
短串联靶标模拟技术及其在植物miRNA功能研究中的应用
2020-12-21 09:56郑文清张倩杜亮
生物技术通报 2020年12期
郑文清 张倩 杜亮
(北京林业大学生物科学与技术学院,北京 100083)
Small RNA包括微小RNA(miRNA)和小干扰RNA(siRNA),它们在植物的生长、发育、表观遗传、染色体完整、抵御病毒感染,以及对环境的改变作出响应中扮演着重要的角色[1-4]。其中,miRNA主要通过转录后水平发挥负调控功能[5],而siRNA在RNA干扰和转录基因的沉默中都发挥着重要的作用[3]。目前为止,随着许多物种的高通量测序的完成,很多物种的miRNA被鉴定出来[6]。尽管如此,研究miRNA的技术仍然相对匮乏[7]。传统的研究基因功能方法最典型的特点是依赖于突变体[8],但是这种方法不适用于miRNA的研究。其原因主要包括:miRNA基因较小、家族成员多、每个miRNA可能靶向调控多个下游基因等。在人们早期对miRNA功能的研究工作中,主要是通过过表达miRNA或者表达miRNA-resistant的靶标基因[9],达到调控miRNA功能的目的。但是,这两种方式都有缺陷:其中,由于每个miRNA都有多个靶标基因,过表达miRNA基因会导致其所有靶标基因表达水平的下降,因此这些方法不能很好地反映某个特定的miRNA的功能;而对于传统miRNA-resistant技术,同样存在靶向性不强的问题。
近年来随着分子生物学、遗传学、细胞生物学等学科的发展,开发了一种对miRNA功能研究比较有效的技术,即“短串联靶标模拟”(Short tandem target mimic,STTM)技术。STTM是一个人工构建的非编码RNA(100 nt左右大小),由两个miRNA互补序列和一段中间序列组成,它可以通过稳定的遗传转化或者病毒介导的瞬时表达系统在植物中表达[10]。通过检测一些影响STTM表达效率的参数,可以推导出设计最好的STTM的一般规则。与之前的几种技术相比,如“靶标模拟”(Target mimic,TM)[11]、“miRNA sponge”[12-15]和其他一些RNA引 诱 的 类 型 如“tough decoy(TuD)RNAs”[16-17],该技术可以在体内特异地诱导miRNA的降解,且已经成功应用于拟南芥[18]、烟草[10]、大豆[19]、番茄[20]、棉花[21]、小麦[22-23]和水稻[24]等的miRNA功能研究中[25]。本文将在介绍这个技术的基础上,重点对应用范围和优缺点等进行总结,以期为更好地利用该技术来研究植物miRNA的功能奠定基础。
1 STTM
1.1 STTM的发现
STTM最初是以“靶标模拟”现象的发现为基础建立的。早期研究发现,当植物处于磷饥饿状态时,miR399和一个非蛋白编码基因IPS1(INDUCE BY PHOSPHATE STARVATION 1)被诱导表达,与此同时,miR399靶基因PHO2的表达量显著地下降[11]。序列比对显示了IPS1的RNA和PHO2的mRNA都可以有效地结合miR399。PHO2mRNA当与miR399结合时,被特异地切割,进而抑制了PHO2的转录水平。但是,当IPS1表达量上升时,IPS1RNA与miR399发生竞争性结合,阻碍了miR399对PHO2mRNA的切割。因此,IPS1RNA有效地模拟(Target mimic)了PHO2mRNA,作为一个诱饵吸引了miR399进而保护了PHO2的mRNA不被切割降解[11]。进一步研究发现,IPS1 RNA与 miR399的结合序列不同于PHO2mRNA与miR399的结合序列,在其第10和11位序列之间加了CUA三个额外的碱基,形成了一个凸起的结构。这3个碱基使得IPS1RNA不能被miRNA399切割。运用IPS1RNA的这种特殊的结构,可以阻碍miRNA对mRNA靶序列发挥作用,间接保护靶基因,这种技术被称为“靶标模拟”。但是,并非所有的miRNA都能通过TM有效地被阻遏与靶基因的结合。例如,修饰过的IPS1RNA在植物中阻碍miR166/165的活性,效果不是特别显著[18]。这可能是由于修饰过的IPS1RNA的结合活性或者结构存在问题,或者是在植物发育到一定阶段时,被抑制的miR166/165又重新被激活。此外,miR166/165所在家族成员众多,这些成员由于序列上的相似性可能会与miRNA166/165竞争性结合IPS1RNA[18]。为了解决这个问题,早期的研究者对模拟序列的结构做了优化,并且发现,当同时引入了两个串联miRNA结合位点序列时,模拟结合的效率大大提高,由此建立miRNA靶标模拟技术[18]。
1.2 STTM的结构的设计
STTM含有两段不能被切割的miRNA结合位点,中间由一段spacer序列连接(图1)。这两段miRNA结合位点可以是相同的,靶标一个或者一类miRNA;或者不相同,靶标同一家族的不同miRNA。为了使STTM达到一个比较好的效果:第一,不推荐使用两个不同家族的完全不同的miRNA,这是因为一段miRNA结合位点不能够有效地靶标一个miRNA。第二,miRNA结合位点的第10和11个碱基(成熟miRNA的5'端)之间应该有一个CUA三碱基的凸起,这样可以保证STTM序列不被miRNA切割。一些miRNA可能恰巧在第10位碱基之后有可以和STTM结构的CUA互补的UAG结构,这可能导致突起位置的转换和STTM的断裂。在这种情况下,应该使用其他的三碱基结构。第三,spacer序列的长度最好是从48-88 nt,且是富含AT,以便形成一个相对稳定的结构。
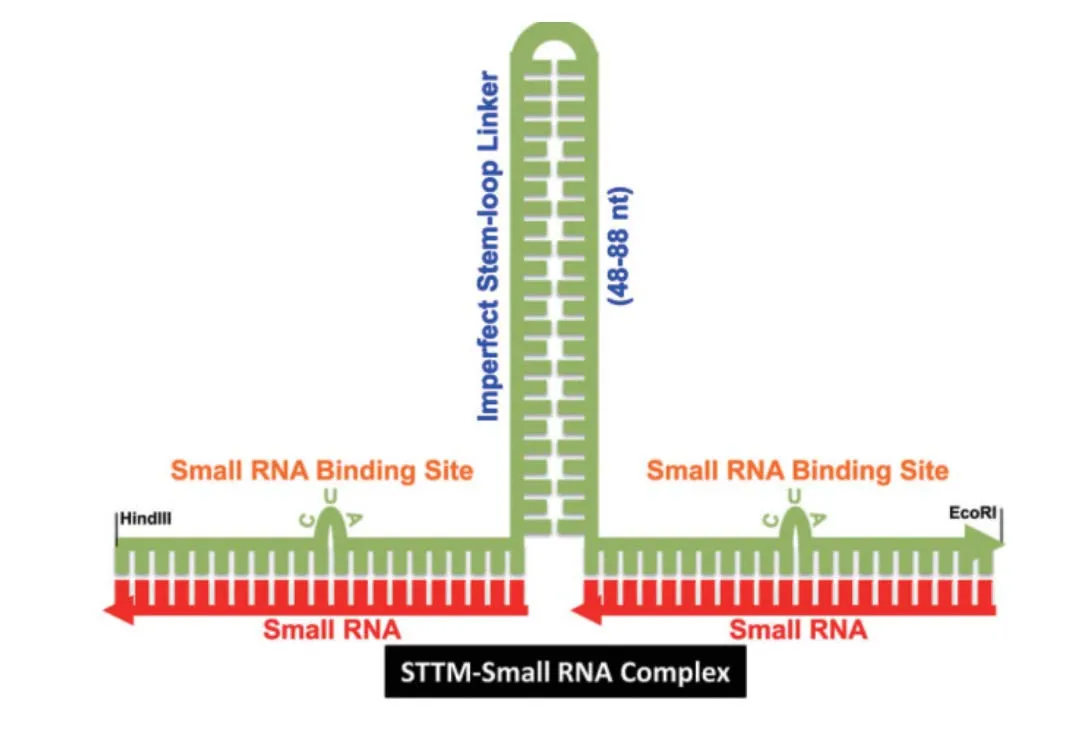
图1 STTM结构示意图[26]
1.3 影响STTM效率的因素
1.3.1 Spacer的序列长度 STTM由两个miRNA结合位点和一段spacer序列构成。这段连接序列可能可以控制细胞内的Dicer酶的活性,因此对于稳定STTM的结构起着至关重要的作用。并且这段中间的连接序列的长短对于miRNA调控靶基因有着比较重要的作用[27-29]。在拟南芥中,分别构建了STTM165/166-8、31、48、88和96 nt(后面的数字表示spacer碱基序列的长度)并导入植物中,通过表型变化的严重程度观察对miR165/166功能的阻碍作用。结果表明,当spacer序列长度为8 nt时,STTM转基因植株表型与野生型相似;当增加spacer序列到31 nt、48 nt和88 nt时,随着spacer序列长度的增加,生长抑制的表型逐渐加重,而spacer序列为96 nt时,转基因植株生长抑制的表型不再加重,表明spacer比较合适的长度是88 nt[24]。由此可见,STTM中spacer序列长度对miRNA的抑制效果有显著影响。
1.3.2 STTM热稳定性和次级结构 Yan等[18]分析了STTM165/166-8、31和48的RNA次级结构。通过RNA折叠分析表明,STTM165/166-8 RNA的结构有-211.7 kcal/mol的dG,相当不稳定。STTM165/166-31和48预测有低于-20 kcal/mol的dG,比STTM165/166-8有更高的稳定性。因此推测STTM165/166-8的效率低是因为它的低稳定性。STTM165/166-31的稳定性比48更高,但是它们的效率恰好相反。对它们的折叠结构的分析结果表明,STTM165/166-48形成了一个比较稳定的茎环结构。总的来说,这些分析表明了STTM转录本的热稳定性和次级结构都会影响STTM的效率,在设计时要综合考虑。
1.3.3 启动子选择 在个体中表达STTM序列时,可以选择组成型、组织特异型或者诱导型表达启动子。大多数情况下,为了研究miRNA在植物和动物中的全部功能,组成型表达STTM可以揭示出miRNA缺失功能在不同的发育阶段的影响。在这种情况下,RNA聚合酶II如35S或者2×35S、CMV/EF-1a和polⅢ启动子如U6都可以使用[16,23-24,26]。为了更加准确地研究miRNA的功能,需要组织特异性的或暂时的表达STTM,在这种情况下,可以使用特异的启动子。但是,当选用组织特异型或者诱导型启动子时,要注意启动子效率对STTM调控效果的影响[18]。
1.4 参与STTM作用过程的核酸外切酶
SDN蛋白家族的核酸外切酶参与了small RNA的降解,可能参与了STTM介导的miRNA降解[30]。为了证实这一想法,2012年Yan等[18]分别在野生型和sdn1-1 sdn2-1拟南芥双突变体背景下表达STTM165/166-48并观察其对植株表型的影响。结果显示95%的含有STTM165/166-48的双突变体没有表现出明显的表型变化,而90%的含有STTM165/166-48的野生型植株都表现出了明显的表型。qRT-PCR结果显示,两种背景下的STTM165/166-48的转录本表达水平相似,但是,突变体背景下的miR165/166的表达水平比野生型的高很多。同时,在sdn1-1 sdn2-1双突变体背景下表达STTM156/157-48,表型变化也出现类似的结果。这些研究表明,SDN蛋白家族是STTM得以发挥作用的重要核酸外切酶[18]。
2 STTM与TM和SP技术比较
MiRNA功能的鉴定通过之前传统的缺失功能的方法被证明是比较困难的,因为大多数miRNA家族成员具有功能冗余性。在miRNA沉默技术中,STTM与TM和SP较为相似,都是通过降低内源的miRNA表达水平发挥调控作用,但又各有特点。
2.1 TM原理以及应用
2007年,Franco-Zorrilla等[11]在研究拟南芥低磷胁迫响应时发现了TM现象,并利用该技术研究了miRNA的一些功能。在该研究中,PHO2是miR399的靶基因,miR399通过剪切PHO2基因进而影响基因的表达。IPS1可以与miR399配对结合,进而影响了miR399与PHO2的配对结合,增加了PHO2基因的表达,维持了体内的磷平衡。进一步的研究发现,IPS1的结构即是在miR399的第10和第11个碱基之间多了3个碱基(CUA),正是由于这3个碱基造成了IPS1能够与miR399结合但是不能被miR399切割(图2)。同时,他们根据IPS1的结构又构建了miR156和miR319的TM载体,并且转化了野生型拟南芥,拟南芥表现出比较明显的表型,miR156和miR319的表达量有一定程度的下降。Todesco等[31]利用TM 技术构建了一系列miRNA的载体,进行了拟南芥的遗传转化,为研究miRNA的功能奠定了一定的理论基础。
2.2 Sponges(SPs)原理及应用
在前人的研究中,证明“miRNA sponges”在动物中对miRNA的抑制效果比较有效[12]。SPs是包含了多个感兴趣的miRNA结合位点的合成转录本,它的工作原理主要是作为miRNA的诱饵,阻碍目标miRNA与其靶标mRNA的结合,进而增强mRNA的表达。每个SP内部含有2个碱基的错配,并且每个miRNA结合位点中间含有4个碱基的连接序列(图3)。SP中多个结合位点和多个连接序列增强了SP结构的稳定性。并且由于SP结构序列较长,因此构建和转化比较困难。SP主要运用在动物中,在植物中的研究较少。Reichel等[33]在植物中构建了cmSP165/166 和cmSP159载体,转化拟南芥植株,产生了比较明显的表型。
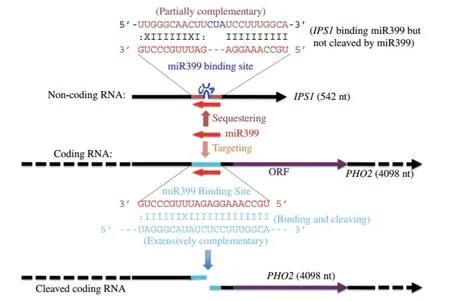
图2 IPS1介导的miR399的TM工作原理示意图[32]
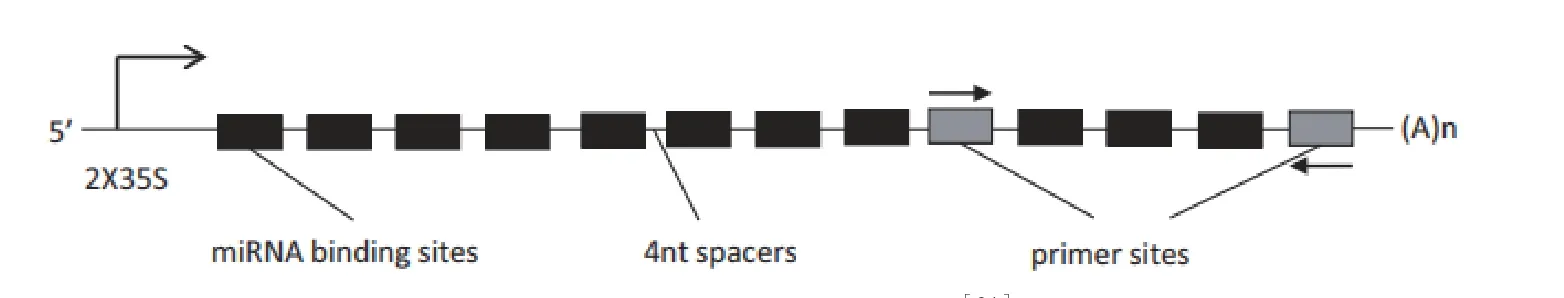
图3 SP技术的构建示意图[34]
2.3 STTM与TM、SP技术的比较
STTM与TM、SP技术的比较首先从其结构上来说,STTM包含两个miRNA结合位点,每个miRNA结合位点的第10和第11个碱基之间多出3个额外的碱基(CUA),连接序列较长(48-88 nt)。TM包含一个miRNA结合位点,并且miRNA结合位点的第10和第11个碱基之间也多出3个额外的碱基(CUA),没有连接序列,而SPs通常包含4-15个miRNA结合位点,每个结合位点内部含有2个碱基的错配,连接序列较短(4 nt)。其次,STTM与TM、SP技术从其作用机制上来说,都是RISC和RNA相互作用诱导miRNA的降解(表1)。

表1 TM、SP和STTM技术的比较
STTM、TM和SP这3个技术均有其优点和缺点。由于STTM的结构较短,因此构建比较容易,并且由于STTM比TM多一个结合位点,因此效率较高,特异性较强。另外,STTM技术介导的miRNA的降解过程不可逆,阻碍miRNA的表达较显著,转基因植株表型稳定[18,35],但是STTM技术不适用于研究低表达丰度的miRNA。TM技术由于其结构较短,因此构建也比较容易,但是由于其只有一个miRNA结合位点,因此不适用于有多个成员的miRNA家族。研究表明,SP技术在动物中的沉默效果较好,但是由于其结构较长,因此构建比较困难[12]。
3 STTM在植物中的应用
3.1 STTM在miR165/166研究中的应用
MiR165/166是植物中最丰富的miRNA家族之一,并且影响着包括叶、花、根和维管组织的许多发育过程。在拟南芥基因组中有2个miR165基 因(miR165a、miR165b)和7个miR166基 因(miR166a-g)[36]。成熟的miR165和miR166基因只在第17个碱基上有C/U的不同。在拟南芥中过表达miRNA165a、miR165b或者miR166a都有相似的表型,说明了它们的功能具有冗余性。目前研究发现,miR165/miR166的靶基因包括HD-ZIPⅢ家族的5个成员,分别是PHABULOSA(PHB)/ ATHB14、PHAVOLUTA(PHV)/ ATHB9、INTERFASCICULAR/FIBERLESS/REVOLUTA(IFL1/REV)、INCURVATA4/ CORONA / ATHB15和ATHB8[34]。这些基因调控许多植物发育过程,如顶端分生组织和侧生分生组织的形成、叶片的极性、维管组织形态和花的发育等过程[37-43]。
在拟南芥中,过表达STTMmiRNA165/166植株的表型与PHB基因的显性抑制突变体phb-1d相似但更明显。Small RNA免疫印迹和深度测序都证明了在STTM165/166转化植株中,超过95%的miRNA165/166被 降 解[18]。拟 南 芥 中 过 表 达STTM160和STTM165/166影响叶片的发育和干旱抗性[44]。在水稻中过表达STTM166,植株叶片卷曲,并且叶片有较小的泡状细胞和不正常的后壁组织细胞,气孔导度和蒸腾速率降低,这些表型与过表达miR166-resietent的OsHB4基因的表型是相似的[45]。另一个研究表明,在水稻中导入STTM165/166 后,转基因植株表现出叶形弯曲、植株矮化等表型[46]。在番茄中,过表达STTM165/166-31,亦能显著降低内源的miRNA165/166水平,同时增强其靶基因的表达水平[47]。在棉花中,过表达pCLCrVASTTM165/166,导致miRNA165/166表达降低,并造成棉花叶片的卷曲[48]。
3.2 STTM在其他miRNA中研究的应用
2017年,Zhang等[24]通过STTM在水稻中产生了35个miRNA家族的转基因株系,以期改良农作物的性状。在STTM转基因株系中,表现出了许多有价值的农业性状的改变,包括株高,分蘖数目,谷物数量,并且在五代以上都是稳定的。其中,STTM398能够增加花序长度,谷物的数量和大小;STTM172的水稻转基因株系相比野生型有非常短的茎,导致了封闭的花序和矮小的植株,而且相比野生型有更短的和密度更大的花序;STTM156的转基因植株根的发育受到影响,表明miRNA156参与了根的发育调控[24]。除此之外,在水稻中还产生 了STTM159、STTM160、STTM171、STTM441和STTM1428株系,且均产生了比较明显的表型[24]。在烟草幼苗中,Liu等[49]通过ZMBJ-CMV-2bN81-STTM载体降低了Nbe-miR165/166和Nbe-miR159的表达,并产生了较明显的表型。在玉米幼苗中,利用同样的载体降低了zma-miR167 和 zma-miR482的表达,分别产生了侧根数目减少和生长迟缓的表型。在番茄中,过表达STTM396的转基因番茄果实及萼片显著变大,miR396a和miR396b的表达量都显著下调,表明miR396对于番茄果实及萼片的大小有调控作用[50]。在番茄中利用STTM技术降低了miR482b的表达,表明了miR482b对番茄抵抗致病疫霉的负调控作用[51-52]。STTM9678转基因小麦增强了种子的萌发率,揭示了miR9678通过产生阶段性的siRNA和调控ABA或者赤霉素信号来影响小麦种子的萌发[53]。在蒺藜状苜蓿的根中,利用STTM技术对miRNA进行了失活[54]。STTM393的转基因大豆表现了对大豆疫霉菌的超敏感,并且异黄酮生物合成基因的表达也急剧下降[55]。在大豆中过表达STTM1507a、STTM1507c、STTM482a、STTM403a、STTM168a、STTM162b和STTM1515a,增强了大豆对花叶病毒的抵抗力[56]。STTM472a转基因杨树揭示了miR472a参与到对坏死真菌的抗性中[57]。STTM164转基因杨树相比野生型表现出木质部加厚的表型,表明miR164在杨树次生细胞壁合成过程中的作用[58]。干旱处理的菜豆中过表达STTM1514a,其靶基因NAC700的表达量上调,并降低了NAC诱导的phasiRNA的表达[59]。大麦中VIGS介导的STTM9683,miR9863a/b.2 和 miR9863c/b.1的表达量降低,Mla1基因表达量上升[60]。
4 展望
miRNA在植物的生长发育过程以及逆境胁迫响应中发挥着比较重要的作用,近年来随着分子生物学技术的发展,很多植物的miRNA被鉴定出来,因此研究miRNA的功能显得尤为重要。研究表明,STTM技术在研究植物miRNA及其家族基因功能中有操作简便、靶向性强等显著优势,是未来植物miRNA功能研究的有效技术。并且,由于STTM的结构比较小,因此可以将STTM串联起来诱导多个miRNA的降解,这将有力推动miRNA调控网络功能以及miRNA之间互作的研究工作。同时,在实际应用中,由于STTM序列的稳定性受多个因素控制,如spacer序列的长度,RNA的热稳定性、次级结构等。因此,选择一种比较稳定的STTM结构对研究miRNA的功能显得尤为重要。研究表明,48-88 nt长度的spacer序列是最理想的,太长序列的spacer可能对STTM的稳定性起不到比较明显的作用,反而会增加转化等的困难,太短长度的spacer序列可能不利于STTM结构的稳定。而且在启动子的选择上面可能也需要根据不同的miRNA表达情况来选择合适的启动子。此外,植物的MiRNA家族成员比较多,并且成员间往往表现为功能冗余;因此,在使用STTM技术对目标MiRNA家族进行表达干扰时,一般需要清楚地知道目标植物的基因组信息,以便进行精确和恰当的STTM序列设计。与此同时,植物的转基因效率和载体表达活性影响着STTM的实际效果,这也是在使用STTM技术时需要考虑的。
猜你喜欢
世界科学技术-中医药现代化(2022年9期)2023-01-17
军民两用技术与产品(2021年10期)2021-03-16
教学考试(高考生物)(2020年6期)2020-11-23
食品与生物技术学报(2020年8期)2020-01-06
世界农药(2019年3期)2019-09-10
学苑创造·B版(2019年5期)2019-06-14
科学24小时(2019年5期)2019-06-11
上海农业学报(2017年3期)2017-04-10
肿瘤影像学(2015年3期)2015-12-09
红领巾·探索(2015年9期)2015-09-10
