
一株莱氏绿僵菌的筛选及其对草地贪夜蛾的毒力
2021-02-23 05:33龙秀珍高旭渊曾宪儒韦德卫江小冬于永浩
中国生物防治学报 2021年6期
龙秀珍,高旭渊,曾宪儒,何 瞻,韦德卫,江小冬,曾 涛,于永浩
(广西农业科学院植物保护研究所/广西作物病虫害生物学重点实验室/农业农村部南宁作物有害生物科学观测实验站,南宁 530007)
草地贪夜蛾Spodoptera frugipera,鳞翅目Lepidoptera,夜蛾科Noctuidae,原产于美洲热带和亚热带地区,具杂食性和迁飞性,可为害353种作物[1]、单雌平均产卵量436~1281粒[2],在风速或风向适宜时每晚可迁飞几百公里[3],是联合国粮农组织全球预警的重大农业害虫。截至2020年11月,草地贪夜蛾已在世界113个国家和地区发生为害[4]。在我国,自2019年1月云南省普洱市江城县首次发现入侵草地贪夜蛾幼虫为害[5],截止到2020年7月9日,我国共有21个省(市、区)1029县(区)发现草地贪夜蛾幼虫[6]。草地贪夜蛾严重为害我国的玉米等作物,2019年该虫为害玉米在云南曲靖市造成的直接经济损失达2902.67万元[7],预计我国每年因草地贪夜蛾为害造成的经济损失可达100亿元人民币[8],草地贪夜蛾对我国农业生产造成了严重威胁。
目前,草地贪夜蛾的防治主要依赖化学农药,但化学农药的长期使用容易使害虫产生3R问题,我国已发现该虫对传统的有机磷类、拟除虫菊酯类及氨基甲酸酯类农药化学杀虫剂产生了抗性[9],广东茂名种群对甲维盐和氯虫苯甲酰胺的抗性达中抗水平[10],加之化学农药的大量使用还会带来一系列的环境生态问题,国家出台了减药行动方案,这使得化学农药防治受到一定制约,必须寻找替代措施。昆虫病原真菌是一类能感染昆虫并在昆虫体表或体内增殖致使昆虫发病甚至死亡,可在昆虫群落中自然传播并造成流行的微生物。昆虫病原真菌对环境友好,对害虫具有持续的控制能力,不易导致害虫抗性,因此,昆虫病原真菌在草地贪夜蛾的防控中具有很大的应用前景[11]。莱氏绿僵菌Metarhizium rileyi,原名莱氏野村菌Nomuraea rileyi(Farlow)Samson,是一种重要的虫生真菌,关于利用其防治草地贪夜蛾的研究在国外已有不少报道[12-15]。国内郑亚强等[16]在云南省曲靖市玉米地发现了自然感病的草地贪夜蛾幼虫僵虫,经分离鉴定为莱氏绿僵菌,1.0×108孢子/mL菌悬液处理草地贪夜蛾3龄幼虫7 d的死亡率为100%;雷妍圆等[17]在广东省广州市野外采集到了自然感染莱氏绿僵菌的草地贪夜蛾幼虫僵虫,该菌1.0×109孢子/mL接种草地贪夜蛾2龄幼虫7 d的校正死亡率为100%,但莱氏绿僵菌对草地贪夜蛾其他龄期幼虫和其他虫态的致病力有待进一步研究。
我们从广西南宁市武鸣区的田间采集到被虫生真菌感染的草地贪夜蛾幼虫,经分离纯化获得1株病原真菌,采用形态学和ITS-rDNA序列分析相结合的方法对病原真菌进行了鉴定,并采用浸虫法研究了该菌对草地贪夜蛾的致病力,发现其以较低的浓度即可获得明显的效果,为草地贪夜蛾的微生物防治提供候选菌种资源。
1 材料与方法
1.1 供试虫源
草地贪夜蛾采集于广西南宁市武鸣区田间玉米植株上,在实验室用玉米叶饲养,待其子代孵化后作为供试虫源。室内饲养条件为温度(27±1)℃,相对湿度70%~90%,光周期14L:10D。
1.2 供试菌株
草地贪夜蛾幼虫僵虫采集自广西壮族自治区南宁市武鸣区田间,将僵虫置于5 mL离心管中带回实验室,在无菌条件下用接种针挑起少量孢子,采用划线法接种于萨氏麦芽糖琼脂酵母培养基(SMAY)(麦芽糖40 g、蛋白胨10 g、酵母浸膏10 g、琼脂18 g、蒸馏水1000 mL),于温度(26±1)℃、相对湿度(80±5)%、光周期12L:12D的人工气候箱中培养。培养7 d后,挑取少量孢子至新的培养基单孢分离纯化培养15 d,将分离纯化的菌株分生孢子置于20%的甘油中,于-80 ℃冰箱中保存备用,菌株编号为CDTLJ1,中国微生物菌种保藏管理委员会普通微生物中心保藏编号为CGMCC No.19608。
1.3 菌株培养特征观察
将分离纯化的CDTLJ1菌株在直径为90 mm的SMAY平板培养基上于温度(26±1)℃、相对湿度(80±5)%、光周期12L:12D的人工气候箱中培养,每天观察记录菌落形态特征;4~5 d后挑取菌丝,在光学显微镜下观察菌株的菌丝和分生孢子梗形态;10~12 d后挑取成熟分生孢子观察分生孢子的形态与大小。
1.4 菌株分子生物学鉴定
以分离纯化获得的 CDTLJ1菌株基因组 DNA为模板,采用真菌通用引物 ITS1(5′-TCCGTAGGTG AACCTGCGG-3′)和 ITS4(5′-TCCTCCGCTTATTGATATGC-3′)进行菌株 ITS-rDNA 序列 PCR 扩增。PCR反应体系 50 μL:2×Es Taq MasterMix(Dye)25 μL,DNA 模板 1 μL,上、下游引物各 2 μL(10 μmol/L),ddH2O 20 μL。扩增程序:94 ℃预变性 5 min;94 ℃ 30 s,56 ℃ 30 s,72 ℃ 30 s,进行 35个循环;72 ℃延伸5 min。PCR产物经1.0%琼脂糖凝胶电泳检测后送广州擎科生物技术有限公司测序。
1.5 菌株对草地贪夜蛾不同龄期幼虫、预蛹和蛹的毒力测定
前期的预试验表明1×107孢子/mL浓度对各龄期幼虫均有较好的毒力,且对预蛹、蛹也有致病力。本试验从SMAY平板上刮取菌株CDTLJ1孢子置于灭菌的0.05%吐温-80溶液中,用血球计数板计数得到的孢子浓度为1.20×107孢子/mL,配制成孢子悬浮液,用该浓度孢悬液进行试验。
选取大小一致、健康的草地贪夜蛾2~6龄各龄期幼虫、预蛹、化蛹12 h内的新鲜蛹,放入配制好的孢子悬浮液中,浸渍5 s后取出,置于无菌滤纸上吸干虫体表面多余水分,单头放入皿底垫有湿润滤纸的灭菌培养皿(d=7.5 cm)中。放置幼虫的皿内放入新鲜玉米叶供其取食,并每天更换新鲜的玉米叶。以浸渍0.05%土温-80无菌水溶液处理作为对照。每处理20头试虫,重复3次。处理后置于温度(26±1)℃、相对湿度(80±5)%、光周期14L:10D的人工气候箱饲养,每天观察记录死亡虫数,并将死亡的试虫继续保留在培养皿中保湿观察,确认是否为供试菌株感染致死。
1.6 菌株对草地贪夜蛾3龄幼虫的毒力测定
从SMAY平板上刮取CDTLJ1菌株孢子置于灭菌的0.05%吐温-80溶液中,配制成5个浓度梯度的孢子悬浮液(1×104、1×105、1×106、1×107、1×108孢子/mL);选取大小一致、健康活泼的草地贪夜蛾3龄幼虫,放入配制好的孢子悬浮液中,浸渍5 s后取出,置于无菌滤纸上吸干虫体表面多余水分,单头放入皿底垫有湿润滤纸的灭菌培养皿(d=7.5 cm)中,皿内放入新鲜玉米叶供其取食,并每天更换新鲜的玉米叶。以浸渍0.05%土温-80无菌水溶液处理作为对照。每处理20头幼虫,单头饲养,重复3次。处理后置于温度(26±1)℃、相对湿度(80±5)%、光周期12L:12D的人工气候箱饲养,每天观察记录死亡虫数,并将死亡的幼虫继续保留在培养皿中保湿观察,确认是否为供试菌株感染致死。
1.7 数据统计与分析
将CDTLJ1菌株的ITS序列在NCBI数据库中进行BLAST比对(https://blast.ncbi.nlm.nih.gov/Blast.cgi),序列经BLAST后下载同源性较高的序列,用MEGA X进行序列处理,应用最大似然法(ML),运行1000次bootstrap验证,构建系统发育树。
数据经Excel整理后,用 SPSS 17.0软件进行处理分析,统计各处理试虫的累积死亡率和累积校正死亡率。利用Duncan’s新复极差法对试验数据进行差异显著性分析,采用Probit方法计算致死中时(LT50),求回归方程及计算致死中浓度(LC50)。死亡率(%)=(处理死亡总虫数/处理总虫数)×100;校正死亡率(%)=(处理死亡率-对照死亡率)/(100-对照死亡率)×100。
2 结果与分析
2.1 菌株的培养形态特征及分子生物学鉴定
研究发现,菌株培养初期菌落较小,短绒毛状凸起,边缘呈黏液状,无分泌物,菌落正面呈乳白色,5~6 d开始产孢,产孢后菌落为淡绿色。菌丝光滑有分隔,分生孢子呈穗状,分生孢子梗产生于营养菌丝,每个分支上有数个分生孢子梗,分生孢子梗短圆柱形,近基部略膨大,顶部略短尖[16];分生孢子着生于顶部,链生,长椭圆形,表面光滑,聚集呈淡绿色,(3~4.5)μm×(2~3)μm(图1A)。菌落生长较缓慢,在SMAY培养基上培养10 d的菌落直径为15 mm(图1B)。
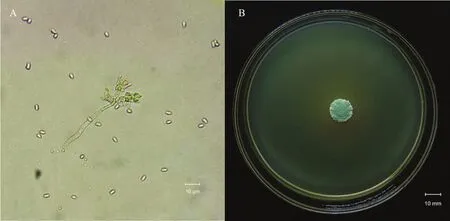
图1 菌株CDTLJ1的菌落和分生孢子形态特征Fig. 1 Morphological characteristics of colony and conidia of M. rileyi CDTLJ1
使用PCR扩增ITS-rDNA序列片段并测序,结果显示扩增片段为592 bp,将该序列在NCBI数据库中进行BLAST比对,发现该菌株与NCBI数据库中的Metarhizium rileyiNr Sf-1(GenBank登录号MN602591.1)菌株的相似性为100%。下载BLAST结果中同源性较高的序列,用MEGA X进行序列处理,经Clustal W多重对比,应用最大似然法(ML)构建系统发育树(图2)。由图2可知,菌株CDTLJ1与数据库中其他莱氏绿僵菌株聚为一大支,说明该菌株与数据库中的莱氏绿僵菌菌株具有较高的相似性。综合形态特征鉴定和ITS序列相似性分析,将该菌株确定为莱氏绿僵菌。
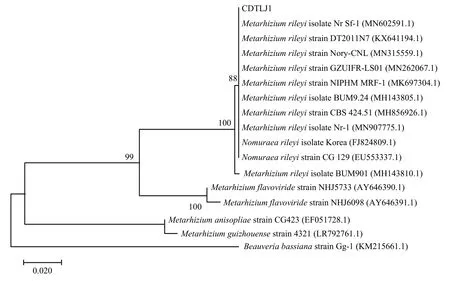
图2 基于ITS序列构建菌株CDTLJ1的ML系统发育进化树Fig. 2 ML phylogenetic tree of strain CDTLJ1 based on ITS sequences
2.2 草地贪夜蛾幼虫、预蛹和蛹感染莱氏绿僵菌CDTLJ1的症状
草地贪夜蛾2~6龄幼虫均可被莱氏绿僵菌CDTLJ1菌株感染。感染初期,幼虫取食行为、取食量、虫体外部形态与健康幼虫无差别;在接种处理后第4 d,各龄期幼虫均出现莱氏绿僵菌感染致死个体。以4龄幼虫为例,幼虫刚死亡时虫体发红、僵硬(图3A),随后体表逐渐长出大量白色菌丝(图3B),死亡后第3 d,虫体表面的菌丝开始产孢(图3C),死亡后第4 d,虫体被厚厚一层绿色分生孢子粉覆盖(图3D)。
部分草地贪夜蛾5龄幼虫被菌株CDTLJ1感染后不能正常化蛹(图3E),虫体逐渐僵硬并长出白色菌丝(图3F~G),随后产生绿色分生孢子(图3H)。化蛹12 h内的新鲜蛹也可被感染,部分6龄幼虫和预蛹在被感染后仍能化蛹,但蛹体2~3 d后逐渐变僵硬(图3I),保湿培养后菌丝从气门、节间等部位长出,并产生分生孢子(图3J~M),但被感染的预蛹和蛹体表长出的菌丝和孢子均比罹病幼虫产生的少。

图3 草地贪夜蛾幼虫、预蛹、蛹感染莱氏绿僵菌CDTLJ1的症状Fig. 3 External symptoms of larvae, prepupa, pupa of S. frugiperda after infection M. rileyi strain CDTLJ1
2.3 莱氏绿僵菌CDTLJ1对草地贪夜蛾不同龄期幼虫、预蛹和蛹的致死率
研究发现,莱氏绿僵菌CDTLJ1对草地贪夜蛾2~6龄幼虫、预蛹、蛹均具有致病力。幼虫、预蛹、蛹均在莱氏绿僵菌CDTLJ1 1.20×107孢子/mL处理后第4 d开始出现死亡,并随着处理时间的延长,草地贪夜蛾幼虫、预蛹、蛹的校正死亡率逐渐升高。处理后第4 d,2~6龄幼虫、预蛹、蛹的校正死亡率分别为63.33%、60.00%、60.00%、55.00%、43.33%、13.33%和8.33%。处理后第6 d,2~5龄幼虫的校正死亡率均达100%,6龄幼虫的累积校正死亡率为98.33%,预蛹和蛹的累积校正死亡率分别为28.33%、20.00%(图4)。
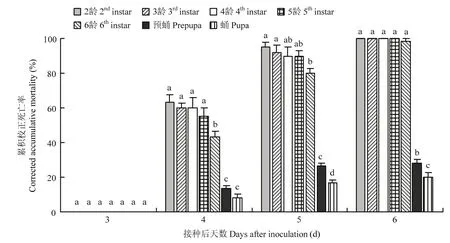
图4 莱氏绿僵菌菌株CDTLJ1对草地贪夜蛾的毒力Fig. 4 The virulence of M. rileyi strain CDTLJ1 against S. frugiperda
2.4 莱氏绿僵菌CDTLJ1对草地贪夜蛾不同龄期幼虫、预蛹和蛹的致死中时
在1.20×107孢子/mL浓度下,随着草地贪夜蛾虫龄的增加,草地贪夜蛾的致死中时(LT50)逐渐延长,即虫龄越大,试虫的LT50值越大。2~6龄幼虫的LT50值从3.91 d升至4.24 d;预蛹和蛹的累积死亡率均低于50%,因此无LT50值(表1)。
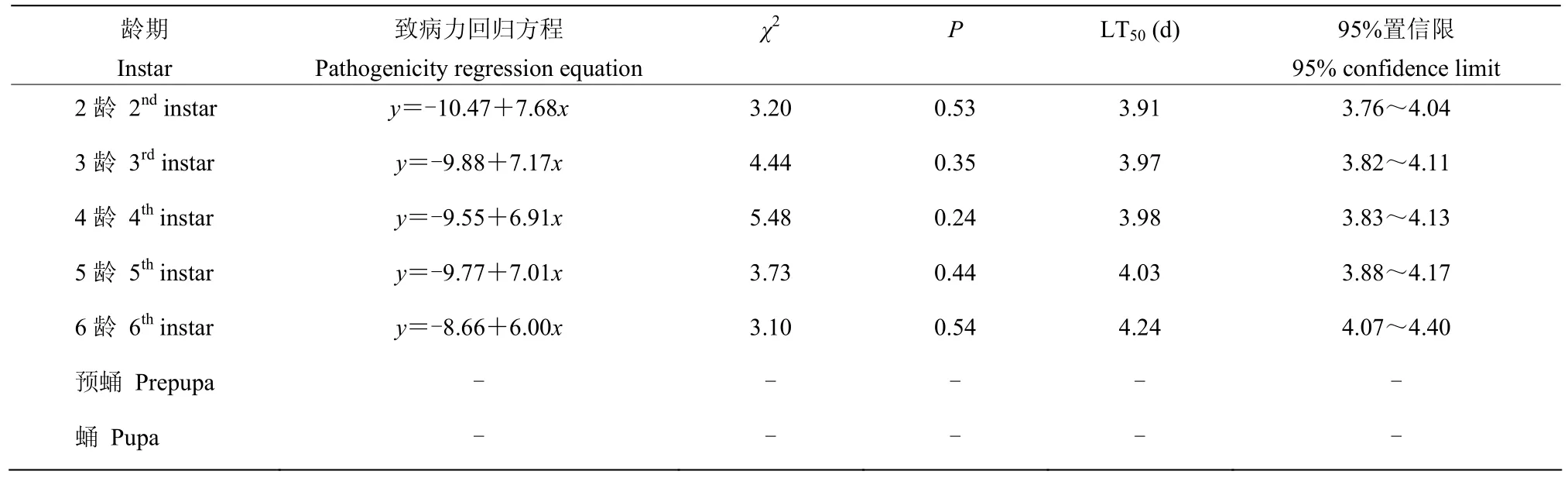
表1 莱氏绿僵菌菌株CDTLJ1对草地贪夜蛾不同龄期幼虫、预蛹和蛹的致死中时Table 1 The median lethal time of M. rileyi strain CDTLJ1 to S. frugiperda larvae, prepupa, and pupa
2.5 莱氏绿僵菌CDTLJ1不同孢子浓度对草地贪夜蛾3龄幼虫的毒力
莱氏绿僵菌CDTLJ1对草地贪夜蛾3龄幼虫具有的致病力较高,随着孢子浓度的增加和处理后时间的推移,草地贪夜蛾3龄幼虫的累积校正死亡率逐渐升高。孢子浓度1×105、1×106、1×107、1×108孢子/mL处理的3龄幼虫在处理后第4 d开始出现死亡,1×104孢子/mL处理的幼虫在处理后第5 d开始出现死亡,随后校正死亡率迅速升高,1×107、1×108孢子/mL处理的幼虫累积校正死亡率在处理后第6 d达到100%,1×105、1×106孢子/mL处理的幼虫累积校正死亡率在处理后第7 d达到100%,1×104孢子/mL处理的幼虫累积校正死亡率在处理后第8 d最高,为96.67%。应用Probit模型,得到莱氏绿僵菌CDTLJ1菌株处理草地贪夜蛾3龄幼虫后第6 d的致病力回归方程y=-3.73+0.92x(χ2=0.056,P=0.91),LC50为1.12×104孢子/mL,95%置信区间为4.48×103~2.06×104孢子/mL(图5)。
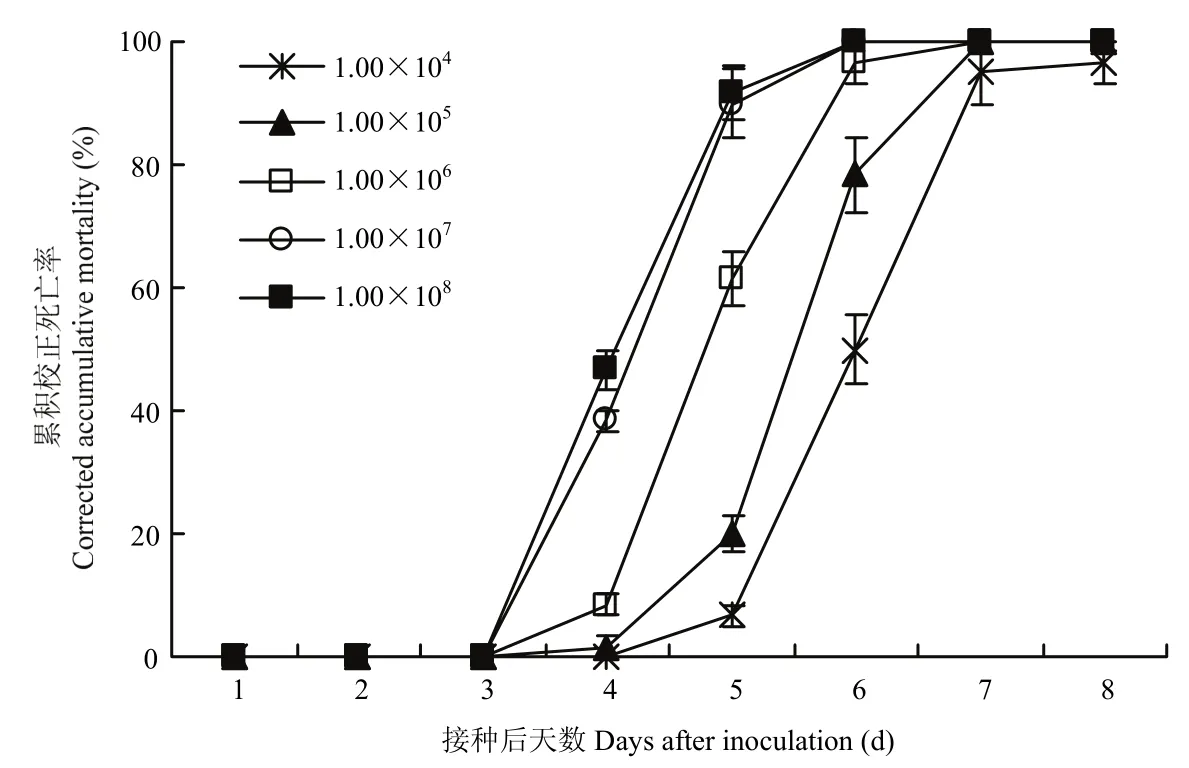
图5 莱氏绿僵菌CDTLJ1不同孢子浓度对草地贪夜蛾3龄幼虫的毒力Fig. 5 The pathogenicity of strain CDTLJ1 with different conidia concentrations against 3rd instar larvae of S. frugiperda
随着莱氏绿僵菌CDTLJ1孢子浓度的增加,草地贪夜蛾3龄幼虫的致死中时(LT50)逐渐缩短,孢子浓度在1×104~1×108孢子/mL范围内,LT50从5.93 d降至4.10 d(表2)。
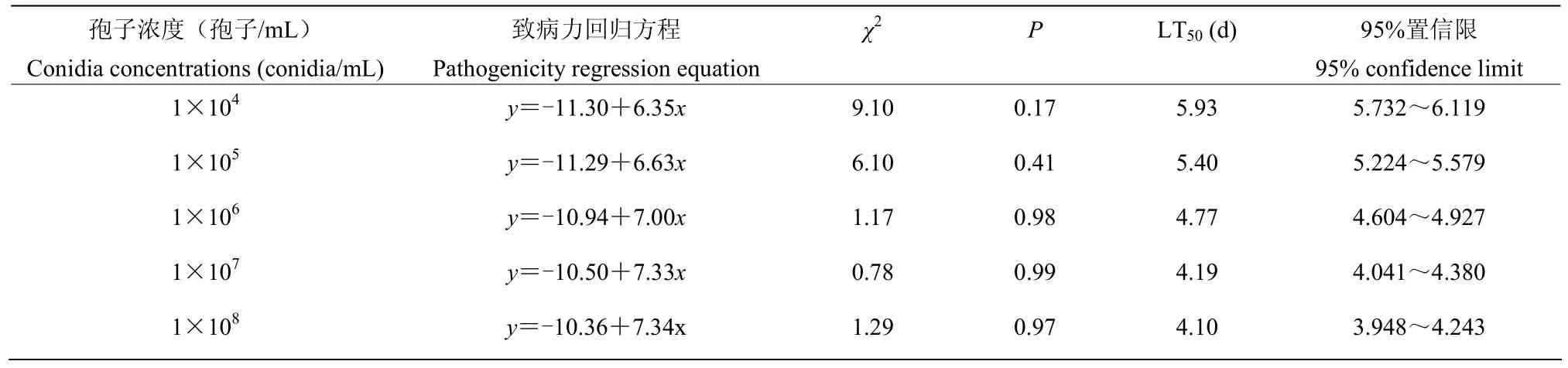
表2 不同孢子浓度下莱氏绿僵菌CDTLJ1对草地贪夜蛾3龄幼虫的致死时间效应Table 2 The median lethal time of M. rileyi strain CDTLJ1 to 3rd instar larvae of S. frugiperda under different conidia concentrations
3 讨论
报道的大部分感染草地贪夜蛾的病原真菌菌株仅对草地贪夜蛾卵和低龄幼虫有较好的防效[16-18],而对3 龄以上的高龄幼虫及其他虫态的防效不显著[19,20]。雷妍圆等[19]研究发现球孢白僵菌GZSL-1菌株1×108孢子/mL孢子悬浮液对草地贪夜蛾4~6龄幼虫的死亡率仅分别为57.47%、55.06%和25.28%。Visalakshi等[21]在印度从自然感染的草地贪夜蛾幼虫僵虫中分离到1株莱氏绿僵菌,2×108孢子/mL孢子悬浮液处理草地贪夜蛾3龄幼虫一周后的死亡率为90%。本研究分离到的莱氏绿僵菌CDTLJ1菌株对草地贪夜蛾2~6龄幼虫均表现出较好的致病力,对预蛹和蛹也具有一定的致病力,1.2×107孢子/mL处理后第 6 d,2~5龄幼虫的累积校正死亡率均达100%,6龄幼虫的累积校正死亡率为98.33%,预蛹和蛹的累积校正死亡率分别为 28.33%、20.00%。莱氏绿僵菌侵染寄主昆虫的过程与其他虫生真菌的相似,包括附着、萌发、穿透寄主表皮、体内增殖发育并产生毒素,最终导致寄主死亡,菌丝从罹病死亡的虫体内穿透寄主体表产生分生孢子[20,22]。莱氏绿僵菌CDTLJ1菌株对草地贪夜蛾2~6龄幼虫和预蛹、蛹表现出不同的致病力,原因可能是预蛹和蛹的体壁结构或成分影响了病原真菌的侵染和穿透能力[23]。本研究发现被感染的预蛹和蛹体表长出的菌丝和孢子均比罹病幼虫产生的少,原因可能是菌丝难以从体内穿透较厚的体壁和蛹壳,仅能从气门、节间等部位穿出。彭国雄等[24]研究发现,金龟子绿僵菌CQMa421和球孢白僵菌ZJU435处理后第5 d,部分蛹体发黑,菌株ZJU435处理组在保湿培养3~5 d后部分蛹腹节处有少量菌丝出现,与本研究结果类似。终上所述,本研究获得的莱氏绿僵菌CDTLJ1菌株对防控草地贪夜蛾具有良好的应用前景,但仍需要进一步开展田间试验。
莱氏绿僵菌原名莱氏野村菌Nomuraea rileyi[25],直至2014年,Kepler等[26]经过多基因系统分析,并分析绿僵菌属及相关近缘属种的形态学特征及寄主的差异,认为野村菌属Nomuraea应归于绿僵菌属,因此N. rileyi相应改名为M. rileyi。因此,文中构建CDTLJ1菌株的系统进化树中,CDTLJ1菌株与M. rileyi和N. rileyi都在同一进化位置。莱氏绿僵菌的生长及产孢条件较为苛刻,在PDA培养基中生长较为缓慢[16],且产孢量较少[17]。唐维媛等[27]研究表明,增加光照有利于莱氏绿僵菌产孢,杨淑仪等[28]研究表明蚕蛹粉与酵母提取物联合使用对产孢能力有增效作用,且雷妍圆等[17]在 PDA培养基中添加草地贪夜蛾蛹壳可显著提高菌落生长速度和产孢量。本研究获得的莱氏绿僵菌CDTLJ1菌株在SMAY培养基上的生长速度也较慢,培养10 d后的菌落直径也仅为15 mm。因此,要开发利用该菌,还需对其营养特异性和培养条件等开展相关的研究。
猜你喜欢
湖北植保(2022年5期)2022-12-24
中国森林病虫(2021年6期)2021-12-20
中国生物防治学报(2021年1期)2021-05-11
今日农业(2020年13期)2020-12-15
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
中国森林病虫(2018年4期)2018-09-19
西部林业科学(2018年2期)2018-05-16
茶叶学报(2018年4期)2018-02-28
塔里木大学学报(2015年1期)2015-04-25
