
桂南茶园土壤团聚体酶活性对植茶年限的响应
2020-11-12 07:52王晟强叶绍明
生态学报 2020年18期
王晟强, 张 喆, 叶绍明
广西大学林学院, 南宁 530004
广西地处我国南疆,雨水充沛,具有优越的植茶气候条件和悠久的制茶饮茶历史,是我国重要的茶叶产区之一。截止至2017年底,广西茶园面积已达7.33万hm2,茶叶产量7.00万t,毛茶产值超过45亿元[1]。在农民增收脱贫方面,茶产业的经济效益发挥着十分关键的作用。据不完全统计,广西有979个村、20.26万户种植茶树,其中有454个贫困村、4.96万贫困户种植茶树。广西80%以上的茶园分布在贫困县,茶产业已成为多数贫困县脱贫致富的支柱产业[2]。然而,在茶园生态系统中,由于茶园的施肥管理、茶树凋落物归还于土壤以及根系分泌物等原因,使得土壤有机碳及养分在植茶过程中发生一系列变化,从而影响茶树的生长发育和茶叶的产量品质[3-7]。目前,关于茶园土壤质量演变的研究集中在江南茶区和西南茶区,而在华南茶区尤其是广西茶园却鲜见报道。
土壤酶是生态系统物质循环和能量流动中最为活跃的生物活性物质,其种类繁多,参与着复杂多样的生物化学反应[8-9]。在土壤碳素循环过程中,β-葡萄糖苷酶和蔗糖酶起着关键作用[10-11],它们可将土壤大分子糖类水解成为活性相对较高的小分子糖类,深刻影响着土壤有机碳的分解与转化[12-13]。作为参与土壤氮素循环常见的水解酶,脲酶能够酶促尿素水解生产氨,蛋白酶可将蛋白质肽链水解生产氨基酸,从而促使土壤有机氮向植物有效态转化[14-15]。土壤有机磷的转化主要依靠磷酸酶来完成,磷酸酶能够有效水解土壤有机磷化合物并生成可直接被植物吸收利用的无机磷,其活性可表征土壤磷素循环状况[16]。可见,通过土壤酶活性来间接反映或预测土壤有机碳及养分利用效率以及土壤质量演变具有可行性。
土壤团聚体是形成良好土壤结构的物质基础[17-18],不同粒径团聚体不仅决定着土壤孔隙分布、数量搭配、形态特征以及土壤对外界应力的敏感性,而且在土壤有机碳及养分保持、供应及转化等方面发挥着不同作用[19-20]。了解土壤酶在团聚体中的活性差异,对调控和提升土壤有机碳及养分利用效率具有重要意义。前期研究发现,土壤有机碳及养分在团聚体中的分布存在明显差异,且随植茶年限变化显著。基于此,本研究以我国四大茶市之一的“西南茶城”——南宁市横县为研究区域,立足于土壤团聚体角度,旨在揭示土壤碳氮磷循环相关酶活性对植茶年限的响应机制,以期为广西茶园提升土壤肥力、保障土壤健康以及促进土壤资源可持续利用奠定理论基础。
1 材料与方法
1.1 研究区域概况
研究区域位于东经108°48′—109°37′和北纬22°08′—23°30′,属亚热带季风气候区。年均温21.6 ℃,极端最高气温40.4 ℃,极端最低气温-2.4 ℃。年均降雨量约1304 mm,全年79.1%的降雨量发生在4—9月期间。地貌以丘陵台地为主,土壤类型为赤红壤。该区域植茶历史悠久且以国家地理标志保护产品南山白毛茶为主,从而形成了具有一定规模不同种植年限的南山白毛茶园。
茶树种植密度约为6×104株/hm2,其中大行距(160±20)cm,小行距(30±10)cm,株距(25±5)cm。每年11月中旬,基肥施猪圈肥12000 kg/hm2和复合肥650 kg/hm2,沿树冠边缘垂直下方开沟,依次施入复合肥、猪圈肥,最后覆土。次年追肥3次,追肥位置与基肥相同。春茶追肥于3月中旬施尿素500 kg/hm2和复合肥1200 kg/hm2;夏茶和秋茶追肥分别于6月下旬和9月上旬施尿素300 kg/hm2和复合肥650 kg/hm2。修剪方式以轻修剪为主,每年修剪一次,在秋茶采摘后剪去树冠面上的突出枝条和树冠表层3—10 cm枝叶,然后归还土壤。在植茶过程中,没有使用除草剂,并用黄色防虫纸板代替化学农药预防害虫。
1.2 样品采集与处理
在野外调查的基础上,根据不同植茶年限茶园的地质条件和施肥情况等进行综合考虑,选择成土母质相同、坡向坡度相似且管理措施一致的植茶年限分别8年、17年、25年和43年的南山白毛茶园土壤为研究对象。首先,针对每个植茶年限布设5个典型茶园(30 m×30 m),茶园间距离约800 m;然后,在每个茶园内按“S”形设置5个采样点,具体采样位置设在树冠边缘垂直下方;最后,在每个采样点的土壤表面收集1 m2(1 m×1 m)的凋落物样品,在0—20 cm土层采集原状土样。
将每个茶园内的凋落物样品和土壤样品分别混合均匀,共得到20个混合凋落物样品和20个混合土壤样品。一方面,将每个混合凋落物样品置于80 ℃烘箱内至恒重,并称重,随后测定其相关指标(表1)。另一方面,将每个混合土壤样品沿自然结构轻轻掰开,并过5 mm筛除去动植物残体和小石块等,一部分用于测定全土性质(表1),另一部分采用改良干筛法[21]将其分为>2 mm、1—2 mm、0.25—1 mm和<0.25 mm粒径团聚体用于测定土壤团聚体性质。
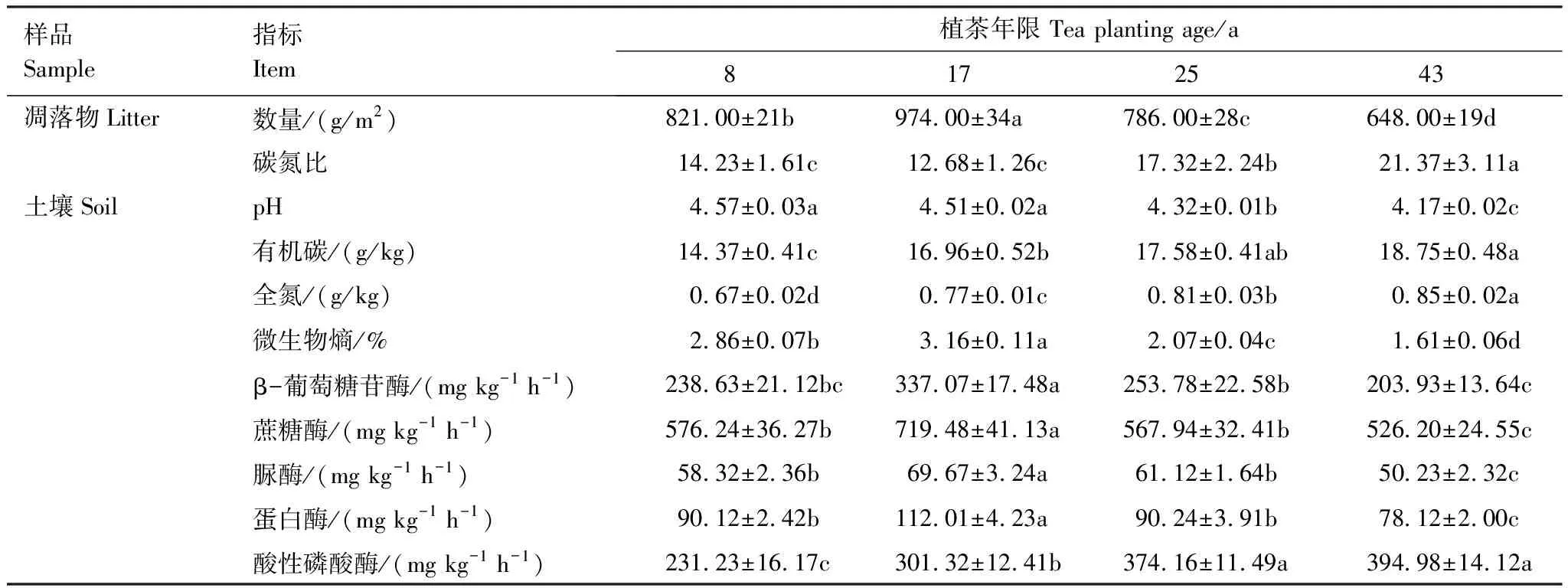
表1 不同植茶年限凋落物及土壤性质
1.3 测定项目及方法
凋落物全碳采用高温外热重铬酸钾氧化-容量法,全氮采用靛蓝比色法[22]。土壤pH值采用电位法,有机碳采用高温外热重铬酸钾氧化-容量法,全氮采用开氏法[22]。土壤β-葡萄糖苷酶采用硝基酚比色法,蔗糖酶采用3, 5-二硝基水杨酸比色法,脲酶采用苯酚-次氯酸钠比色法,蛋白酶采用茚三酮比色法,酸性磷酸酶采用磷酸苯二钠比色法[23]。
1.4 数据处理
第i粒径团聚体对全土酶活性的贡献率(CRi,%)采用邱莉萍等[24]的公式计算:
式中,EAi为第i粒径团聚体酶活性,Wi为第i粒径团聚体的质量百分含量(%),EA为全土酶活性。
全土酶活性的回收率(RR,%)采用以下公式计算:
RR=∑CRi
式中,CRi为第i粒径团聚体对全土酶活性的贡献率(%)。
统计分析在DPS 11.0软件中进行,不同处理之间多重比较及其显著性水平(P<0.05)通过最小显著性差异法进行检验,图表采用Microsoft Excel 2007软件制作。
2 结果与分析
2.1 土壤团聚体组成特征
不同植茶年限土壤团聚体组成以>2 mm粒径团聚体为主,其含量显著高于其他粒径团聚体,平均值为52.57%;其次是<0.25 mm粒径团聚体,平均值为26.08%;而1—2 mm和0.25—1 mm粒径团聚体含量较低,平均值分别为13.88%和7.47%(表2)。随着植茶年限的延长,各粒径团聚体含量的变化趋势有所不同,>2 mm粒径团聚体含量先升高后降低,在植茶17年时显著高于其他年限,<0.25 mm粒径团聚体含量则呈相反的变化趋势,而1—2 mm和0.25—1 mm粒径团聚体含量在植茶过程中变化不明显。

表2 不同植茶年限土壤团聚体组成差异
2.2 土壤团聚体酶活性
2.2.1β-葡萄糖苷酶
不同植茶年限土壤团聚体β-葡萄糖苷酶活性在>2 mm粒径中处于最高水平,其活性显著高于其他粒径团聚体,平均值为255.00 mg kg-1h-1;1—2 mm粒径团聚体次之,其β-葡萄糖苷酶活性平均值为194.78 mg kg-1h-1;而0.25—1 mm和<0.25 mm粒径团聚体β-葡萄糖苷酶活性显著低于较大粒径团聚体,平均值分别为133.36 mg kg-1h-1和128.96 mg kg-1h-1(图1)。随着植茶年限的延长,各粒径团聚体β-葡萄糖苷酶活性的变化趋势有所不同,>2 mm和1—2 mm粒径团聚体β-葡萄糖苷酶活性先升高后降低,在植茶17年时显著高于其他植茶年限,而0.25—1 mm和<0.25 mm粒径团聚体β-葡萄糖苷酶活性在植茶过程中变化不显著。

图1 不同植茶年限土壤团聚体β-葡萄糖苷酶活性差异
2.2.2蔗糖酶
土壤蔗糖酶活性在不同粒径团聚体中的变化规律与β-葡萄糖苷酶活性相似(图2)。不同植茶年限土壤>2 mm粒径团聚体蔗糖酶活性显著高于其他粒径团聚体,平均值为581.07 mg kg-1h-1;1—2 mm粒径团聚体次之,其蔗糖酶活性平均值为462.88 mg kg-1h-1;而0.25—1 mm和<0.25 mm粒径团聚体蔗糖酶活性处于较低水平,平均值分别为402.80 mg kg-1h-1和360.76 mg kg-1h-1。随着植茶年限的延长,>0.25 mm各粒径团聚体蔗糖酶活性的变化规律相似,均在植茶17年时高于其他植茶年限,而<0.25 mm粒径团聚体蔗糖酶活性在不同植茶年限间差异不显著。
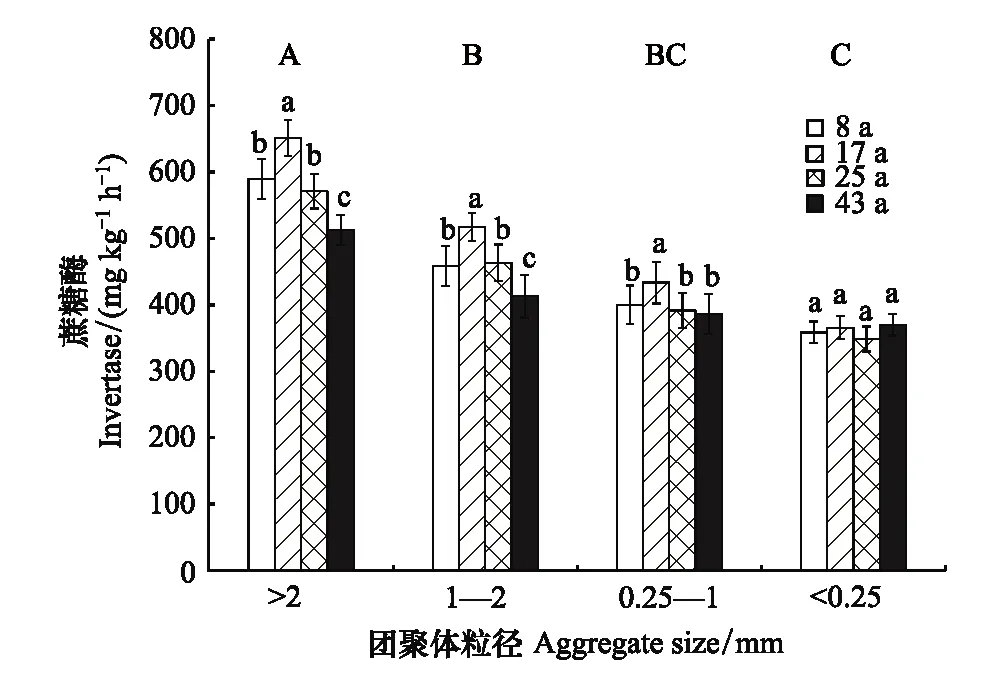
图2 不同植茶年限土壤团聚体蔗糖酶活性差异
2.2.3脲酶
不同植茶年限土壤>2 mm粒径团聚体脲酶活性最高,平均值为52.88 mg kg-1h-1,且显著高于其他粒径团聚体;1—2 mm粒径团聚体次之,其脲酶活性平均值为46.40 mg kg-1h-1;而0.25—1 mm和<0.25 mm粒径团聚体脲酶活性处于较低水平,平均值分别为39.94 mg kg-1h-1和39.02 mg kg-1h-1(图3)。随着植茶年限的延长,各粒径团聚体脲酶活性的变化趋势有所不同,>2 mm和0.25—1 mm粒径团聚体脲酶活性先升高后降低,在植茶17年时最高,其中>2 mm粒径团聚体脲酶活性在植茶17年时显著高于其他植茶年限,而1—2 mm和<0.25 mm粒径团聚体脲酶活性在植茶过程中变化不显著。

图3 不同植茶年限土壤团聚体脲酶活性差异
2.2.4蛋白酶
土壤蛋白酶活性在不同粒径团聚体中的变化规律与脲酶活性相似(图4)。不同植茶年限土壤>2 mm粒径团聚体蛋白酶活性最高,平均值为84.28 mg kg-1h-1,且显著高于其他粒径团聚体;1—2 mm粒径团聚体次之,其蛋白酶活性平均值为70.11 mg kg-1h-1;而0.25—1 mm和<0.25 mm粒径团聚体蛋白酶活性处于较低水平,平均值分别为58.14 mg kg-1h-1和53.64 mg kg-1h-1。随着植茶年限的延长,>2 mm和0.25—1 mm粒径团聚体蛋白酶活性的变化规律相似,均在植茶17年时最高,而1—2 mm和<0.25 mm粒径团聚体蛋白酶活性在不同植茶年限间差异不显著。
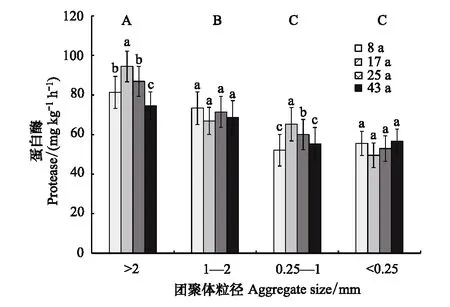
图4 不同植茶年限土壤团聚体蛋白酶活性差异
2.2.5酸性磷酸酶
各植茶年限土壤团聚体酸性磷酸酶活性在不同粒径间差异不显著,说明酸性磷酸酶活性受土壤团聚体粒径变化的影响较小(图5)。在植茶过程中,不同粒径团聚体酸性磷酸酶活性均逐年升高,其中植茶25年和43年土壤团聚体酸性磷酸酶活性显著高于植茶8年和17年。

图5 不同植茶年限土壤团聚体酸性磷酸酶活性差异
2.3 不同粒径团聚体对土壤酶活性的贡献率
不同植茶年限土壤β-葡萄糖苷酶、蔗糖酶、脲酶、蛋白酶以及酸性磷酸酶活性的平均回收率分别为80.72%、83.53%、80.13%、78.55%以及84.60%(表3)。土壤酸性磷酸酶活性的回收率在不同年限间差异不显著,而土壤碳氮循环相关酶活性(包括β-葡萄糖苷酶、蔗糖酶、脲酶以及蛋白酶)的回收率在植茶17年时显著低于其他植茶年限。另外,不同粒径团聚体对土壤酶活性的贡献率存在显著差异。不同植茶年限>2 mm粒径团聚体对土壤β-葡萄糖苷酶、蔗糖酶、脲酶、蛋白酶以及酸性磷酸酶活性的贡献率最高,为34.69%—59.07%;其次是<0.25 mm粒径团聚体,贡献率为5.44%—31.53%;而1—2 mm和0.25—1 mm粒径团聚体对土壤酶活性的贡献率较低,分别为8.07%—12.77%和2.79%—6.67%。

表3 不同植茶年限各粒径团聚体对土壤酶活性的贡献率
3 讨论
3.1 土壤团聚体粒径对酶活性的影响
土壤酶活性是评价土壤生物活性与土壤肥力的重要指标[25]。一般情况下,土壤团聚体酶被土壤胶体吸附或与之结合,从而受到物理保护,免受降解和变性失活[10-11]。由于粘结不同粒径团聚体的主要胶结物质并不相同,导致包被在团聚体内的土壤酶与胶体的结合方式及吸附能力有所差异,进而影响土壤酶活性在团聚体中的变化情况[26-27]。一些研究认为小粒径团聚体具有较高的酶活性,这主要归因于两方面,一是土壤酶易被小粒径团聚体中有机无机复合体吸附,从而受到物理保护;二是多数酶主要由细菌分泌产生,小粒径团聚体中细菌群落的丰富度和多样性较高,从而有助于土壤酶在小粒径团聚体中积累[12-13]。与之相反,另一些研究认为土壤团聚体粒径越大,较新的颗粒有机质相对较多,这为酶促反应提供了更多易被利用的基质,使得土壤酶活性在大粒径团聚体中较高[27-28]。由此可见,土壤酶活性在团聚体中的变化情况较为复杂,土地利用方式、植被覆盖类型以及生物群落组成均可能影响团聚体中酶的分布。
不同植茶年限土壤碳氮循环相关酶活性(包括β-葡萄糖苷酶、蔗糖酶、脲酶以及蛋白酶)在团聚体中的变化情况具有一致性,即>2 mm粒径团聚体酶活性最高,其次是1—2 mm粒径,而0.25—1 mm和<0.25 mm粒径团聚体酶活性处于较低水平。可见在>2 mm粒径团聚体中,较高的碳氮循环相关酶活性促进了土壤有机质的分解,进而有利于土壤养分的释放与供应[8-9]。已有研究表明,土壤碳氮循环相关酶活性在团聚体中的变化取决于土壤有机质含量在团聚体中的分布[12-13, 24]。而在本研究中,土壤团聚体碳氮循环相关酶活性与土壤有机质含量的分布并非具有一致性。不同植茶年限土壤有机碳和全氮含量在>2 mm和1—2 mm粒径团聚体中均较高(表4)。不同的是,土壤碳氮循环相关酶活性虽然在>2 mm粒径团聚体中较高,但在1—2 mm粒径团聚体中却处于较低水平。导致这一现象的原因可能与团聚体中有机质有效性的分布差异有关。
作为土壤肥力水平和质量状况的有效指标,土壤微生物熵(微生物量碳/有机碳)能够准确反映土壤有机质质量变化,其值越大,有机质有效性越高[29]。不同植茶年限土壤微生物熵在>2 mm粒径团聚体中显著高于1—2 mm粒径(表4)。说明土壤微生物利用底物的有效性在1—2 mm粒径团聚体中相对较低,从而导致该粒径团聚体中酶活性较低。相反,>2 mm粒径团聚体中土壤有机质有效性处于较高水平,能够为微生物的生长提供更多的能量与养分,从而有助于微生物的生长和酶活性的提高。这也进一步表明,在本研究中土壤团聚体有机质质量对碳氮循环相关酶活性分布的影响较有机质数量更大。不同植茶年限土壤酸性磷酸酶活性在不同粒径团聚体间差异不显著,说明该酶活性受土壤团聚体粒径变化的影响较小。

表4 不同植茶年限土壤团聚体有机碳、全氮和微生物熵的差异
3.2 土壤团聚体酶活性对植茶年限的响应
土壤碳氮循环相关酶活性(包括β-葡萄糖苷酶、蔗糖酶、脲酶以及蛋白酶)是参与土壤有机质转化与循环的重要酶类,可作为评价土壤有机质固持与分解潜力的敏感指标[8-9]。在本研究中,长期植茶有助于土壤有机质积累(表1)。但不同的是,随着植茶年限的延长,土壤碳氮循环相关酶活性则先升高后降低,在植茶17年时处于较高水平,究其原因有以下三点。其一,茶树凋落物是茶园生态系统的重要组成部分,不同植茶阶段进入土壤的凋落物数量与质量不尽相同,从而影响土壤碳氮循环相关酶活性[30-31]。在植茶前期(8年—17年),茶树凋落物的C/N处于较低水平(表1),说明在此期间凋落物中易分解组分相对较多,且归还土壤的数量逐渐升高(表1),这为酶促反应提供了更多易被利用的基质,使得土壤碳氮循环相关酶活性有所升高;而在植茶后期(17年—43年),由于茶树逐渐衰老,凋落物的数量和质量均显著降低(表1),从而导致土壤碳氮循环相关酶活性也随之降低。其二,在植茶过程中土壤多酚类化合物逐年积累[32],而酚类物质能够抑制部分水解酶活性,如β-葡萄糖苷酶和蔗糖酶[33]。其三,植茶17年以后土壤pH值显著降低(表1),土壤酸化通过抑制微生物分泌酶和基质有效性来降低土壤碳氮循环相关酶活性[32-33]。由此可见,在植茶前期土壤积累的有机质有效性相对较高,且土壤碳氮循环相关酶活性逐年升高,说明在该阶段土壤积累的有机质更易被矿化分解,从而释放更多养分供茶树生长;而在植茶后期土壤积累的有机质有效性相对较低,同时伴随着土壤碳氮循环相关酶活性逐年降低,表明在该阶段土壤积累的有机质分解速率较低,从而有助于土壤有机碳及养分的贮存。值得注意的是,不同粒径团聚体碳氮循环相关酶活性对植茶年限的响应有所不同。随着植茶年限的延长,>2 mm粒径团聚体碳氮循环相关酶活性在植茶17年时最高;而在<0.25 mm粒径团聚体中,土壤碳氮循环相关酶活性在各植茶年限间差异不显著。说明大粒径团聚体碳氮循环相关酶活性对植茶年限的响应较小粒径团聚体更为敏感。在酸性土壤环境中,酸性磷酸酶参与着土壤有机磷的活化,其活性高低直接影响着土壤有机磷的分解转化及其生物有效性[16]。在本研究中,土壤酸性磷酸酶活性在植茶过程中有所升高,这可能与土壤有机质逐年积累有关[34-35]。
3.3 不同粒径团聚体对土壤酶活性的贡献率
不同土壤酶活性在团聚体分级过程中均有不同程度损失,尤其是蛋白酶,其活性的平均回收率已低于80%。这是因为在团聚体分级过程中团聚体遭到破坏,导致其中土壤酶因缺少物理保护而失活,从而造成土壤酶活性的损失[36-37]。土壤碳氮循环相关酶活性(包括β-葡萄糖苷酶、蔗糖酶、脲酶以及蛋白酶)的回收率在植茶17年时显著低于其他植茶年限,而酸性磷酸酶活性的回收率在不同植茶年限间差异不显著。在本研究中,植茶17年土壤含有更多>2 mm粒径团聚体(表2)。该粒径团聚体中碳氮循环相关酶活性虽然较高,但这些酶也会因较大粒径团聚体稳定性较低而更易遭到破坏,从而导致植茶17年土壤碳氮循环相关酶活性在团聚体分级过程中损失更为严重。
将不同粒径团聚体的组成比例与其中酶活性进行综合考虑,可以全面反映各粒径团聚体对土壤酶活性的贡献率[12]。在本研究中,不同粒径团聚体对土壤酶活性的贡献率由高到低表现为:>2 mm团聚体>、<0.25 mm团聚体>、1—2 mm团聚体、>0.25—1 mm团聚体。可见,>2 mm粒径团聚体是茶园土壤酶的主要载体。值得注意的是,虽然土壤碳氮循环相关酶活性在<0.25 mm粒径团聚体中处于较低水平,但由于土壤中该粒径团聚体的组成比例较高(表2),所以该粒径团聚体对土壤碳氮循环相关酶活性的贡献率才显示出较高值。这也说明,不同粒径团聚体对土壤酶活性的贡献率与团聚体的组成比例高度相关。
4 结论
土壤碳氮循环相关酶活性在植茶过程中先升高后降低,在植茶17年时处于较高水平,而土壤酸性磷酸酶活性则逐年升高。土壤碳氮循环相关酶活性在>2 mm粒径团聚体中处于较高水平,而酸性磷酸酶活性受团聚体粒径变化的影响较小。作为土壤酶的主要载体,>2 mm粒径团聚体的形成与稳定对于促进土壤有机碳及养分循环具有重要意义。在茶园管理中,植茶17年以后应注意土壤>2 mm粒径团聚体的崩解与破坏问题,以保障土壤质量及维持土壤有机碳及养分利用效率。
猜你喜欢
中草药(2022年20期)2022-11-15
腐植酸(2020年1期)2020-11-29
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
中成药(2018年5期)2018-06-06
中成药(2017年8期)2017-11-22
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
天然产物研究与开发(2016年6期)2016-06-05
医学研究杂志(2015年8期)2015-06-22
天然产物研究与开发(2014年3期)2014-04-27
中国医学科学院学报(2013年6期)2013-03-11
