
柚皮苷对破骨细胞分化和极化影响的初步探究
2020-12-18 05:54刘朝晖马剑雄田爱现孙晓雷张杨郭悦马信龙
天津医药 2020年12期
刘朝晖,马剑雄,田爱现,孙晓雷,张杨,郭悦,马信龙△
骨质疏松症(osteoporosis,OP)是一种以骨量降低与骨微结构退化为特征的运动系统疾病,骨折是其最常见的并发症[1]。预计到2050年,我国骨质疏松症患者将增至599万例,相关治疗费用将高达254.3亿美元[2]。因此,OP的防治具有重要的社会意义。
破骨细胞起源于造血干细胞,由单核细胞融合分化而成,是唯一具有骨吸收功能的细胞,其分泌的组织蛋白酶K(CK)和基质金属蛋白酶-9(MMP-9)是破骨细胞降解骨基质的重要蛋白水解酶[3]。破骨细胞具有独特的细胞器分布和质膜结构域,与骨组织接触的细胞膜区域通过特殊的黏附结构形成封闭区,其间的细胞膜具有微绒毛结构样褶皱缘,两者形成相对独立的骨吸收间隙,完成细胞极化[4]。整合素(integrin)是细胞表面受体超家族蛋白,调节黏附、迁移及封闭区的形成,对破骨细胞分化和激活具有重要意义[5-6]。非受体酪氨酸激酶(c-src)是一种原癌基因,c-src及其磷酸化产物p-src调节细胞极化、迁移及褶皱缘的形成,敲除c-src的小鼠体内破骨细胞无法形成正常伪足,导致小鼠患骨硬化症[7-8]。
骨碎补是治疗OP的主要中药之一,具有不良反应小,疗效明显等优点,其主要有效成分是双氢黄酮类单体柚皮苷。有关柚皮苷抗OP 的分子机制的研究多集中于相关信号通路方面[9-11],然而关于柚皮苷和破骨细胞分化极化关系的研究并未见报道。本文旨在探究柚皮苷能否通过影响破骨细胞分化与极化进而影响破骨细胞的生物学功能。
1 材料与方法
1.1 材料 柚皮苷购自美国Sigma 公司,小鼠单核细胞RAW264.7细胞株购自武汉普诺赛公司,高糖DMEM培养液、胎牛血清(FBS)均购自美国Gibco公司,重组小鼠可溶性核因子κB 受体活因子配体(sRANKL)、兔integrin β3 一抗购自美国Peprotech 公司,抗酒石酸酸性磷酸酶(TRAP)染色试剂盒购自北京索莱宝公司,小鼠MMP-9 一抗购自美国R&D Systems公司,兔CK一抗、羊抗鼠二抗、羊抗兔二抗购自英国Abcam 公司,兔p-src 一抗购自美国Cell Signaling Technology公司,实时荧光定量逆转录聚合酶链反应(qPCR)引物购自武汉塞维尔公司,qPCR试剂盒购自北京Takara公司。
1.2 细胞培养 小鼠RAW264.7 细胞系培养于含10% FBS的DMEM 培养基中,于37 ℃、5%CO2条件下培养,隔1 d换液1次,细胞密度约为80%时传代至6孔板。配置含10%FBS,100 μg/L sRANKL。对照组加入0 μg/L 柚皮苷培养液,实验组分别加入2、20 和200 μg/L 柚皮苷培养液,每组3 个复孔,于37 ℃、5%CO2条件下培养,隔日换液,连续培养7 d。
1.3 TRAP 染色 细胞培养7 d 后,弃掉培养液,PBS 冲洗3次,4%多聚甲醛固定60 s,PBS冲洗,使用TRAP染液于37 ℃下避光孵育1 h,PBS 冲洗后使用苏木素复染30 s。4 组细胞均在100 倍视野下,随机读取10 个视野进行阳性细胞计数(阳性标准:细胞核≥3个且胞质呈酒红色)。
1.4 qPCR 检测CK、MMP-9、integrin β3及c-srcmRNA 相对表达量 不同剂量柚皮苷干预细胞7 d,待细胞融合至90%时,依次使用Trizol、氯仿、乙醇及DEPC 水等抽提总RNA,操作均在冰上进行,根据试剂说明书进行RNA 的逆转录及cDNA 的扩增。逆转录条件:42 ℃15 min,85 ℃5 s。扩增条件:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火30 s,72 ℃延伸15 s,共40个循环。以GAPDH为内参,目的基因相对表达量采用2-ΔΔCt法。引物序列见表1。

Tab.1 Amplification of gene primers by fluorescence quantitative PCR表1 qPCR扩增基因引物序列
1.5 蛋 白 免 疫 印迹(Western blot)法检测CK、MMP-9、integrin β3 及p-src 蛋白相对表达量 不同剂量柚皮苷干预细胞7 d,待细胞融合至90%时,在孔板中加入含PMSF 的高效组织裂解液RIPA,冰上反复吹打,12 000×g离心10 min 后取上清。加入上样缓冲液,95 ℃加热10 min 使蛋白变性,10%SDS-PAGE 分离蛋白后,转至PVDF 膜上,5%脱脂奶粉封闭抗原1 h 后,稀释一抗CK(1∶1 000)、MMP-9(1∶800)、integrin β3(1∶1 000)及p-src(1∶1 000),4 ℃孵育过夜,隔天TBST洗膜3次,稀释二抗,羊抗鼠二抗(1∶20 000)、羊抗兔二抗(1∶20 000),室温孵育1 h,TBST洗膜3次,加入ECL化学发光液显影曝光。分析各组CK、MMP-9、integrin β3及p-src蛋白相对表达量。
1.6 统计学方法 采用SPSS 20.0 进行统计学分析,计量资料采用均数±标准差(±s)表示,多组间比较采用单因素方差分析,组间多重比较采用Dunnet-t检验,P<0.05为差异有统计学意义。
2 结果
2.1 柚皮苷对破骨细胞计数结果的影响 对照组、2 μg/L 组、20 μg/L 组和200 μg/L 组阳性破骨细胞计数(个/100 倍视野)分别为29.50±2.22、23.20±1.81、20.10±0.88和16.20±1.03。与对照组相比,各实验组TRAP 染色阳性细胞数均显著降低(n=10,F=104.612,P<0.05),见图1。
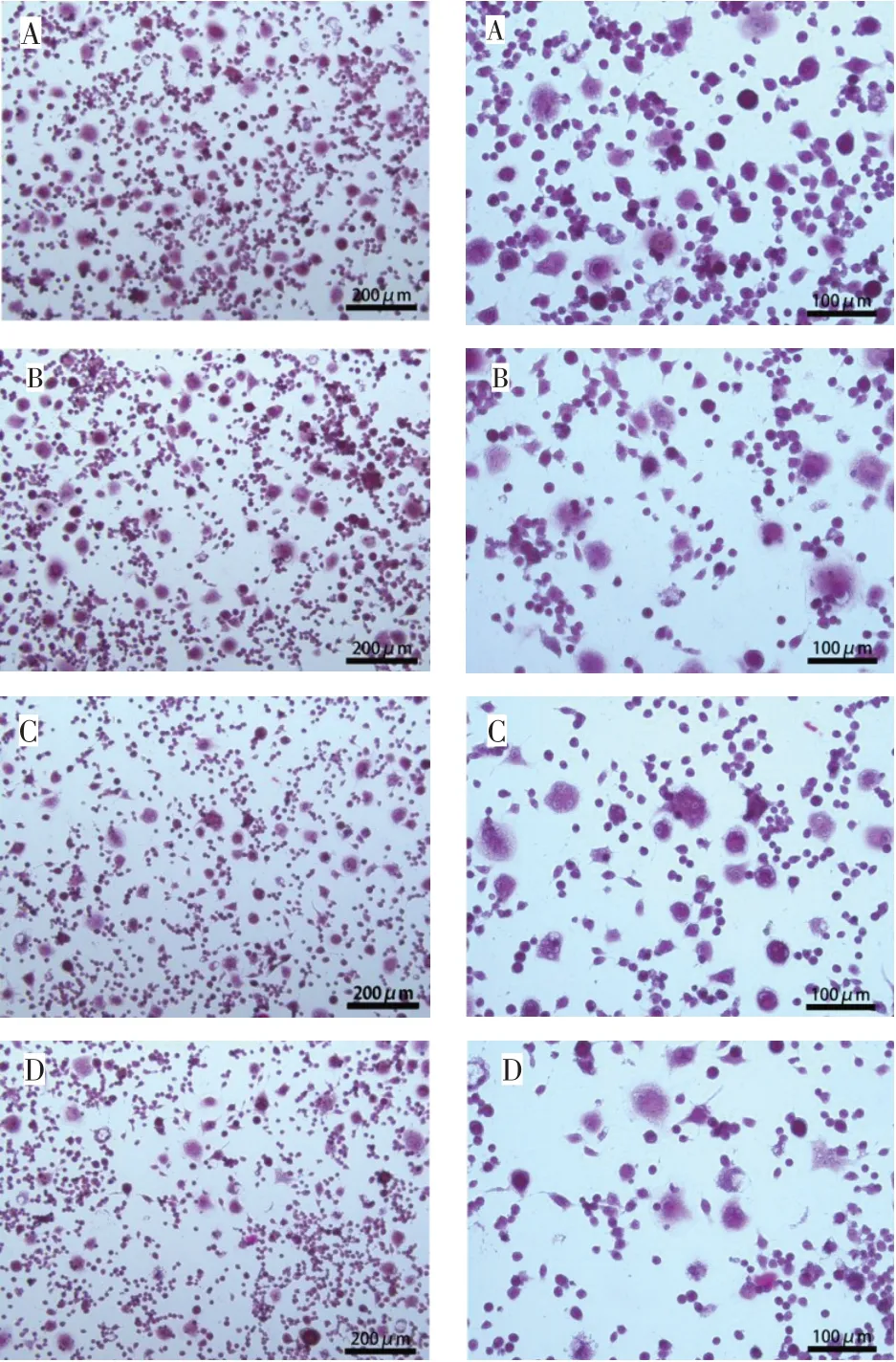
Fig.1 Effects of naringin on TRAP staining(left×100,right×200)图1 柚皮苷对破骨细胞计数结果的影响(左×100,右×200)
2.2 柚皮苷对CK、MMP-9、integrin β3、c-srcmRNA表达水平的影响 与对照组比较,20 μg/L和200 μg/L 组CK、MMP-9、integrin β3mRNA 表达量显著降低(P<0.05),各实验组c-srcmRNA 表达量显著降低(P<0.05),见表2。
Tab.2 Effects of naringin on mRNA expressions of CK,MMP-9,integrin β3 and c-src表2 各组CK、MMP-9、integrin β3及c-src mRNA表达水平的比较(n=3,±s)

Tab.2 Effects of naringin on mRNA expressions of CK,MMP-9,integrin β3 and c-src表2 各组CK、MMP-9、integrin β3及c-src mRNA表达水平的比较(n=3,±s)
*P<0.05,a与对照组比较,b与2 μg/L组比较,c与20 μg/L组比较,P<0.05;表3同
组别对照组2 μg/L组20 μg/L组200 μg/L组F CK 1.00±0.00 0.77±0.21 0.67±0.01a 0.44±0.10ab 11.607*MMP-9 1.00±0.00 0.75±0.16 0.67±0.19a 0.60±0.18a 8.725*intergrin β3 1.00±0.00 0.73±0.15 0.68±0.16a 0.49±0.08a 9.857*c-src 1.00±0.00 0.75±0.06a 0.68±0.09a 0.39±0.09abc 37.453*
2.3 柚皮苷对CK、MMP-9、integrin β3、p-src蛋白表达水平的影响 与对照组比较,20 μg/L组和200 μg/L 组CK 和p-src 蛋白表达水平显著降低(P<0.05),各实验组MMP-9和integrin β3蛋白表达水平均显著降低(P<0.05)。见图2、表3。
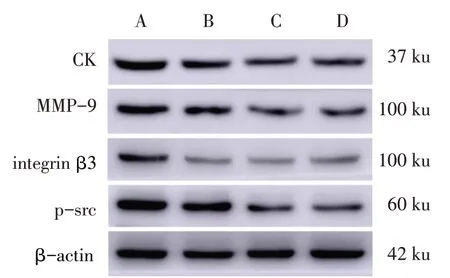
Fig.2 Effects of naringin on protein expressions of CK,MMP-9,integrin β3 and p-src图2 各组CK、MMP-9、integrin β3及p-src蛋白相对表达量的比较
Tab.3 Effects of naringin on protein expressions of CK,MMP-9,integrin β3 and p-src表3 各组CK、MMP-9、integrin β3及p-src蛋白相对表达水平的比较(n=3,±s)

Tab.3 Effects of naringin on protein expressions of CK,MMP-9,integrin β3 and p-src表3 各组CK、MMP-9、integrin β3及p-src蛋白相对表达水平的比较(n=3,±s)
组别对照组2 μg/L组20 μg/L组200 μg/L组F CK 1.00±0.00 0.86±0.10 0.59±0.06ab 0.55±0.10ab 21.467*MMP-9 1.00±0.00 0.73±0.06a 0.56±0.04ab 0.50±0.05ab 72.850*integrin β3 1.00±0.00 0.53±0.04a 0.51±0.06a 0.49±0.04a 121.361*p-src 1.00±0.00 0.83±0.14 0.49±0.14ab 0.36±0.08ab 22.341*
3 讨论
骨是一种矿化的结缔组织,破骨细胞和成骨细胞之间的信号传导对于骨重塑和骨微环境稳态具有重要意义[12]。破骨细胞通过空泡腺苷三磷酸(ATP)酶、补体3a、轴突导向因子4D及微小RNA等影响成骨细胞骨形成;成骨细胞通过骨保护素/核因子受体活化因子/核因子受体活化因子配体(OPG/RANK/RANKL)、核因子受体活化因子配体/G 蛋白偶联受体4/核因子受体活化因子(RANKL/LGR4/RANK)、促红细胞生成肝细胞激酶受体相互作用蛋白B2/促红细胞生成肝细胞激酶B4(EphrinB2/ephB4)和死亡受体/死亡配体(Fas/Fasl)等通路影响破骨细胞分化与凋亡[13]。破骨细胞特有的超微结构是完成其骨吸收功能的结构基础,通过重组细胞骨架形成极化构象,将细胞表面膜分为4个区域,即封闭区、褶皱缘、基底外侧区和功能分泌区[14]。极化的破骨细胞通过足质体黏附于骨表面,肌动蛋白环围绕褶皱缘形成相对独立的微环境,通过释放CK 和MMP-9 等水解酶实现骨吸收[15-16]。
柚皮苷又称柚柑、异橙皮苷,是中药骨碎补的重要成分。其化学本质是一种双氢黄酮类单体,分子式为C27H32O14,部分空间结构与雌激素类似,具有抗炎、抗氧化及改善微循环等功效。本研究中TRAP染色结果发现实验组中阳性细胞数显著降低,提示柚皮苷对破骨细胞的分化具有一定的抑制作用。qPCR 与Western blot 结果提示柚皮苷可以抑制CK与MMP-9 在破骨细胞中的表达。现已发现多种组织蛋白酶与破骨细胞功能相关,包括组织蛋白酶BE及组织蛋白酶G等,但是CK是骨吸收区酸性环境下发挥骨吸收作用的主要蛋白酶。同时,临床研究发现CK抑制剂可以有效降低骨吸收指标,提高患者骨密度[3]。MMP-9 是一种锌依赖蛋白水解酶,在哺乳动物胚胎期软骨内成骨及骨重建过程中起到胶原酶的作用,在细胞外基质重塑中发挥重要作用,是破骨细胞和血管内皮细胞侵入矿化组织所必须的生物因子,破骨细胞分化过程中,MMP-9表达上调,加速骨吸收[17]。Kim 等[18]在斑马鱼骨质疏松模型中发现,抑制MMP-9可以增加钙结节茜素红染色程度及椎体骨密度,改善骨质疏松程度。
目前许多研究证明柚皮苷可以抑制破骨细胞功能。Ang 等[19]发现柚皮苷可以抑制RANKL 介导的核转录因子(NF)-κB和细胞外调节蛋白激酶(ERK)信号通路,干扰破骨细胞的形成和功能。Li 等[20]证实柚皮苷通过下调抗凋亡蛋白B 淋巴细胞瘤-2(Bcl-2),上调促凋亡蛋白Bcl-2 相关X 蛋白(Bax)、半胱氨酸天冬氨酸蛋白酶3(caspase-3)及细胞色素C促进破骨细胞凋亡。Yu等[21]通过对骨吸收陷窝面积定量分析,发现柚皮苷对骨吸收有明显抑制作用。目前研究大多聚焦于柚皮苷对信号通路的影响,然而破骨细胞的极化是其发挥骨吸收功能的重要生理过程。本研究重点探讨柚皮苷对破骨细胞黏附与极化功能影响,发现2 μg/L 组和20 μg/L 组integrin β3与src的表达降低,提示柚皮苷可以抑制破骨细胞黏附和极化。近年来研究发现,integrin β3 和src 在破骨细胞黏附和极化过程中发挥着重要作用,integrin与含有精氨酸-甘氨酸-天冬氨酸序列的蛋白(RGD)结合是破骨细胞极化的先决条件[22]。Mo等[23]在研究牙龈炎时发现,牙龈炎导致integrin β3表达上升,增强破骨细胞骨吸收功能,使用integrin β3特异性拮抗剂西仑吉肽后,破骨细胞功能受到抑制。基于蛋白芯片技术研制的新型蛋白质抑制剂IPS-02001 可以抑制integrin 与骨桥蛋白结合,其抗骨质疏松作用已经在动物实验中得到验证,目前美国食品药品监督管理局(FDA)尚未批准integrin 拮抗剂类药物上市,但临床前评估已经取得了令人满意的结果[24]。1991年Soriano等[25]首次发现c-src缺失会导致破骨细胞功能不全。c-src 与integrin 在骨吸收过程中是相互偶联的。integrin αvβ3是integrin的一种异二聚体,脾酪氨酸激酶(Syk)是一种调节破骨细胞功能的酪氨酸激酶,位于免疫调节蛋白Dap12 和 免 疫 球 蛋 白FcRγ 下 游,细 胞 内c-src、integrin αvβ3及Syk形成三元复合物,通过自身磷酸化激活鸟嘌呤核苷酸交换因子(Vas3),将小G 蛋白超家族(Rho)蛋白转化成结合鸟苷三磷酸(GTP)的激活态,在肌动蛋白相关蛋白2/3(Arp2/3)复合体作用下形成肌动蛋白环,在突触结合蛋白Ⅶ协同下,肌动蛋白环收缩形成褶皱缘,完成细胞极化[15,26-27]。本研究初步发现柚皮苷可以在一定程度上抑制破骨细胞分化,降低CK、MMP-9、integrin β3、c-src 及psrc的表达,提示抑制破骨细胞分化与极化可能是柚皮苷抗OP 的机制之一。本研究的不足之处是仅观察了基因与蛋白水平的变化,尚未进行上游信号通路方面的深入研究。另外,本研究目的重点是分析干预的有效性,实验分组尚未筛选出抑制效果最显著的药物剂量,有待于后续研究进一步探讨。
猜你喜欢
中国老年学杂志(2022年19期)2022-11-21
现代财经-天津财经大学学报(2022年5期)2022-06-01
航天电子对抗(2022年2期)2022-05-24
中国饲料(2022年5期)2022-04-26
中国骨质疏松杂志(2022年1期)2022-02-17
北京航空航天大学学报(2021年9期)2021-11-02
山东医药(2020年36期)2020-12-31
农产品加工(2020年17期)2020-10-22
食品安全导刊·中旬刊(2020年2期)2020-06-01
航天电子对抗(2019年4期)2019-06-02
