
昆明市东川区不同海拔高度水稻生长的差异性分析
2020-08-28 08:58吴永斌张茂松黄中艳
西南农业学报 2020年7期
吴永斌,朱 勇,朱 斌,张茂松,黄中艳
(1.云南省气象灾害防御技术中心,云南 昆明 650034;2.云南省气候中心,云南 昆明 650034;3.云南省气象学会,云南 昆明 650034)
【研究意义】地处低纬高原的云南境内,海拔高度变化大,地形地貌错综复杂,具有明显的高原立体气候特点[1]。地理气候的多样性孕育了丰富的水稻物种资源,已鉴定出6000多份水稻物种;加之云南四季太阳辐射强度差别不大[2],多数地区适种水稻且生长期长,云南已成为全国稻作生态及其遗传资源最具多样性的中心,从南部红河州河口县海拔76 m到西北部丽江市宁蒗县海拔2695 m都有水稻分布[3]。但是,云南不同地理气候区水稻的生长和产量差异显著,分析不同海拔高度种植地水稻生长的差异性及其成因,有助于深入认识水稻生长发育和产量形成的重要影响因素和影响机理。【前人研究进展】水稻生长的差异性与气候条件密切相关[4-5];夏季气温偏低、日照偏少,有效积温不足是较高海拔地区水稻生长的主要不利因素之一[6]。云南水稻冷害历史故有,2个亚种籼稻和粳稻对海拔高度的适应范围有差别,且不同生育阶段低温冷害特点和指标差异显著[7-8]。农业气象学者在水稻冷害方面做了大量研究[9],也有关于气象因素影响水稻生长和产量的分析[10-11]。比如,兰华雄等从光照、温度、降水和风速等方面揭示了云南涛源水稻高产的气象生态环境[12];邹丽云等分析了云南水稻产量对气候的敏感性及其灾害性天气[13];钟楚、孙秀芬等分析了气温与水稻的颖花量、秕谷率和空壳率、千粒重的密切关系[8,14]。【本研究切入点】根据以往研究,关于海拔或气象因素对水稻生长和产量的影响机制尚不够清晰。为此在昆明市东川区4个不同海拔高度上进行盆栽试验,分别观测籼稻和粳稻主要发育期茎鞘、叶干重、叶面积的差异,以及光合作用响应和产量性状指标的变化。【拟解决的关键问题】旨在分析水稻生长和产量性状随海拔高度变化的特点及其内在机理,探讨粳稻和籼稻在云南地区适宜种植的海拔范围,为云南水稻种植规划和气象为农服务提供科学依据和参考。
1 材料与方法
1.1 试验地点和栽培方式
在昆明市东川区选取4个不同海拔地点进行试验:新村1250 m(26°55′N,103°5.26′E)、乌龙1500 m(26°1′29′′N,103°5′44′′E)、半坡1800 m(26°4′33′′N,103°5′16′′E)、松毛棚2100 m(25°56′33′′N,103°06′35′′E),试验点间的最大距离约10 km。昆明市东川区内海拔高差悬殊,垂直气候差异大,具有典型的云南地理气候代表性。
根据当地水稻种植经验,籼稻适宜中低海拔地区、粳稻适宜中高海拔地区。据此选取内5优H25籼稻和滇杂31粳稻2个品种,进行盆栽试验[15],使用长30 cm×宽15 cm×高45 cm的塑料盆,籼稻毎盆栽3株,粳稻毎盆栽4株。一共种植730盆,具体栽种地分布为:新村(410盆籼稻)、乌龙(籼稻和粳稻各80盆)、半坡(籼稻和粳稻各40盆)、松毛棚(80盆粳稻);其中每个试验点分别选取30盆用于不同海拔比较试验,其余用于不同生育时期低温处理等试验。2016年3月10日统一育种;3月20日取土,供试土壤取自当地水稻和玉米轮作大田表层20 cm土壤,经晾晒、粉碎、过筛后,含水量均一致;4月19日装盆,每盆装土3 kg。4月24日插秧盆栽。5月6日对4个试验点以盆为单位施肥,施用碳酸氢铵6 g,5月18日再次施肥,用量相同,同时分别喷洒农药,用药为井·唑·多菌灵50 g,井冈霉素2 g。每个实验点均有人员定期看护,及时浇水。5-10月在水稻分蘖、拔节、孕穗、乳熟和成熟期分别采收样本、观测指标,10月底完成试验。
1.2 试验方法
1.2.1 不同海拔高度水稻生物量差异性试验 分别观测水稻主要生育期起始日期并调查各试验点的水稻生物量,分析不同海拔高度粳稻和籼稻生物量的差异显著性。在每个试验点随机取样3株,将叶片、茎鞘分开,分别测量茎鞘干重和叶片干重;采用称重法测量水稻叶面积,裁取中段等宽叶片10 cm,测量叶片宽度,计算其面积,再将叶片烘干称重,计算出单位面积叶片重量。每个试验点取样10盆水稻,分别统计每盆的穗数(包括有效和无效穗数),充分晒干后统计每盆水稻粒数(包括有效和空瘪粒数),计算不同海拔籼稻和粳稻的有效穗数和穗粒数(平均每穗的有效粒数)。
1.2.2 不同海拔高度水稻光合作用差异性试验 利用Li-6400XT便携式光合测定系统测定倒2叶(拔节期)或旗叶(孕穗和乳熟期)的光合作用光响应曲线和CO2响应曲线。光响应曲线测定初始斜率(表观量子效率)、最大净光合速率(Pmax)、光补偿点(Ic)和暗呼吸速率(Rd)、光饱和点(Isat)的变化,具体方法:设定光合有效辐射(PAR)梯度为2500、2000、1800、1500、1000、800、500、200、150、100、50、20、10、0 μmol/(m2·s),叶室温度粳稻为30 ℃,籼稻32 ℃,开放式气路,相对湿度维持在60 %~70 %,气体流量500 μmol/s;叶片先在PAR值为1800 μmol/(m2·s) 下适应20~30 min,然后按PAR梯度由高往低进行测量。CO2响应曲线测定在不同CO2浓度下的光合速率,具体方法:设定叶室CO2浓度梯度为2000、1700、1500、1200、1000、800、600、400、300、200、150、100、70、50 μmol/mol,叶室温度粳稻为30 ℃、籼稻32 ℃,PAR为1800 μmol/(m2·s);叶片先在CO2浓度为400 μmol/mol下适应20 min,然后按设定的CO2浓度梯度进行测量。每试验点(处理)选取3盆,每盆选取一个主茎的完全展开叶进行测量。
1.2.3 不同海拔试验地的气象观测 在4个试验点分别建立雨量、气温两要素自动气象站各一套,进行气象要素观测。与海拔梯度种植水稻试验相配合,2016年4-10月水稻生长季内全程观测记录降水量、气温数据,重点分析气温的变化和差异。
2 结果与分析
2.1 不同海拔试验地水稻主要发育期的差异
随海拔高度升高,水稻各主要发育期不同程度延后。4个海拔试验地的水稻在4月底至5月中旬相继进入分蘖期,6月初至6月中旬相继进入拔节期,7月初至8月中下旬处于孕穗期至乳熟期,9月下旬至10月中旬相继进入成熟期(表略)。同一品种不同海拔水稻分蘖出现日期相差2~5 d,拔节期相差3~5 d,孕穗期相差4~6 d,乳熟期相差5~7 d,成熟期相差6~12 d。无论粳稻还是籼稻,都表现为海拔越高,同一发育期出现越晚;越到后期,不同海拔高度上同一发育期相差越大。比如,籼稻孕穗期海拔1250 m为7月5日,海拔1500 m为7月9日,海拔1800 m为7月15日;粳稻成熟期海拔1500 m出现在9月26日,海拔2100 m则晚至10月18日。
2.2 不同海拔试验地气温和降水的差异
试验年新村气象站4-10月平均气温为23.7 ℃,比历年同期偏高0.2 ℃,其热量条件属正常年份。如表1所示,不同海拔高度气温差别显著,海拔越高气温越低。极端最高气温出现在海拔最低试验点新村站的6月16日,为38.6 ℃;极端最低气温出现在海拔最高试验点松毛棚站的4月7日,为0.9 ℃;海拔1250 m新村站 ≥10℃活动积温最高,为5 060 ℃·d,≥10 ℃活动积温随海拔高度增加而减少;水稻低温敏感期(孕穗至乳熟期)7-8月的最低气温也随海拔高度升高而降低。在水稻生长季内,累计降水量随海拔升高而增加,新村1250 m降水量最少,为523.3 mm。试验通过及时浇水灌溉,消除降水量的差异对各海拔水稻生长的影响。
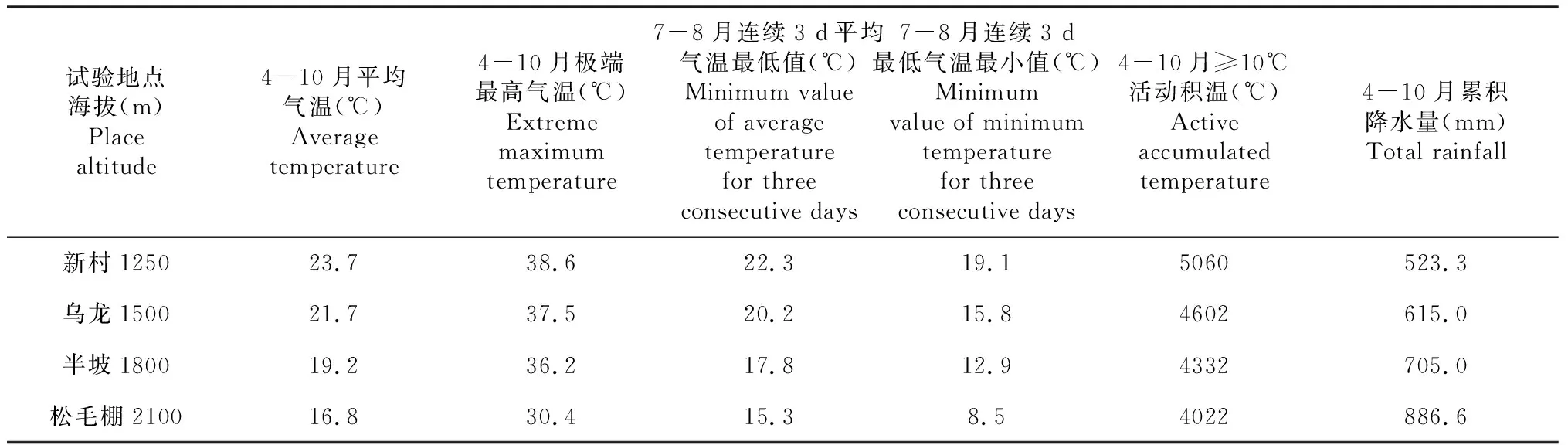
表1 不同海拔试验地水稻大田生长期气象观测数据
2.3 不同海拔试验地水稻茎鞘、叶干重和叶面积的差异
3个海拔试验地粳稻茎鞘干重均随时间呈上升趋势(图1),水稻拔节期和孕穗期时茎鞘干重随海拔高度的增加逐渐下降。水稻乳熟期时在海拔1800 m的水稻茎鞘干重最大,与2100 m的水稻差异显著。

图中同一生育期数据上不同小写字母表示差异达0.05显著性水平。下同Different small letters on data of the same growth period mean significant difference at 0.05 level.The same as below
3个海拔试验地粳稻叶片干重均随时间呈上升趋势,但在海拔1500 m的水稻叶片干重的增速较缓。在拔节期和孕穗期随海拔高度的增加,水稻叶片干重逐渐下降,孕穗期时1500和1800 m的显著高于2100 m的。但3个海拔高度乳熟期水稻叶片干重没有显著差异。
在海拔1800和2100 m的水稻叶面积随时间推移逐渐增大,在海拔1500 m的粳稻从拔节至孕穗期叶面积逐渐增加,孕穗至乳熟期呈下降趋势;在海拔1500 m的粳稻拔节至孕穗期叶面积要显著高于1800和2100 m的,但至乳熟期时海拔1500 m粳稻叶面积显著低于1800和2100 m的。
不同海拔高度籼稻茎鞘干重随时间变化具有明显差异(图2)。在海拔1250 m的籼稻分蘖至孕穗期茎鞘干重逐渐增加,但增加的速率越来越小,至乳熟期略有降低。在海拔1500 m的籼稻分蘖至孕穗期茎鞘干重都保持较快的增长,分蘖期显著低于1250 m的,孕穗期反而超出;但籼稻乳熟期茎鞘干重迅速降低,与1250 m的相差很小。海拔1800 m上籼稻茎鞘干重呈指数快速上升,在分蘖期、拔节期显著低于1250 m的,但乳熟期的茎鞘干重要显著高于1250和1500 m的。

图2 不同海拔试验地籼稻茎鞘、叶干重和叶面积随时间变化Fig.2 Changes of the dry matter of stem sheath and leaf,the leaf area of indica rice at different planting altitudes
在海拔1250和1500 m的籼稻叶片干重随时间的变化相似,分蘖至乳熟期上升,两者仅在拔节期差异显著,至乳熟期两者差异很小。在海拔1800 m的籼稻拔节至乳熟期叶片干重几乎呈直线增加,各生育期叶片干重都显著低于海拔1250 m的。3个海拔高度籼稻叶面积变化与叶片干重的变化类似,在1250和1500 m的叶面积较高且二者无显著差异。在海拔1800 m的籼稻分蘖期、拔节期叶面积显著低于1250和1500 m的,3个海拔高度的籼稻分蘖至孕穗期叶面积都逐渐增大,到乳熟期在1250 m海拔的明显下降。
研究表明,在较低海拔2种水稻营养生长阶段生长较充分,干物质积累较多,进入生殖生长后期有利于其干物质向籽粒转移;而在较高海拔区2种水稻因前中期气温较低而生长缓慢,以致进入生殖生长后期时其营养生长和干物质积累尚不充分。
2.4 不同海拔试验地籼稻光合作用的差异
2.4.1 不同海拔试验地籼稻光响应特征 由图3可见,在海拔高度较低的新村,净光合速率于拔节至孕穗期明显增加,于乳熟期快速降低;乌龙(1500 m)试验地水稻净光合速率随时间呈现明显逐渐降低趋势;半坡(1800 m)则表现为先快速降低,随后明显增加。而在相同的生育期内,随海拔高度的增加,水稻叶片净光合速率总体呈逐渐下降,尤其孕穗期表现最为明显。当光合有效辐射达2000 μmol/(m2·s)以上时,净光合速率逐渐接近饱和点;水稻最大光合饱和点在较低海拔的新村(1250 m)出现在孕穗期,海拔较高的另2个点出现在拔节期。说明气温是影响光合速率的重要因素,水稻进入到生殖生长阶段(孕穗期)若气温偏低(在海拔较高地区)会显著降低其光合效率。正是这个制约因素,造成较高海拔区水稻的干物重、叶面积一直到乳熟期都呈增加趋势(图2),表明由于生长前中期气温较低,拔节、孕穗期光合效率较低(图3),并造成旺盛生长期缩短,以致在中后期时营养生长和光合积累尚未充分,进而不利于后期形成较高经济产量。

图3 不同海拔试验地籼稻拔节期、孕穗期和乳熟期的光合响应曲线变化Fig.3 The photosynthetic response curves of indica rice at jointing,booting and milk-ripe stages at different testing altitudes
根据表2,随着海拔高度的增加,初始斜率(表观量子效率)由营养生长后期基本相同变为生殖生长期的逐渐减小,即海拔较高种植地的水稻在光照强度增大时其光合作用效率的敏感性下降;最大净光合速率(Pmax)随海拔高度升高,在拔节期先增大后减小、孕穗期逐渐减小、乳熟期逐渐增大;光补偿点(Ic)在拔节期略呈增大、孕穗期略有减小、乳熟期明显增大,表明较高海拔区水稻生殖生长期要求较高的光合照强度才能产生光合积累;光饱和点(Isat)在拔节期先略减小后增大、孕穗期逐渐增大、乳熟期明显变小,表明较高海拔区因气温强度不足,水稻旺盛生长峰期出现在孕穗抽穗期,至生殖生长后期水稻对光照利用率显著降低;暗呼吸速率(Rd)在拔节期逐渐增大、孕穗期逐渐减小、乳熟期逐渐增大,说明水稻夜间呼吸消耗能量较大的生长阶段低海拔地为水稻生长中期(孕穗抽穗期)、高海拔地为生殖生长后期(乳熟期),从而较高海拔地的水稻不利于形成较高产量。

表2 不同海拔试验地籼稻叶片对不同光照强度光合响应的特征参数
2.4.2 不同海拔试验地籼稻对CO2浓度的光合响应特征 由图4可见,除在海拔1500 m的乌龙,水稻叶片对CO2浓度光合响应变化各生育期都接近外,较低海拔的(1250 m)和较高海拔(1800 m)两地点水稻的光合作用对CO2浓度响应不同生育期均有较大差异。当CO2浓度低于200 μmol/mol时,各海拔高度处水稻光合响应相差较小,净光合速率增加的幅度基本相近。在海拔1250和1800 m,当CO2浓度高于300 μmol/mol以后,相同海拔高度下不同生育期的响应曲线明显不同;其中海拔较低的新村站不同生育期差异最为明显,表明较低海拔地带因气温较高,水稻在CO2浓度升高时,其光合效率提高的效果更明显。

图4 不同海拔试验地籼稻主要发育期对CO2浓度的光合响应曲线Fig.4 The photosynthetic response curves of rice to CO2 concentration at three developmental phases at different testing altitudes
在海拔1250 m,当CO2浓度达最大值(1000~1200 μmol/mol)时旺盛生长期(孕穗期)水稻光合效率开始出现明显下降,海拔1500 m的水稻光合速率为最大值时的CO2浓度拐点为1200~1500 μmol/mol,而海拔1800 m的水稻光合速率最大时CO2浓度拐点值为1200~1600 μmol/mol,即随海拔高度升高,水稻最大光合效率要求的CO2浓度有增大的趋势。海拔1800 m水稻的初始斜率最大,其次为1250和1500 m的最低。随着海拔高度的增加,光合能力逐渐减小,海拔1500~1800 m水稻对CO2浓度的光合响应较1250~1500 m海拔区间的变化要大。CO2饱和点随海拔高度的增加而增大,1500 m与1250 m之间相差较小。CO2补偿点以海拔1250 m 最高,其次为1800 m;呼吸速率在海拔1500 m的最低,其次为1250 m,在海拔1800 m明显较高(表3)。

表3 不同海拔试验地籼稻叶片对CO2浓度光合响应的特征参数
研究表明,与CO2浓度变化的影响相比,水稻光合速率(Amax)对海拔高度(气温)变化的敏感度更大,较高的温度很有利于提高光合速率; 同时,CO2饱和点和补偿点及光呼吸速率(Rp)随海拔变化特点,反映了在较高海拔区由于气温较低,水稻光合效率较低。乳熟期高海拔区水稻光合速率、CO2饱和点高于低海拔区同期水稻,是因为高海拔生长的水稻进入乳熟时营养生长仍很旺盛。
2.5 不同海拔试验地水稻产量的差异
如图5所示,籼稻总穗数随着海拔升高而变少,粳稻总穗数则随海拔升高还略有增多。籼稻的平均有效穗数和总穗数随着海拔高度的升高而减少。相比籼稻而言,粳稻的平均有效穗数和总穗数与海拔高度的关系较为复杂,总穗数在海拔1500 m以上随海拔升高还略增多,有效穗数在1500和1800 m较为近似;但到海拔2100 m,有效穗数急剧下降。

图5 水稻有效穗数随海拔高度的变化Fig.5 Changes of effective panicles’ number of rice with different planting altitudes
如图6所示,籼稻的总粒数随着海拔的升高逐渐减少。粳稻的总粒数则随海拔升高有增大的趋势。籼稻的穗粒数和总粒数都是海拔1250 m的最高,随着海拔的升高籼稻的穗粒数和总粒数逐渐减少。粳稻的穗粒数在1500和1800 m基本接近,但到海拔2100 m穗粒数急剧下降。
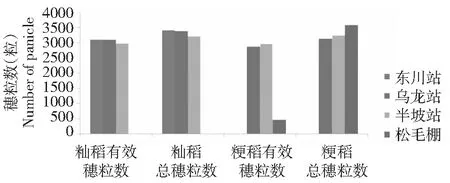
图6 水稻有效穗粒数随海拔高度的变化Fig.6 Changes of effective grains’ number per spike of rice with different planting altitudes
3 讨 论
(1)不同海拔高度水稻生长特性和经济产量性状的差异,造成水稻经济产量不同。水稻经济产量由单位面积有效穗数、平均穗粒数和千粒重决定[16]。随海拔高度升高,水稻生长越缓慢,水稻干物重、叶面积越小,有效穗数越小,生育期越延长(表1),低温冷害的影响越大(穗粒数越少),从而造成产量不同。
(2)气温是影响水稻光合速率和光合效率的主导因素。根据图3~4,低海拔区因气温较高,水稻光合速率大。高海拔区水稻拔节和孕穗期光饱和点高于中海拔和偏低海拔的水稻,同时乳熟期CO2饱和点和补偿点都较高且暗呼吸速率高,这些特点共同表明高海拔区水稻的光合效率显著偏低,以致生殖生长后期水稻营养生长积累仍不充分。这说明气温对光合速度和光合效率起主导性作用。
(3)气温变化的影响是不同海拔试验地水稻生长和产量性状差异的根本原因。试验地从海拔1250~2100 m,4-10月≥10 ℃积温从5060 ℃下降到4022 ℃,同时平均气温从23.7 ℃下降到16.7 ℃。意味着海拔每升高100 m,有效积温减少122 ℃·d,平均气温下降0.82 ℃。总热量的减少和热量强度的下降,造成海拔高度升高后水稻光合速率和光合效率下降,从而导致干物质积累量降低、生长缓慢(叶面积减小和发育期延长),以及水稻产量性状(有效穗数和穗粒数)变差。无论是籼稻还是粳稻,气温对光合效率、干物质积累和产量性状的影响是类似的。
(4)海拔升高会加大水稻低温冷害发生的机率和危害,低温是影响水稻产量的重要因素。水稻孕穗至乳熟期低温冷害是制约云南水稻产量水平的主要原因之一[8]。根据表2,试验当年7-8月连续3 d平均气温和平均最低气温的最低值在海拔1800 m的半坡分别为到17.8和12.9 ℃,两者都低于云南地区粳稻夏季低温冷害临界值[8]。因此,海拔1800 m的籼稻有效穗数、海拔2100 m的粳稻有效穗数和穗粒数分别显著下降、大幅度下降。有效穗数与有效分蘖密切相关[16],而穗粒数主要受空瘪率影响[11]。海拔2100 m的粳稻总穗数不降但有效穗数严重下降,主要是穗粒数不足造成。
4 结 论
(1)海拔1250 m种植的籼稻生长特征指标最优,在海拔1500 m种植的籼稻茎鞘干重、叶片干重和叶面积下降明显,有效穗数也受到影响;在海拔1800 m的籼稻生长和产量性状指标(尤其是有效穗数)下降幅度大。粳稻比籼稻有较大的海拔高度范围适应性。粳稻生长特征指标表现为海拔在1500 m的最优,海拔1800 m的粳稻生长指标下降明显而产量性状未受显著影响,海拔2100 m的粳稻两类指标均受到严重影响。
(2)随海拔高度的增加,因气温逐渐下降,水稻孕穗期叶片最大净光合速率和表观量子效率逐渐降低,光饱和点和CO2饱和点增大,即光合强度减弱;水稻生殖生长后期光饱和点减小、暗呼吸和光呼吸速率及光补偿点都增大,造成光合效率和光合积累量下降,从而不利于水稻生长发育和产量形成。水稻光合作用对种植地海拔高度变化的响应特点,与其生长特征和产量性状指标随海拔变化是一致的。
(3)随海拔高度升高,气温逐渐降低,水稻光合速率和效率下降,干物质积累减少,稻株营养生长变缓慢,有效分蘖减少(尤其籼稻); 甚至到生殖生长后期(乳熟期)水稻营养生长仍未充分,影响光合生成物转化为经济产量,导致穗粒数显著减少和籽粒饱满度不足(空瘪率增加),最终造成产量降低。
(4)不同海拔高度气象条件差异显著,气象条件变化对水稻生长的显著直接影响,表现为不同海拔高度对水稻生长的间接影响。粳稻、籼稻分别适宜生长于相对较低、较高的气温条件,水稻生育期内有效积温随海拔高度增加而变少,海拔高度引起的温度变化对两种水稻生长均影响显著。在昆明市东川区适宜籼稻和粳稻种植的海拔上限分别为1500 m以下和2000 m以下。
(5)较高海拔区水稻易受低温冷害影响,并造成水稻经济产量水平下降。低温对水稻的影响,籼稻首要表现是有效穗数(有效分蘖数)不足,粳稻则主要是有效籽粒数(穗粒数)下降。低温冷害是较高海拔区制约水稻种植适性和产量水平的主要不利因素。
猜你喜欢
安徽农学通报(2022年8期)2022-05-06
安徽农学通报(2022年6期)2022-04-07
新农民(2021年15期)2021-09-16
北京汽车(2021年1期)2021-03-04
江苏农业科学(2020年22期)2020-03-03
中国种业(2020年2期)2020-02-26
作文中学版(2018年11期)2018-11-29
现代农业科技(2018年14期)2018-10-31
意林·少年版(2018年11期)2018-07-04
世界热带农业信息(2014年3期)2014-08-12
