
接种牛粪菌系对稻秸发酵特性及固液相菌群的影响*
2020-12-30 09:19邓玉营黄振兴阮文权郝卫强
环境卫生工程 2020年6期
邓玉营,黄振兴,阮文权,郝卫强
(1.常州工程职业技术学院,江苏 常州 213164;2.江南大学环境与土木工程学院,江苏 无锡214122)
1 引言
厌氧发酵被认为是农作物秸秆资源化的重要途径之一,但秸秆结构复杂,水解过程被称为厌氧发酵的限速步骤。因此,高效接种物有利于提高稻秸发酵效率,常见接种物包括沼液沼渣、堆肥、活性污泥等[1]。单一秸秆原料含氮量低,无法满足厌氧微生物营养需要,导致其降解效率不高,因此在工程应用中,常添加畜禽粪便等来提高秸秆产甲烷效率,研究大多关注共发酵原料配比[2]、C/N 调控[3-4]、提高接种量[5]等。产甲烷酶活性及菌群结构的变化等指标有利于揭示体系发酵效率提高的机制。如在猪粪和玉米秸秆共发酵试验中,甲烷菌群丰度的提高是甲烷产率强化的关键[6]。Wang 等[7]通过菌群结构分析了猪粪和玉米秸秆共发酵中与微生物的相互作用,结果发现,单一原料发酵菌群结构易受到挥发性脂肪酸(volatile fatty acids,VFAs) 的影响,协同作用下降;添加猪粪提高了甲烷丝菌属(Methanosaeta)的相对丰度,从而提高了甲烷产率。在猪粪与稻秸共发酵试验中,接种量提高改变甲烷菌群结构,从而提高了甲烷产率[8]。在牛粪与玉米秸秆序批式共发酵试验中,工艺操作通过改变细菌结构发挥作用;但甲烷八叠球菌属(Methanosarcina) 受影响较小,与产甲烷效率正相关[5]。由此可见,畜禽粪便中含有的菌系是提高秸秆厌氧共发酵效率的关键[4],然而只将牛粪菌系作为接种物来研究菌群结构对稻秸发酵体系影响的报道并不多。
秸秆整个发酵过程是固液相微生物共同作用的结果。固相上水解菌群的变化,会导致酶活性的差异,影响秸秆结构的降解[9-10]。水解后产生的VFAs 进入液相,能通过互营氧化产生乙酸、氢等,该过程的氧化菌主要隶属于互养菌门、厚壁菌门和变形菌门[11],产物最终被甲烷菌所利用。然而除了对猪粪与小麦秸秆共发酵过程沼渣沼液中微生物变化有过研究外[12],很少有通过固液相菌群分析揭示秸秆发酵机制的报道。
为此,本研究设计牛粪菌系接种的批次试验,对稻秸厌氧发酵特性进行评价,分析产甲烷量分别与水解酶活性、单个VFAs 浓度变化之间的关系。并利用MiSeq 测序技术,分析固液相微生物结构特征,讨论特定功能种属变化对秸秆水解、发酵产酸、互营氧化及产甲烷的影响。以期为牛粪菌系接种稻秸发酵过程中微生物强化机制研究提供参考。
2 材料与方法
2.1 接种物和原料
接种污泥取自处理餐厨垃圾的厌氧反应器中,通过连续投加少量葡萄糖活化,提高产甲烷活性。总固体(total solids,TS) 质量分数为14.80%,挥发性固体(volatile solid,VS) 质量分数为6.65%。新鲜牛粪取自奶牛养殖场,为了减少对稻秸成分测定及产气的影响,通过两层纱布滤掉固渣后获得牛粪菌系,TS 质量分数为15.97%,VS 质量分数为13.68%。稻秸在50 ℃干燥箱中烘至恒量,粉碎后过40 目筛,TS 和VS 分别为98.46%和91.58%,保存于真空干燥袋中备用。
2.2 试验设计
接种污泥、牛粪菌系和稻秸的质量分数分别为8%、2%和2%,对照中不添加牛粪菌系。因为接种物没有经过脱气处理,分别设置了不投加稻秸原料的空白试验。体系利用改进的BY 提供金属营养元素[13],主要包括1.00 mg/L FeSO4·7H2O、1.00 mg/L CoCl2·6H2O、0.30 mg/L NiSO4·6H2O 等微量元素。添加K2HPO4·3H2O(1.34 g/L)、KH2PO4(0.51 g/L) 和NaHCO3(5.00 g/L) 组成缓冲体系,使初始pH 维持在6.8~7.1。
试验分别在厌氧瓶和甲烷潜力测试系统(AMPTS) 中进行,有效体积分别为80、250 mL,39 ℃条件下发酵44 d。每个体系设置11 个厌氧瓶作为平行试验,沼气通过铝箔袋收集。每隔4 d 各取1 个厌氧瓶,沼液用于水解酶活性、VFAs、pH指标测定,沼渣用于计算干基质量降解率。AMPTS 系统产生的沼气通过4 mol/L 氢氧化钠溶液吸收后测量甲烷体积,扣除空白试验的数值即为每天产甲烷量。发酵结束后沼渣沼液用于秸秆组分和固液相微生物分析。
2.3 指标分析
2.3.1 常规指标
样品用两层纱布过滤后,固渣TS 在105 ℃下进行测定,通过厌氧瓶中TS 的变化计算干基质量降解率。秸秆和固渣中纤维素、半纤维素及木质素含量用A2000i 纤维分析仪测定,残渣均需在65 ℃干燥箱中烘至恒量,通过发酵前后秸秆各组分变化计算纤维素、半纤维素降解率及木质素变化率,为了减少分析误差,添加接种物后再测定秸秆各组分质量分数。沼液水解活性通过纤维素酶或木聚糖酶活性评价,参考以前的方法[14],每毫升上清液释放1 μg 葡萄糖或木糖的酶量规定为1 个酶活力单位(U)。在12 000 r/min 下离心10 min,所得上清液用于指标分析,每个指标重复3 次。VFAs 使用GC-2010 Plus 气相色谱测定,pH 则通过DELTA 320 pH 计测定。
2.3.2 MiSeq 基因测序
AMPTS中沼液在800×g 下离心15 min,沉淀为固相样品;上清液继续在27 000×g 下离心30 min获得液相样品。使用Power soil®DNA 试剂盒提取初始接种物和固液相样品DNA,引物对338F 和806R 构建扩增文库后[15],利用MiSeq 平台完成测序。原始序列经加工处理,提交到NCBI,序列号为:SRP123575 (SRA),并在Silva 16S rDNA 数据库中比对,进行物种注释及菌群分析。
3 结果与讨论
3.1 产甲烷分析
产甲烷量日变化情况如图1 所示。对照体系最高产甲烷量出现在第1 天,为122.80 mL/d,然后迅速降低。牛粪菌系添加体系有两个产甲烷峰,分别出现在第4 天和第38 天,最高产甲烷量为120.35 mL/d。通过计算,甲烷产率(以VS 计) 为269.32 mL/g,相比对照体系的199.83 mL/g,提高了35%(表1)。

图1 产甲烷量日变化情况

表1 稻秸发酵结束后甲烷产率、干基质量降解率及秸秆组分变化情况
稻秸水解阶段是整个发酵的限速步骤,而牛粪中含有的水解微生物有利于稻秸复杂结构的降解[1]。添加牛粪菌系前2 天受到短暂抑制,随后出现了两次水解过程,对应两个产气峰,稻秸甲烷产率(以VS 计) 达到269.32 mL/g。Mussoline等[1]认为,即使通过各种物理化学预处理,稻秸甲烷产率(以VS 计) 也大多在92~280 mL/g 变动。在稻秸干法连续发酵中,张敏等[4]利用鲜牛粪构建厌氧微生物体系,在稻秸有机负荷4 kg/(m3/d)时,甲烷产率(以VS 计) 最高达到了227.8 mL/g,但试验使用了“黄储”破碎秸秆并添加猪粪为氮源。齐利格娃等[2]发现猪粪与稻秸共发酵时存在协同效应,比例为2∶1 时所有原料甲烷产率(以VS 计) 也只有213.4 mL/g;而单稻秸原料甲烷产率(以VS 计) 仅为124.7 mL/g,小于本试验对照体系。由此可见,本研究牛粪菌系接种稻秸产甲烷具有明显成本优势。
3.2 稻秸降解酶和水解酶活性
甲烷的产生来自稻秸的水解产物,因此甲烷产率的提高与底物降解率和各组分变化密切相关。如表1 所示,牛粪添加样品的干基质量降解率为41.79%,高于对照体系的39.32%。发酵结束后,固渣中纤维素和半纤维素的降解率分别为53.97%、56.14%,和对照相比,半纤维素降解相差不大,但纤维素的水解明显高于对照体系。木质素的苯丙烷结构降解过程需要氧气,在厌氧条件下难以降解[9],本研究发酵结束后木质素质量分数升高,变化率达353.77%。
纤维素酶和木聚糖酶活性变化如图2 所示。牛粪菌系接种体系出现了两个纤维素酶活性峰值,分别为26.54、32.68 U/mL,均值为18.82 U/mL;而对照中只在第8 天出现了峰值,之后快速下降,均值为15.10 U/mL。牛粪菌系接种体系中木聚糖酶活性均值为214.55 U/mL,明显高于对照的137.45 U/mL。

图2 纤维素酶和木聚糖酶活性的变化
稻秸的结构表明,纤维素成分被木质素和半 纤维素基质包裹在内部,阻止了水解酶的作用,而高活性的木聚糖酶降解半纤维素后暴露出微纤维结构,有利于纤维素的水解[16]。如在高粱厌氧发酵时,发现木聚糖酶活性峰值比纤维素外切活性提前,两类酶协同作用加快了高粱的水解[10]。Gu等[9]在接种牛粪稻秸发酵中,发现高活性的木聚糖酶提高了半纤维素降解率而且加快了纤维素的水解,使得沼气产率(以VS 计) 达325.3 mL/g。本研究中木聚糖酶活性较高,半纤维素先降解,在第4 天出现了第1 个产甲烷峰;随着稻秸结构的裂解,在发酵后期出现了纤维素酶活性峰值,纤维素降解,并在第38 天出现了第2 个产甲烷峰。
3.3 VFAs 浓度和pH 的变化
单个VFAs 的积累和消耗反映了水解发酵产酸与产甲烷之间的平衡。如图3 所示,对照体系中乙酸质量浓度在第4 天达到峰值3.69 g/L,随后逐渐降低。而牛粪菌系接种体系乙酸浓度在第4 天达到了4.35 g/L,说明开始阶段水解产酸的速度大于产甲烷,导致乙酸积累。发酵过程中出现两个峰值,质量浓度分别达到了4.95、3.89 g/L,36 d 后被完全利用。上述数据表明,牛粪菌系接种体系以产乙酸发酵为主,在短暂积累后被嗜乙酸型产甲烷途径消耗,其浓度与产甲烷活性紧密相关,该现象与一些秸秆发酵产甲烷的代谢类型一致[14,17-19]。

图3 发酵过程中单个VFA 浓度和pH 的变化
牛粪菌系接种体系开始阶段出现了丙酸积累,4 d 后达1.63 g/L,高于对照体系的1.12 g/L。最高浓度出现在第12 天,为3.65 g/L,20 d 后浓度下降为0,而丁酸则在16 d 后浓度下降为0。对照体系中丙酸和丁酸最高浓度大于接种体系,分别达4.42、1.98 g/L。一般认为,丙酸和丁酸是水解过程产生的还原氢来不及被利用的产物,只有通过互营氧化作用才能克服热力学障碍,而氧化菌生长缓慢,因此易导致体系的酸化[11]。本研究牛粪菌系接种开始阶段丙酸积累表明了嗜氢型产甲烷受到抑制,这也解释了体系直到第4 天才出现第1次产甲烷峰。同时这也是pH 在第20 天时降至6.73 的原因,但共接种物中含有的互营氧化菌群能利用丙酸和丁酸,使得pH 在整个发酵过程中均值维持在7.48,处于适宜的产甲烷范围[6]。
3.4 微生物结构特征
发酵结束后细菌结构在门水平上相对丰度的变化如图4 所示,其中厚壁菌门(Firmicutes)、互养菌门(Synergistetes) 和热袍菌门(Thermotogae)在发酵后固液相中都占主导地位。拟杆菌门(Bacteroidetes) 和WWE1 在液相中含量提高明显,而绿弯菌门(Chloroflexi)、变形菌门(Proteobacteria) 则在固相上的相对丰度增加。Sun 等[20]研究认为无论接种物来自于反应器还是活性污泥,纤维素或稻秸发酵体系中,Firmicutes 都占主导地位。其中梭菌目(Clostridiales) 中的种属,如梭菌属(Clostridium)、瘤胃球菌属(Ruminococcus)、毛螺菌属(Lachnospira)、丁酸弧菌属(Butyrivibrio)等,能在秸秆降解中发挥重要作用[21]。Synergistetes 能和嗜乙酸型甲烷菌形成互营菌群[22],牛粪菌系接种体系中该门菌属相对丰度的提高表明了互营氧化作用的强化。Thermotogae 能利用碳水化合物产VFAs[23],有利于发酵活性的提高。Spirochaetes中的螺旋体目(Spirochaetales) 则与水解木质纤维素、发酵产酸活性的增强有关[19]。液相中Bacteroidetes 菌属,如拟杆菌属(Bacteroides)、普氏菌属(Prevotella) 的出现既参与秸秆水解又能发酵产酸[24-25],从而提高了发酵代谢活性。而WWE1首先在处理活性污泥的厌氧反应器中被发现,能利用发酵产物产酸[26],在对照体系中相对丰度的提高与VFAs 积累紧密相关。固相中相对丰度提高的菌群中,Chloroflexi 主要是碳水化合物的降解菌[27]。Proteobacteria 如互营杆菌属(Syntrophobacter)、消化肠状菌属(Pelotomaculum) 等,常出现在活性污泥降解体系中,在互营氧化过程中发挥作用[11]。固液相菌群结构的分析有利于揭示厌氧发酵体系的强化机制[6]。

图4 发酵结束后细菌结构在门水平上相对丰度的变化
3.5 功能种属分析
3.5.1 固相功能菌群
固相中来源于接种污泥及牛粪菌系特定功能微生物的变化如表2 所示,Clostridium 是体系中的主要水解菌,对照体系丰度的升高易导致VFAs 的积累[5]。来源于牛粪菌系的高效纤维素水解菌瘤胃球菌(Ruminococcus)、纤维杆菌(Fibrobacter)在体系中的相对丰度高于对照,这也解释了牛粪菌系接种后稻秸纤维素水解酶酶活及降解率显著提高的现象。Chapleur 等[28]在纤维素的降解中,添加瘤胃液和市政废弃物污泥共发酵时,发现瘤胃水解菌从体系中消失,研究者归因于无法与污泥中功能菌竞争,这与本研究所得结论不一致。Lutispora 及沉积杆菌属(Sedimentibacter) 等属于多糖降解产乙酸菌[19,29],在牛粪添加体系中相对丰度的提高表明产乙酸代谢途径得到强化,提高了产甲烷效率。如Deng 等[14]在研究稻秸和瘤胃内含物共发酵时,发现固相上这两类菌属的含量提高,与产甲烷活性增强密切相关。
一些来源于污泥的部分菌属在牛粪菌系接种体系中含量降低或消失。如在沼气反应器中能降解各种多糖物质的Bacteroides[24],毛螺菌科(Lachnospiraceae) 中可溶性多糖降解菌Lachnospira[30],该类菌群的变化被认为与稻秸降解中不同水解微生物存在竞争有关[29]。
如表2 所示,牛粪菌系接种体系中甲烷八叠球菌属(Methanosarcina) 是主要类型,这和牛粪菌系接种中乙酸的积累有关。来源于牛粪的甲烷短杆菌属(Methanobrevibacter) 在发酵结束后相对丰度升高,达到了0.009 5%;而对照体系甲烷菌类型中鬃毛甲烷菌(Methanosaeta) 的相对丰度最高。在猪粪和稻草共发酵配比试验中,发现猪粪含有的易降解成分易导致VFAs 积累,降低了Methanobrevibacter 的相对丰度[2],与本研究甲烷菌的变化不一致。在青贮牧草降解试验中,Methanosarcina 被发现位于固渣表面,研究者认为由于纤维素水解产生的乙酸在固相上来不及扩散,乙酸浓度的升高增强了Methanosarcina 的附着生长[18]。在稻秸厌氧发酵试验中,通过残渣微生物形态和功能菌群分析发现水解菌和Methanosarcina共存在固相上时,水解效率大大提高[14]。李奥等[8]在猪粪与稻秸共发酵试验中,也发现了发酵结束后Methanosarcina 相对丰度显著提高。这也从甲烷菌类型上解释了牛粪菌系接种体系中嗜乙酸产甲烷代谢强化的现象。在猪粪与玉米秸秆共发酵试验中,Methanosarcina 占优势地位,但随着有机负荷升高,嗜氢型Methanobrevibacter 相对丰度有所提高,并获得高的甲烷产率[6],与本研究甲烷菌结构特点一致。研究表明,牛粪中存在的水解菌Ruminococcus、Fibrobacter 和 嗜 氢 型Methanobrevibacter 存在种间氢传递,当体系产生的氢被Methanobrevibacter 利用后,能够实现氧化型NAD+载体的重生,水解和产甲烷活性都得到提高[21]。如在柳枝稷发酵过程中,发现Methanobrevibacter对纤维素结构的水解起了重要作用[31]。而另一方面,嗜氢型甲烷菌活性如果受到抑制,会使水解微生物含量下降。如添加抑制产甲烷活性的溴乙基磺酸钠(BES),导致高效水解菌Ruminococcus 从小麦秸秆发酵体系中消失[32]。本研究中接种牛粪菌系提高了Methanobrevibacter 在体系中的相对丰度,有利于加快种间氢传递,使稻秸的纤维素水解和产甲烷活性都得到了提高。
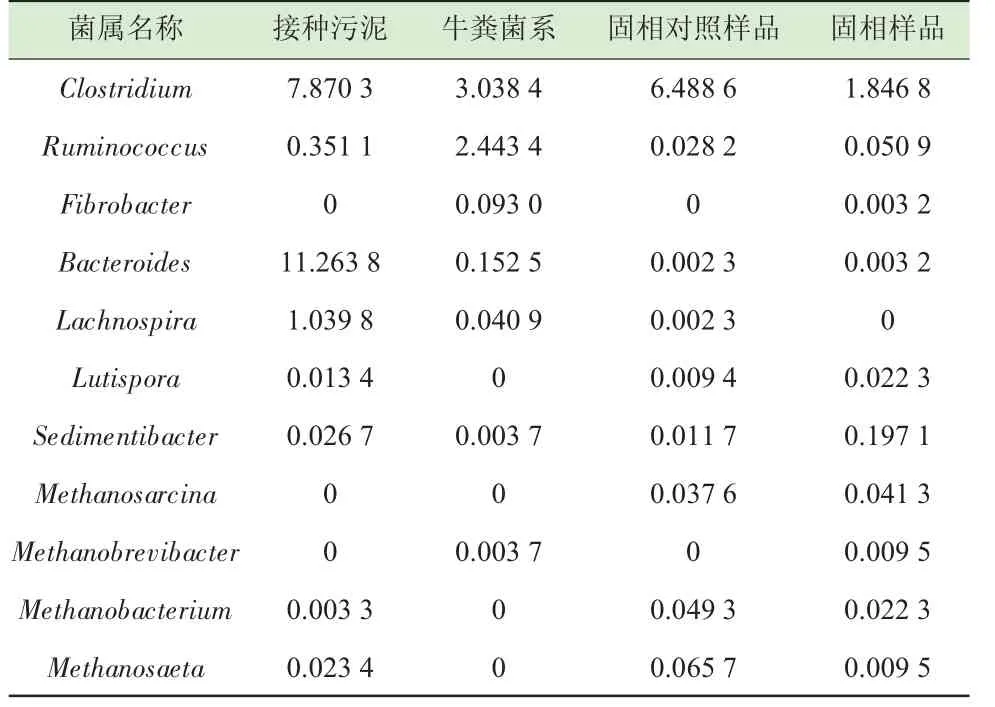
表2 固相上特定功能细菌和甲烷菌属相对丰度的变化 %
3.5.2 液相功能菌群
液相中丁酸弧菌属(Butyrivibrio)、密螺旋体菌属(Treponema)、Prevotella 等属于产酸微生物[19,21],和纤维素水解菌存在协同作用。如半纤维素降解菌Butyrivibrio、Treponema 可以和Ruminococcus、Fibrobacter 共同结合在稻秸和苜蓿上,首先裂解结构中的木聚糖成分,而后实现对纤维素结构的降解[33]。Prevotella 种属能参与半纤维素成分的降解,发酵后产生不同类型的VFAs[25]。如表3 所示。降解产酸菌的出现也解释了体系中木聚糖酶活性的提高。发酵结束后,对照体系中该类产酸菌属相对丰度较高,降解产生的丁酸、乳酸及琥珀酸积累,易导致体系酸化,反而抑制了产甲烷活性[17]。
来源于接种物的氧化菌能够和伴生菌形成互营氧化菌群,有利于降低VFAs 的浓度[11]。如表3所示,本研究中互营杆菌(Syntrophobacter) 和丙酸氧化菌(Pelotomaculum) 等主要存在于对照体系中;而氨基杆菌属(Aminobacterium)[22]及丁酸型互营单胞菌属(Syntrophomonas) 等在接种体系相对丰度较高,能通过互营氧化作用降低丙酸和丁酸浓度。但在猪粪添加小麦秸秆发酵的研究中,发现液相中存在Pelotomaculum,与Methanosaeta 形成互营菌群,增强了丙酸氧化效率[12],与本研究接种体系液相中互营菌类型不一致。液相中菌属的变化也进一步表明了体系中嗜乙酸产甲烷途径占主导优势,降低了丙酸和丁酸的积累。

表3 液相上功能菌相对丰度的变化 %
4 结论
1) 牛粪菌系接种稻秸发酵试验中,分别在第4 天和第38 天出现了产甲烷峰,稻秸甲烷产率(以VS计) 达269.32 mL/g,比对照体系提高了35%。稻秸的干基质量降解率达41.79%,纤维素和半纤维素的降解率分别为53.97%、56.14%。纤维素酶和木聚糖酶活性均值分别为18.82、214.55 U/mL。
2) 发酵结束后,细菌菌群结构的变化明显,固相上水解功能菌Ruminococcus、Fibrobacter 的相对丰度高于对照体系,产乙酸菌Lutispora、Sedimentibacter 相对丰度提高。Methanosarcina 是主要甲烷菌类型,Methanobrevibacter 相对丰度升高。在液相中,Treponema、Prevotella 等产酸微生物主要存在于对照体系中。Aminobacterium 和Syntrophomonas 等互营氧化菌在接种体系中相对丰度较高,而Syntrophobacter 和Pelotomaculum 是对照体系的主要类型。
猜你喜欢
意林彩版(2022年2期)2022-05-03
今日农业(2021年10期)2021-11-27
化工环保(2021年5期)2021-10-19
化学与生物工程(2021年8期)2021-08-26
粉末冶金技术(2021年1期)2021-03-29
江苏理工学院学报(2020年2期)2020-10-23
趣味(语文)(2020年11期)2020-06-09
中国沼气(2019年4期)2019-12-06
中学化学(2019年3期)2019-07-08
中学化学(2016年2期)2016-05-31
