
雌激素受体基因对泥鳅雌雄和倍性生长差异的表达调控作用
2020-12-02 05:27李丹阳罗丽飞曹文怡周小云高泽霞
华中农业大学学报(自然科学版) 2020年6期
李丹阳,罗丽飞,曹文怡,周小云,高泽霞,2
1.华中农业大学水产学院/农业农村部淡水生物繁育重点实验室/长江经济带大宗水生生物产业绿色发展教育部工程研究中心/农业动物遗传育种与繁殖教育部重点实验室, 武汉 430070;2.湖北省名优鱼育种与健康养殖工程技术研究中心, 武汉 430070
在哺乳动物中,17β-雌二醇(17β-Estradiol,17β-E2)可通过雌激素受体在个体发育中发挥调控摄食行为、调节能量平衡的重要作用。有研究表明,将E2注射到小鼠不同的大脑区域会改变动物的摄食行为和体质量[1]。有研究报道卵巢切除的小鼠出现体质量增加、脂肪含量升高的表型,并且这种肥胖表型可以通过体外补充E2来减弱[2-3]。雌激素受体(estrogen receptor,ER)是雌激素发挥生理作用的关键所在[4]。在哺乳动物中发现雌激素受体只有ERα和ERβ2种亚型[5-6],而在硬骨鱼类中普遍存在ERα、ERβ1和ERβ2三种亚型,例如尼罗罗非鱼(Oreochromisniloticus)[7]、斑马鱼(Daniorerio)[8]、大鳞副泥鳅(Paramisgurnusdabryanus)[9]。ER除可以调节性腺生长和分化外,在机体能量代谢方面也发挥重要作用。在哺乳动物中,ERα基因在调节机体能量代谢过程中比ERβ基因发挥着更为重要的生理功能,例如一些学者研究发现ERα基因突变体小鼠的体质量明显比正常小鼠大,具有典型的肥胖特征,而ERβ特异缺失的小鼠的体质量和正常小鼠没有显著差异[10-11]。性别异形现象在鱼类中普遍存在,部分鱼类雌雄生长速度差异显著,且表现出物种特异性。此外,鱼类多倍体通常表现为比二倍体更快的生长速度[12]。目前,关于雌激素在鱼类中的研究还停留在初步的探索阶段,还没有雌激素受体相关基因在调控鱼类个体生长速度差异方面的研究报道。
泥鳅(Misgurnusanguillicaudatus)是我国重要的小型淡水经济鱼类,生长速度快,繁殖能力强,在自然界中普遍存在多倍体现象[12-13]。泥鳅雌性生长速度明显快于雄性[14],且四倍体泥鳅的生长速度明显快于二倍体[15]。本研究克隆泥鳅雌激素受体3个基因ERα、ERβ1、ERβ2的cDNA核心序列,采用qRT-PCR对二倍体泥鳅生长发育的5个关键时期4种组织以及泥鳅二倍体和四倍体成鱼不同组织中ER基因的表达量进行分析,旨在为研究ER调控鱼类雌雄和倍性生长差异的分子机制提供基础。
1 材料与方法
1.1 试验材料
试验所用泥鳅均为笔者所在实验室的人工繁殖群体。采集二倍体泥鳅性成熟前后生长发育阶段5个时期(3、6、9、12、18月龄)和18月龄四倍体泥鳅,随机选取体型正常泥鳅雌雌各6尾,其中二倍体泥鳅各月龄阶段雌雄生长差异见图1;采集的18月龄四倍体泥鳅雌性、雄性平均体质量分别为(11.57±5.31) g和(7.35±2.08) g,均显著高于18月龄二倍体泥鳅雌雄体质量(P<0.05)。个体经MS-222(100 mg/L)麻醉后,迅速采集脑、肌肉、肝脏和性腺组织,所有样品液氮速冻后转-80 ℃冰箱保存,用于后续RNA提取。

*表明雌雄间存在显著差异(P<0.05);**表明雌雄间存在极显著差异(P<0.01)。Note: The super “*” means significant difference between females and males (P<0.05); “**” means extremely significant difference between females and males (P<0.01).
1.2 总RNA提取、cDNA合成及检测
采集的各样品总RNA利用Trizol(TaKaRa,大连)法按照说明书步骤提取。后续RNA的纯度、完整性检测以及高质量RNA反转录成cDNA的方法均参考陈宇龙等[16]方法。
1.3 ER基因的克隆及序列分析
将NCBI上下载的斑马鱼ERα、ERβ1和ERβ2基因的cDNA 序列与实验室前期已获得的泥鳅转录组数据进行blast,获得泥鳅3个目的基因的cDNA片段。 用Primer Premier 5.0软件针对泥鳅3个基因设计简并引物,引物由武汉擎科有限公司合成(表1)。以反转录得到的cDNA为模板,将3个基因合成引物分别进行PCR扩增。用1%琼脂糖凝胶电泳检测PCR产物,将条带单一的产物送武汉擎科有限公司测序。测序结果经blast(https://blast.ncbi.nlm.nih.gov/Blast.cgi)后,将正确的目的基因片段利用Mega 7.0软件进行拼接。最后将拼接序列与斑马鱼基因序列进行比对以确定编码区是否完整。

表1 基因克隆和qRT-PCR分析引物信息 Table 1 Primer sequences for genes clone and qRT-PCR analysis
1.4 泥鳅ER基因理化性质分析及进化树的构建
在NCBI的ORF finder 里分析ERα基因开放阅读框的序列信息(https://www.ncbi.nlm.nih.gov/orffinder/);使用ProtParam(http://web.expasy.org/protparam/)对ERα基因的分子质量及等电点等理化性质进行分析;使用DNAman软件对来自不同物种、不同进化地位的脊椎动物的ERα基因进行氨基酸序列比对,并使用Mega 7.0软件构建ER基因3个亚型的系统进化树,进行雌激素受体基因3个亚型的聚类分析。
1.5 ER基因表达分析
基于所得到的泥鳅ERα、ERβ1和ERβ2的序列信息,利用Primer Premier 5.0设计其定量引物,由武汉擎科有限公司合成(表1),以泥鳅β-actin为内参基因。使用QuantStudio 6 Flex实时荧光定量PCR仪以及SYBR©Premix Ex TaqTM II (TliRNaseH Plus)试剂盒(TAKARA,大连),分别以二倍体泥鳅不同发育时期不同组织样品以及二倍体和四倍体成鱼的脑、肌肉、肝脏和性腺组织的cDNA为模板,进行qRT-PCR反应。用2-ΔΔCt计算出ER基因的3个亚型各自在泥鳅各组织样品中的相对表达量。通过SPSS软件中的Duncan’s Multiple Range Test来分析雌激素受体3个基因亚型在二倍体泥鳅性成熟前后5个关键时期的表达情况及二倍体、四倍体相关组织间的差异表达。
2 结果与分析
2.1 泥鳅ER理化性质分析
利用同源克隆拼接核心序列,获得了泥鳅ERα、ERβ1和ERβ2基因的cDNA核心序列,分别为1 821、1 008、1 197 bp。其中ERα基因获得了完整的CDS序列,ProtParam软件预测结果显示,泥鳅ERα基因由607个氨基酸残基构成,分子式为C2903H4606N840O898S43,分子质量为67.02 ku,理论等电点(PI)为8.63,为碱性蛋白。氨基酸残基中Arg(6.6%)、Gly(7.2%)、Ser(12.4%)、Leu(9.1%)的频率较高,极性氨基酸占62.4%,非极性氨基酸占37.6%(图2)。消光系数(M-1cm-1γ=280 nm)为61 530,不稳定系数为66.06,属不稳定类蛋白;疏水指数为69.41,平均亲水性为-0.45,属于可溶性蛋白。

图2 泥鰍ERα基因氨基酸频率分布图
2.2 泥鳅ER氨基酸序列比对分析
使用DNAman软件对人(Homosapiens)、鼠(Musmusculus)、鸡(Gallusgallus)、泥鳅(Misgurnusanguillicaudatus)等几个物种3个雌激素受体基因的氨基酸进行同源性分析,并使用 Mega软件构建进化树。在进化树中,ERα和ERβ亚型各聚为一类,形成2个大分支(图3);在ERβ亚型中,鱼类ERβ1和ERβ2各单独聚为一支,再与其他脊椎动物的ERβ聚在一起;泥鳅ERβ1和ERβ2均与鲤和斑马鱼在进化树中距离最近。泥鳅ERα蛋白的氨基酸序列与鲤(83.44%)和斑马鱼(85.02%)具有较高的同源性;与其他鱼类具有中度同源性(55%~60%),而与人(46.39%)、鼠(46.83%)、家鸡(46.8%)具有低度同源性(图4)。
2.3 泥鳅ER基因在雌雄生长发育过程中的表达比较
采用基因荧光定量表达分析方法比较了ERα、ERβ1和ERβ2基因在泥鳅3、6、9、12、18月龄的雌雄个体中的差异表达。结果显示,3个基因在泥鳅生长发育过程中的表达存在一定差异(图5)。其中在雌性的各个组织中,ERα在脑、肌肉、肝脏和性腺组织中的表达量整体呈现出先上升后下降的趋势(图5A、B、C、D)。ERβ1在脑、肌肉和性腺中的表达量整体呈现出先下降后上升的趋势(图5E、F、H),而在肝脏组织中的表达量整体呈现出先上升后下降的趋势。ERβ2在脑和肝脏组织中的表达量整体呈现出先上升后下降的趋势(图5I、K),而在肌肉和性腺组织中整体呈现先下降后上升的趋势(图5J、L)。在雄性的各个组织中,ERα在肝脏和精巢组织中的表达量随着个体的生长整体呈现出先上升后下降的趋势(图5C、D);在脑组织中表达量逐渐下降(图5A),而在肌肉组织逐渐上升(图5B)。ERβ1在各个组织中的表达量整体均呈现出先下降再上升的趋势(图5E、F、G、H)。ERβ2在脑和肌肉组织中的表达量整体均呈现出逐渐下降的趋势(图5I、J),而在肝脏和精巢组织中的表达量整体呈现出逐渐上升的趋势(图5K、L)。在泥鳅雌雄生长发育过程中,ERβ1和ERβ2在脑组织中的表达模式表现相反,ERβ1整体呈现出先下降再上升的趋势,而ERβ2整体呈现出先上升再下降的趋势。此外,ERα和ERβ2在肌肉中的表达模式相反;ERα、ERβ1和ERβ2在雌性肝脏组织中有相同的表达模式,均呈现出先上升后下降的表达趋势;ERβ1和ERβ2在卵巢中也有相同的表达模式,均呈现出先下降后上升的表达趋势;ERα基因在雌雄相同组织中的表达比较结果显示,ERα在雄性各生长时期的脑组织、肌肉组织和性腺组织中的表达量均显著高于雌性(P<0.05)(图5A、B、D),而在雌性肝脏组织中的表达量显著高于雄性(图5C)。ERβ1在各个生长时期精巢组织中的表达量均显著高于卵巢(图5H),而在雌性各个时期肝脏组织中的表达量显著高于雄性(图5G)。ERβ2在雄性各个生长时期肌肉和精巢组织的表达量均显著高于雌性(图5J、L),3、6、18月龄雄性脑组织的表达量显著高于雌性,而在9、12月龄雌性脑组织的表达量显著高于雄性(图5I)。
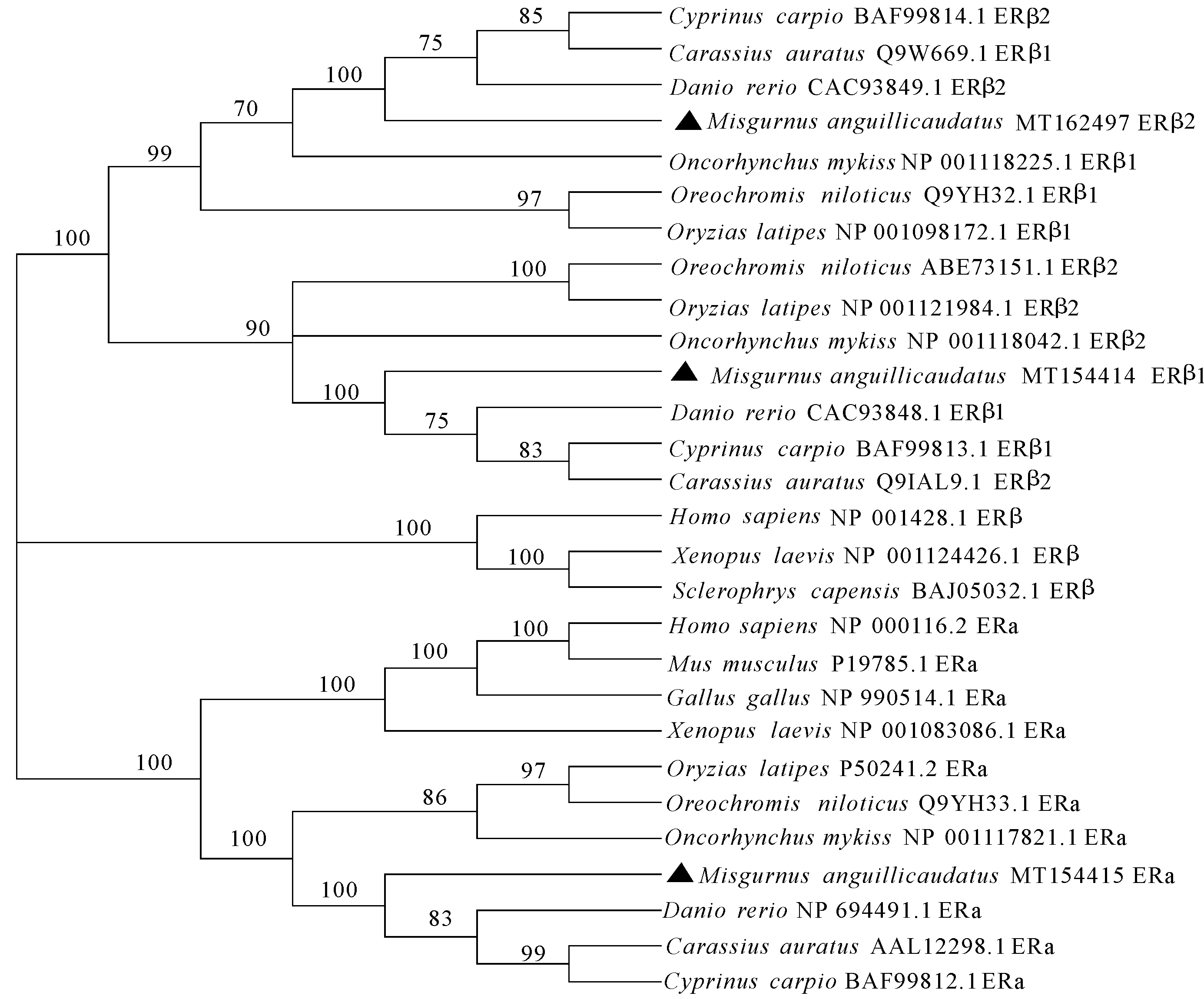
图3 ERs的系统进化树分析

a:鲤 Cyprinus carpio; b:斑马鱼 Danio rerio; c:虹鳟 Oncorhynchus mykiss; d:青鳉 Oryzias latipes; e:罗非鱼 Oreochromis niloticus; f:人 Homo sapiens; g:鼠 Mus musculus; h:鸡 Gallus gallus.

组间标注不同小写字母表示这些组中雌激素受体基因的表达具有显著性差异(P<0.05)。Different lowercase letters between groups indicate that the expression of estrogen receptor genes in the groups is significantly different (P<0.05).
2.4 二倍体和四倍体泥鳅ER基因的表达差异分析
泥鳅雌激素受体基因ERα、ERβ1和ERβ2在泥鳅不同倍性以及不同性别间表达均有差异,如图6所示:在脑组织中,同一性别的二倍体泥鳅ERα和ERβ2 表达量均显著高于四倍体(P<0.05)(图6A和6C);在同一倍性中,雄性ERα和ERβ2的表达量显著高于雌性(P<0.05)(图6A和6C)。雄性二倍体ERβ1的表达量显著高于四倍体(P<0.05),二倍体、四倍体雌性间无显著差异(图6B)。在肌肉组织中的表达结果显示,雄性二倍体ERα的表达量显著高于四倍体(P<0.05)(图6D);雌雄ERβ1和ERβ2在四倍体的表达量均高于二倍体(P<0.05)(图6E和6F)。在肝脏组织中,雌雄ERα在四倍体的表达量显著高于二倍体(P<0.05)(图6G);在雌性个体中,四倍体ERβ1和ERβ2的表达量均高于二倍体(P<0.05);在雄性个体中,二倍体ERβ1和ERβ2的表达量均高于四倍体(P<0.05)(图6H和6I)。在性腺组织中,无论倍性,ERα均在雄性中的表达量显著高于雌性(P<0.05)(图6J);ERβ1在雄性四倍体的表达量显著高于二倍体的(P<0.05)(图6K),雌性倍性间无显著差别;无论倍性,ERβ2在雄性中的表达量均显著高于雌性(P<0.05)(图6L)。
3 讨 论
本研究获得了泥鳅雌激素受体基因ERα、ERβ1和ERβ2的CDs区核酸的序列信息,分别采用PCR、基因克隆和测序等技术,并通过ProtParam进一步分析ERα基因的理化性质,可为研究ER基因对泥鳅雌雄和倍性生长差异的表达调控作用提供理论依据。对泥鳅ERα基因序列分析发现,该基因的长度和其所编码的氨基酸数量与斑马鱼和鲤最相似,相似度在80%~90%。在进化树中也表现出相似的结果,ERα基因与斑马鱼和鲤距离最近,再次验证了ERα基因与各个物种的亲缘关系。物种亲缘关系的远近可以由基因的同源性来间接表明。本研究克隆得到的泥鳅ERα、ERβ1和ERβ2 氨基酸序列比对结果表明泥鳅的3种ERs与鲤和斑马鱼氨基酸序列的一致性高达80%以上,表明ERs在鲤科鱼类中高度保守。
本研究中ERα、ERβ1和ERβ2三个基因表达结果显示,在二倍体泥鳅不同生长发育阶段,ERα都表现为在雄性的脑、肌肉和性腺组织的表达量显著高于雌性;倍性间比较结果显示,ERα、ERβ1和ERβ2基因在二倍体泥鳅脑组织的表达量均显著高于四倍体。结合泥鳅雌雄之间以及二倍体和四倍体的生长特性,结果表明雌激素受体ERα普遍在生长速度较慢个体的脑组织中高表达。已有报道表明无论是在人类还是其他哺乳动物中,ERα基因在调节体质量平衡上扮演着重要的角色。例如,Xu等[11]和 Heine等[17]通过基因敲除技术获得只在脑组织中特异缺失ERα基因的小鼠,实验小鼠表现为摄食增多、活动减少,从而导致肥胖。而抑制ERβ则不会出现肥胖表型,此外ERα选择性激动剂可以有效地抑制摄食,而ERβ选择性激动剂则不会改变摄食行为[10-11]。由此我们推测脑组织中ERα的高表达可能在一定程度上起到了抑制雄性泥鳅生长的作用。ERβ1和ERβ2在脑组织中的表达模式刚好相反,ERβ1整体呈现出先下降再上升的趋势,而ERβ2整体呈现出先上升再下降的趋势,说明这两个基因可能在脑组织中相互协调发挥一定的生理作用。有研究证据表明,ERβ在预防饮食引起的肥胖方面起着重要的作用,但ERβ具体的调节机制有待进一步研究[18]。ERα在各月龄雄性的肌肉组织中整体呈现出一个逐渐上升的趋势,而ERβ2整体呈现出一个逐渐下降的趋势,表明ERα和ERβ2在泥鳅性成熟前后的肌肉组织中相互协调发挥一定的生理作用。ERα和ERβ2在雄性肌肉组织的表达量均显著高于雌性,这在一定程度上证实了雌激素可以直接抑制肌肉中GHR编码mRNA的水平的结论[19],同时,这也可能是泥鳅雄性的生长速率比雌性低的原因之一。本研究中,ERα基因在雌雄肝脏组织中的表达量均呈现出先上升再逐渐下降的趋势,且各个月龄雌性的表达量显著高于雄性。这与海水青鳉[20]和斑马鱼的研究结果一致[21]。众所周知,雌二醇通过肝脏合成的卵黄蛋白原释放到血液再转运到卵母细胞。ERα、ERβ1和ERβ2 基因在精巢的表达量均高于卵巢,这与在斑马鱼[22]和虹鳟[23]中的研究结果一致,提示ER基因在介导雌激素调控早期精子的发生、成熟和排出中发挥重要的作用。并且ER基因在性腺组织的这种表达规律在脊椎动物中的保守性较高[24]。
泥鳅雌激素受体基因3个亚型(ERα、ERβ1和ERβ2)在不同组织间表达丰度不同,表明其在不同组织可能参与不同的生理功能。在泥鳅生长发育过程中无论雌雄,ERβ1和ERβ2在脑组织中的表达模式刚好相反,ERβ1整体呈现出先下降再上升的趋势,而ERβ2整体呈现出先上升再下降的趋势;除此之外,ERα和ERβ2在肌肉中的表达模式刚好相反;ERα、ERβ1和ERβ2在雌性肝脏组织中有相同的表达模式,均呈现出先上升后下降的表达趋势;ERβ1和ERβ2在卵巢中也有相同的表达模式,均呈现出先下降后上升的表达趋势。这些研究结果表明:雌激素受体的各亚型组成一个调控体系,在鱼类生长发育不同阶段各组织器官的生长和新陈代谢过程中发挥着重要的作用。
本研究通过对泥鳅雌激素受体基因ERs的克隆及同源性分析,发现ERs在鲤科鱼类中具有较高的保守性。泥鳅生长发育5个关键时期以及在二倍体、四倍体间不同组织的定量表达分析结果表明脑组织中ERα基因对鱼类个体生长速度可能具有一定的抑制作用。ERα、ERβ1和ERβ2三个基因组成一个复杂的调控体系,彼此之间相互调节,在机体生长发育中共同发挥作用。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
今日畜牧兽医(2022年10期)2022-12-23
现代园艺(2017年13期)2018-01-19
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
中成药(2016年4期)2016-05-17
Coco薇(2015年12期)2015-12-10
中国果业信息(2015年6期)2015-01-23
食品科学(2013年17期)2013-03-11
大学(2008年4期)2008-07-10
