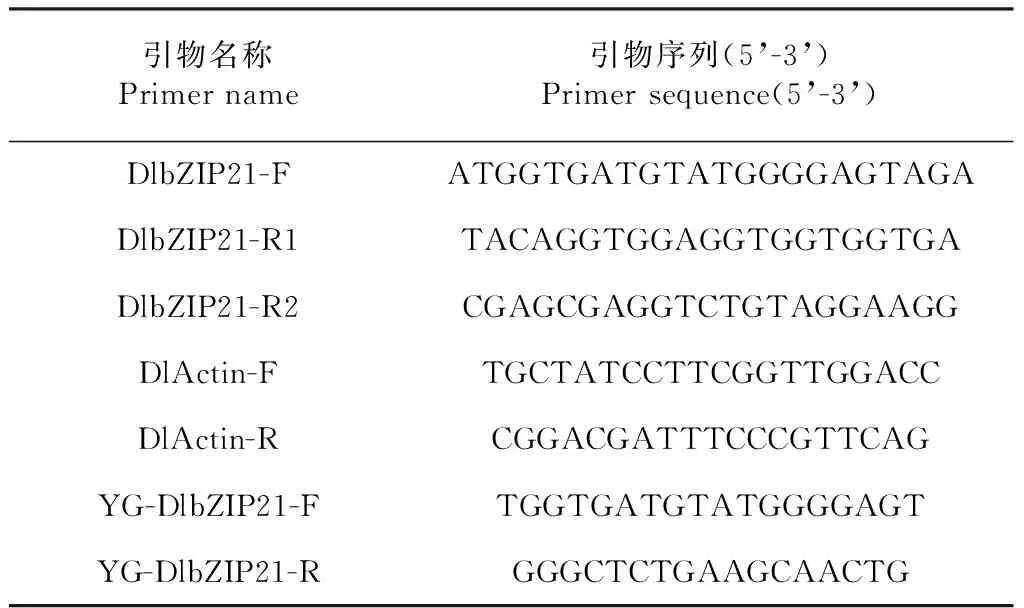
表1 引物序列
2 结果与分析
2.1 龙眼总RNA提取及DlbZIP21基因克隆
1.0 %琼脂糖凝胶电泳检测结果表明,使用总RNA提取试剂盒提取的龙眼组织RNA无降解且纯度较高(图1-A),可用于后续试验;根据特异性引物进行PCR扩增,目的基因为1353 bp,电泳检测扩增产物获得一条清晰明亮、与目的基因大小一致的条带(图1-B),符合后续试验要求,纯化后连接到pMD18-T载体,并转化大肠杆菌DH5α感受态细胞,通过筛选克隆测序,获得龙眼DlbZIP21基因序列信息。
2.2 DlbZIP21基因编码蛋白的理化性质
生物信息学分析结果表明,DlbZIP21-sx和DlbZIP21-sj基因的ORF均为1353 bp,各编码450个氨基酸残基;预测龙眼蛋白分子式分别为C2143H3433N647O695S13和C2144H3435N647O695S13,分子量分别为49.80和49.81 kD,等电点均为6.53;脂溶指数均为77.02,不稳定系数分别为62.44和62.71,属于不稳定蛋白;亲水指数分别为-0.544和-0.543,属于疏水蛋白;带正电残基总数(Arg+Lys)均为49,带负电荷残基总数(Asp+Glu)均为51;用在线分析工具PSORT II Prediction预测DlbZIP21均定位于细胞核;SignalP 4.1 Server在线预测DlbZIP21蛋白均无信号肽,利用TMHMM在线预测DlbZIP21蛋白均无跨膜结构区。由此推测DlbZIP21蛋白合成后不经转运,在细胞核中行使催化功能。
2.3 DlbZIP21蛋白保守结构域分析结果
以NCBI-CDS对2个DlbZIP21蛋白保守结构域进行在线预测,结果(图2)表明,两个DlbZIP21蛋白保守结构域相同,均属于bZIP蛋白超家族[13],具有调节细胞发育功能,该结构包括1个 bZIP-HBP-1b结构和1个DOG1结构域,DOG1结构域是一种特殊的植物种子休眠调节剂,具有bZIP家族典型结构特征。

M:Marker DL2000;1:石硖;2:四季蜜M:Marker DL2000;1:Shixia;2:Sijimi
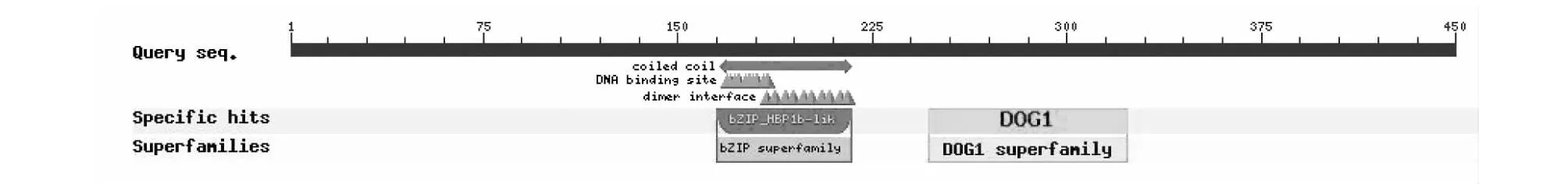
图2 DlbZIP21编码蛋白的保守功能域Fig.3 Conserved domain of the protein encoded by the DlbZIP21
2.4 DlbZIP21蛋白磷酸化位点分析结果
蛋白磷酸化位点分析结果(图3)显示,DlbZIP21-sx和DlbZIP21-sj蛋白均含有丝氨酸、苏氨酸和酪氨酸3种氨基酸磷酸化位点,其中,丝氨酸磷酸化位点分别为45和44个,苏氨酸磷酸化位点分别为14和15个,酪氨酸磷酸化位点均为2个;可能是潜在磷酸化位点的为第27位氨基酸,其阈值均为0.995;两个蛋白中丝氨酸的磷酸化位点最多。

A:DlbZIP21-sx磷酸位点预测;B:DlbZIP21-sj磷酸位点预测A:Phosphorylation site prediction of DlbZIP21-sx;B:Phosphorylation site prediction of DlbZIP21-sj
2.5 同源氨基酸序列比对分析结果
氨基酸序列比对分析结果(图4)表明,DlbZIP21-sx和DlbZIP21-sj蛋白的氨基酸序列与白羽扁豆(Lupinusalbus,KAE9602904.1)和大豆(Glycinemax,ABI34658.1)蛋白的氨基酸序列具有较高的一致性,相似性为72.69 %。
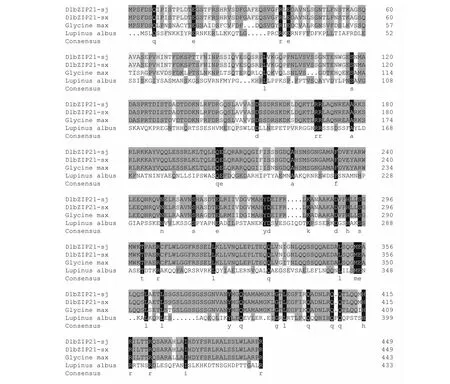
图4 DlbZIP21与其他物种bZIP的氨基酸序列比对分析结果Fig.4 Sequence alignments of amino acid sequence of DlbZIP21 compared with other species
2.6 DlbZIP21蛋白系统发育进化树分析结果
通过NCBI对DlbZIP21-sx和DlbZIP21-sj蛋白的氨基酸序列进行BLAST比对(图5),获得以下属于bZIP基因家族的成员,分别为连蕊茶(Camelliafraterna,QCQ29284.1)、茶树(Camelliasinensis,AGO05995.1)、可可豆(Theobromacacao,EOY04829.1)、拟南芥(Arabidopsisthaliana,NP_173381.1)、菜豆(Phaseolusvulgaris,AAK39130.1)、大豆(Glycine max,ABI34658.1)、水稻(OryzasativajaponicaGroup,ABA96884.1)、中华猕猴桃(Actinidiachinensisvar.chinensis,PSS29131.1)、土瓶草(Cephalotusfollicularis,GAV61199.1)、澳洲棉(Gossypiumaustrale,KAA3453989.1)、白羽扁豆(Lupinusalbus,KAE9602904.1)、糙叶山黄麻(Parasponiaandersonii,PON35707.1)、甜杏仁(Prunusdulcis,BBG96166.1)和榆树(Tremaorientale,PON58161.1)。进一步利用MEGA 6.0构建bZIP21蛋白的系统发育进化树,结果表明DlbZIP21-sx与DlbZIP21-sj蛋白的亲缘关系最近,同时与大豆、水稻和菜豆的亲缘关系较近,但与拟南芥的亲缘关系较远。

图5 基于bZIP21氨基酸序列相似性构建的系统发育进化树Fig.5 Phylogenetic tree analysis based on bZIP21 amino acid sequences homology
2.7 DlbZIP21基因表达水平分析结果
为了探讨DlbZIP21基因在石硖和四季蜜两个龙眼品种不同组织和年周期表达水平的差异性,分别采集两个龙眼品种的根、茎、叶片、叶柄、花芽、果皮、果肉和种子进行荧光定量检测。从图6可看出,在四季蜜龙眼各组织中均检测到DlbZIP21基因表达,其中在花芽中的相对表达量最高,在叶柄、茎和叶片中的相对表达量次之;同时,DlbZIP21基因在全年均有表达,其中在成花阶段相对表达量较高,在果实发育期相对表达量较低。在石硖龙眼中,检测到DlbZIP21基因在叶片中相对表达量最高,在茎和花芽中的相对表达量次之;同时DlbZIP21基因在全年均有表达,其中在成花阶段相对表达量较高,在果实发育阶段相对表达量较低。
从图6还可看出,在不同组织中,DlbZIP21基因在2个龙眼品种的果皮和果肉中表达量均偏低,在四季蜜龙眼花芽中的表达量较在石硖龙眼花芽表达量高5倍;在年周期中,从1-12月DlbZIP21基因在两个利用品种中的表达量总体上呈先上升后下降再上升的变化趋势,在四季蜜龙眼中的表达量普遍高于石硖龙眼,其中在花芽分化期和成花期(12月至翌年4月)的表达量高于石硖龙眼6~8倍;在果实采收后(8-10月)DlbZIP21基因在四季蜜龙眼中的表达量呈上升趋势,说明DlbZIP21基因在四季蜜龙眼成花方面发挥了重要作用,且可能参与诱导四季蜜龙眼一年多次成花。

图6 DlbZIP21基因在石硖和四季蜜龙眼中的不同表达水平分析结果Fig.6 Analysis results of different expression levels ofDlbZIP21 in Shixia and Sijimi
3 讨 论
植物bZIP转录因子基因家族参与植物成花调控已被学术界广泛认可。通过多年调查发现石硖龙眼一年只能开花结果一次,而四季蜜龙眼可一年多次开花结果,故克隆石硖和四季蜜龙眼品种的关键基因进行生物信息学分析及表达分析,旨在揭示龙眼的成花分子机制。已有研究表明,bZIP转录因子基因家族中的FD基因在植物花期开始前表达于茎尖分生组织(SAM)中,但不能单独诱导开花,FD基因与FT基因互作对FD基因促进成花具有重要作用[1,14]。此外,在光周期作用下可诱导FT基因转运到SAM,FT基因与FD基因互作,最终形成具有转录活性的FD-FT复合物,上调下游基因SOC1、AP1和LFY而促进成花[15]。目前,关于龙眼bZIP转录因子基因家族参与成花的研究相对较少,本研究克隆石硖和四季蜜龙眼DlbZIP21基因并分析其在龙眼不同组织和年周期的表达量,可为挖掘龙眼关键成花基因打下基础。
DlbZIP21基因属于bZIP转录因子基因家族成员,本研究克隆的DlbZIP21基因在石硖和四季蜜龙眼中的ORF均为1353 bp,编码450个氨基酸,两个品种具有相同的保守结构域;通过系统发育进化树分析发现,DlbZIP21基因与大豆、水稻和菜豆的亲缘关系较近,但与拟南芥的亲缘关系较远,表明DlbZIP21基因与豆科植物功能具有相似性,可为以后研究DlbZIP21基因的功能提供参考;进行氨基酸序列比对分析发现,不同植物间的bZIP转录因子基因其磷酸化位点也具有较高的一致性,通过对DlbZIP21蛋白编码的磷酸化位点进行预测,确定其磷酸化位点包括丝氨酸、苏氨酸和酪氨酸,与曹红利[16]对茶树CsbZIP的分析结果一致。已有研究发现,在拟南芥中,一些AtbZIP基因可调节ABRE启动子的顺式元件,包括ABF1、ABF2和ABF4,同时有研究表明,CsbZIP72和CsbZIP76含有bZIP转录因子启动子中识别位点的GCN_Motif顺式元件(G-box)[17],而本研究发现DlbZIP21蛋白存在一种特殊的植物种子休眠调节剂DOG1结构,能参与植物种子发育,此结构与上述研究结果相似,均可参与植物生物学进程。
本研究结果显示,DlbZIP21基因在四季蜜和石硖龙眼不同组织中均有表达,其中在四季蜜龙眼花芽中的相对表达量高出在石硖龙眼花芽中相对表达量的5倍;在年周期中,DlbZIP21基因在两个龙眼品种中的相对表达量总体上呈先上升后下降再上升的变化趋势,但在四季蜜龙眼中的相对表达量普遍高于石硖龙眼,在花芽分化期和成花期(12月至翌年4月),DlbZIP21基因在四季蜜龙眼中的相对表达量高于石硖龙眼6~8倍,由于四季蜜龙眼在果实采收后(8-10月)继续开花,同时DlbZIP21基因在四季蜜龙眼中的相对表达量也出现上升趋势,而一年开花一次的石硖龙眼相对表达量较低且无明显变化,与四季蜜龙眼具有一年多次开花结果特性相吻合[18],但具体作用机制尚需进一步探究。
4 结 论
DlbZIP21基因在四季蜜和石硖龙眼不同组织中均有表达,其中在花芽中的相对表达量四季蜜龙眼显著高于石硖龙眼,在叶片和种子中的相对表达量表现为石硖龙眼显著高于四季蜜龙眼;在年周期中,DlbZIP21基因在四季蜜龙眼中的相对表达量普遍高于石硖龙眼,其中在花芽分化期和成花期(12月至翌年4月)的相对表达量以四季蜜龙眼显著高于石硖龙眼,在果实采收后(8-10月)DlbZIP21基因在四季蜜龙眼中的相对表达量呈上升趋势。可见,DlbZIP21基因在诱导龙眼成花方面发挥重要作用,可能参与诱导四季蜜龙眼一年多次成花。
猜你喜欢
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2022年1期)2022-02-28
扬子江诗刊(2022年1期)2022-01-10
扬子江(2022年1期)2022-01-07
金山(2021年10期)2021-11-02
青年文学家(2020年22期)2020-08-31
作文大王·低年级(2019年4期)2019-05-13
世界热带农业信息(2018年5期)2018-11-09
世界热带农业信息(2018年6期)2018-03-05
分析化学(2017年12期)2017-12-25
