
橙色大白菜同核异质细胞质雄性不育系的分子鉴定与园艺性状比较
2021-03-14 05:54王若凡卢倩倩张鲁刚
中国蔬菜 2021年2期
王若凡 马 帅 陈 霖 卢倩倩 孟 艳 张鲁刚
(旱区作物逆境生物学国家重点实验室,西北农林科技大学园艺学院,陕西杨凌 712100)
大白菜〔Brassica campestrisL.ssp.pekinesis(Lour)Olsson〕是我国广泛栽培和食用的十字花科蔬菜之一,具有非常明显的杂种优势。目前,大白菜杂交种制种主要是利用自交不亲和系和雄性不育两用系,但这两种制种方法存在着杂交率低和制种麻烦的弊端。利用胞质雄性不育系制种是作物杂种优势利用最经济有效的途径,因此大白菜细胞质雄性不育系的选育成为生产上迫切需要解决的问题。
大白菜细胞质雄性不育主要分为大白菜波里马甘蓝型油菜细胞质雄性不育(Pol CMS)和萝卜细胞质雄性不育(Ogu CMS)两种类型。细胞质雄性不育性是由细胞核不育基因与细胞质不育基因互作、共同控制的不能产生有活力花粉的母性遗传性状(徐一兰 等,2006)。研究表明,orf138、orf222、orf224分别为Ogu 型萝卜、Nap 型甘蓝型油菜、Pol 型甘蓝型油菜细胞质不育类型的细胞质基因(万正杰 等,2006)。
在大白菜Pol CMS 的相关研究中,柯桂兰等(1992)用大白菜3411-7 为父本转育Pol CMS,最终获得了大白菜胞质雄性不育系CMS3411-7。杨晓云等(2006)以不结球白菜Pol CMS 为不育源,用大白菜自交系作转育父本,通过杂交、多代回交及不育性选择,育成稳定性强、耐低温的大白菜Pol CMS 不育系A7,并且已经用于杂交种配制。但Pol CMS 在偏低温度条件下常产生少量花粉,从而影响杂交种的纯度。Ogu CMS 的研究虽然取得了很大进展,但还存在随着转育代数增加材料出现退化等问题亟需解决,因此该不育系未能广泛用于生产之中(崔冰 等,2018)。此外,张德双等(2002)从法国引进了甘蓝型油菜细胞质雄性不育系材料CMS96,经过多代回交转育,成功地转育出5 份不同类型的大白菜细胞质雄性不育系,这些不育系材料不育性稳定,具有较高的利用价值。
本试验在前人研究基础上,以西北农林科技大学大白菜课题组选育的优良橙色大白菜自交系15S1094 及其同核异质不育系18C5、18C6 为研究对象,对保持系和不育系的植株性状、花器形态等进行观测、比较,鉴定细胞质不育基因及不育系与保持系基因组的相似性,以期加深对这2 个胞质不育系的了解,为研究其雄性不育机理及应用于配制优良杂交种奠定基础。
1 材料与方法
1.1 试验材料
本试验使用的叠抱头球型同核异质橙色大白菜细胞质雄性不育系及其保持系均由西北农林科技大学园艺学院十字花科蔬菜作物遗传改良技术创新实验室培育而成(表1),不育源12XC5 为普通白菜,11-8 为大白菜。试验材料于2018 年2 月和8月种植在西北农林科技大学杨凌蔬菜试验站,每份材料取3 个单株进行试验。
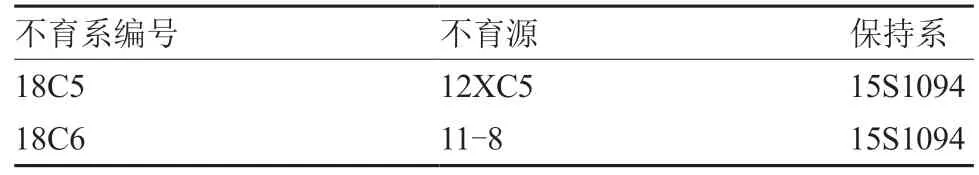
表1 同核异质橙色大白菜细胞质雄性不育系及其保持系
1.2 不育系不育细胞质基因鉴定
1.2.1 试验材料种植及DNA 提取 分别取各试验材料种子20 粒进行浸种催芽,待种子露白时播种育苗;当幼苗长至4~5 片真叶时移栽到大田;缓苗后,分别选3~5 株生长整齐一致的单株,取叶样。采样时,先将2 mL 的离心管灭菌,然后用镊子取0.2 g 左右的叶片放入离心管中,置于-80 ℃冰箱中冷冻保存。采用李荣华等(2009)提出的改进CTAB 法提取基因组DNA,提取时用磨样器(Retsch MM400 球磨仪)磨样。采用1%琼脂糖凝胶电泳检测DNA 的完整性,采用NanoDrop 2000c 微量紫外分光光度计检测DNA 的质量和浓度,统一稀释到100 ng·μL-1作为PCR 扩增模板。
1.2.2 引物合成 选用已知的3 对细胞质雄性不育基因扩增引物(表2)(王国芳 等,2014)进行鉴定,所有引物均由生工生物工程(上海)股份有限公司合成。
1.2.3 反应体系及扩增程序 PCR 扩增反应程序参照王国芳等(2014)的方法。扩增体系总体积为10 μL,其中包括2 ×TaqMaster Mix for PAGE(Vazyme)5 μL,模板DNA 1 μL,正向引物0.5 μL,反向引物0.5 μL,ddH2O 3 μL。

表2 鉴定细胞质雄性不育基因的特异引物
1.3 不育系与保持系基因组DNA 一致性鉴定
1.3.1 DNA 的提取及检测 提取及检测方法同1.2.1。
1.3.2 引物合成 选用已知的28 对大白菜核心SSR 引物(隋光磊 等,2014)对橙色大白菜不育系及其保持系材料进行基因组一致性分析。所有引物均由生工生物工程(上海)股份有限公司合成。
1.3.3 反应体系及扩增程序 扩增反应程序参照隋光磊等(2014)的方法,扩增体系同1.2.3。
1.3.4 电泳检测 扩增产物用浓度为12%的非变性聚丙烯酰胺凝胶电泳分离。电泳参数设置:电压200 V,电流100 mA,时间2 h。电泳结束后,用0.1%硝酸银染色5 min,1.5%氢氧化钠和1%甲醛进行显影,并拍照记录。选择清晰易辨的多态性带型进行统计,在相同迁移率位置,有条带记为“1”,无条带记为“0”,缺失记为“—”。利用Excel 软件计算不育系与保持系的遗传相似系数(即不育系和保持系共有扩增条带数占总扩增条带数的比值),分析其遗传相似率。
1.4 不育系与保持系花器比较观察
2018 年冬季种植不育系及其保持系,2019 年春季开花,在盛花期晴天上午10:00 左右观察不育系及其各自保持系的主花序花器官形态并测量。每种材料随机取5 株,每株观察6 朵花,利用游标卡尺测量四强雄蕊长、两弱雄蕊长、四强花丝长、花瓣长、花瓣宽、萼片长、雌蕊长、花柱长,观察蜜腺大小、花药形状、花瓣形状并拍照记录,使用IBM SPSS Statistics 23 软件对试验数据进行差异显著性分析。
1.5 不育系与保持系植物学性状观察
2018—2019 年秋季种植不育系及其保持系,在结球初期和成熟期分别对不育系和保持系的相关性状进行调查,莲座期观测叶色、叶缘锯齿、叶缘波状、绒毛密度、泡状凸起数量、泡状大小、叶色光泽度、叶脉等8 个性状,成熟期测定株高、开展度、单株质量、单球质量、球高、球宽、外叶数等7 个性状。从植物学性状上统计比较不育系和保持系的一致性,具体标准参考《植物品种特异性、一致性和稳定性测试指南—大白菜》(GB/T 19557.5—2004)。
株高为地上植株基部至植株最顶处的高度(cm),开展度为植株外叶开展纵、横两个垂直方向最大距离(cm),单株质量为去除根部后的大白菜整株质量(kg),单球质量为去除外叶后的大白菜质量(kg),球高为大白菜叶球底部至顶部的高度(cm),球宽为大白菜球肩最宽部位的宽度(cm),外叶数为成熟期叶球外绿色叶片数。
2 结果与分析
2.1 橙色大白菜细胞质雄性不育系的不育细胞质基因鉴定
利用3 对已知的细胞质雄性不育基因特异引物,对参试不育系和保持系材料进行PCR 扩增,其中orf224的引物在3 份材料中均未扩增出产物;orf138的引物从18C5 和18C6 中均扩增出长度约300 bp 的片段,测序结果与orf138序列一致性为99%;orf222的引物仅在18C6 中扩增出700 bp 的条带,测序结果与orf222的序列高度同源、一致性为99%(图1)。说明18C5 仅含有orf138基因,应该属于Ogu 细胞质雄性不育;18C6 同时含有orf138和orf222基因,简称Ogu-Nap CMS;3 对引物在保持系中均没有扩增出产物,即保持系不含不育基因。
2.2 橙色大白菜细胞质雄性不育系与保持系细胞核DNA 相似性的分子鉴定
用已知的28 对大白菜核心SSR 引物对参试不育系和保持系的各3 个单株进行PCR 扩增,结果表明18C5 与保持系15S1094 的平均遗传相似系数为91.11%,18C6 与保持系15S1094 的平均遗传相似系数为93.76%(表3)。由此可知,18C6 与保持系15S1094 的遗传相似性更相近。

表3 橙色大白菜细胞质雄性不育系与保持系的遗传相似性分析
2.3 橙色大白菜细胞质雄性不育系与保持系花器比较观察
2 个同核异质不育系在春季开花早期低温条件下均有败蕾现象,但随着温度升高开花趋于正常。对花器观测发现,不育系与保持系花色均为黄色;保持系开放花的柱头略低于雄蕊,花丝较长,花药呈近长方体,形态饱满、有大量淡黄色花粉,雄蕊、蜜腺均正常;不育系18C5 的柱头明显高于雄蕊,雄蕊花丝短,花药呈长锥体、干瘪扁平,甚至退化成丝状,无花粉;不育系18C6 的柱头略高于雄蕊,雄蕊花丝较短,但较18C5 长,花药呈锥体、无花粉。2 个不育系的蜜腺数量均正常,虽然18C5 的蜜腺较小(图2),但均可以吸引蜜蜂、蝴蝶,正常授粉结籽。

表4 橙色大白菜细胞质雄性不育系及其保持系的花器官性状指标
从表4 可以看出,2 个不育系的花瓣长、花瓣宽、四强雄蕊长、四强花丝长均小于保持系,且18C5 小于18C6,达到显著或者极显著差异水平;2 个不育系的两弱雄蕊长均极显著小于保持系,但2 个不育系之间差异不显著;2 个不育系的花柱长和雌蕊长与保持系差异不显著;2 个不育系的萼片长变化比较特殊,18C5 的萼片长大于保持系,而18C6 的萼片长小于保持系。这说明从不同不育源转育来的2 个同核异质不育系花器官的雌蕊没有受到不育性的影响,而其他器官都不同程度变小,其中18C5 变小较18C6 更严重。
2.4 橙色大白菜细胞质雄性不育系与保持系植物学性状比较
从图3 和表5、6 可以看出,莲座期,18C5 的叶色、叶缘锯齿、绒毛密度、泡状凸起数量、泡状大小、叶色光泽度等6 个性状都与保持系一致,18C6 的叶色、叶缘波状、泡状大小、叶色光泽度等4 个性状与保持系一致,说明莲座期18C5 与15S1094 更为相似;成熟期。18C6 的株高、开展度、球高、球宽、外叶数均与保持系差异不显著,但单株质量与保持系的差异达到极显著水平,单球质量与保持系的差异达到显著水平;18C5 的开展度、球高、球宽、外叶数均与保持系差异不显著,但株高、单株质量与保持系的差异达到极显著水平,单球质量与保持系的差异达到显著水平,说明成熟期18C6 与保持系15S1094 更为相似。

表5 橙色大白菜细胞质雄性不育系与保持系的莲座期性状
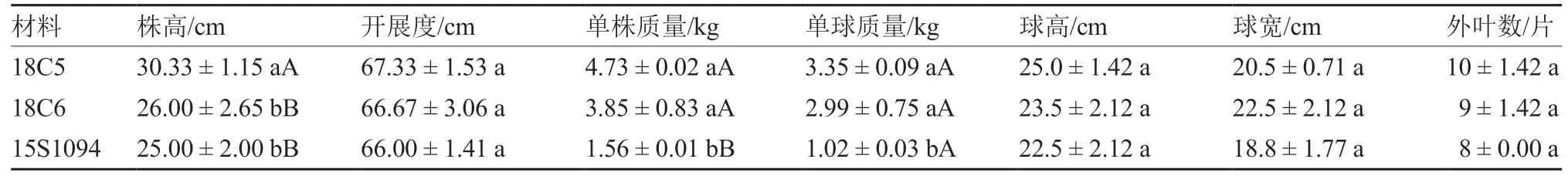
表6 橙色大白菜细胞质雄性不育系与保持系的成熟期性状
3 讨论与结论
橙色大白菜富含类胡萝卜素,对人类健康有益,因此培育橙色大白菜细胞质雄性不育系对开展橙色大白菜杂种优势利用有重要意义。本试验对叠抱头球型橙色大白菜的同核异质细胞质雄性不育系的不育基因、雄蕊特征、植物学性状进行了系统观察,分析了2 个不育系的不育基因、雄蕊退化特征和植物学性状的异同,加深了对这2 个细胞质雄性不育系的了解。
细胞质不育基因分子鉴定结果表明,18C5和18C6 携带不同的细胞质不育基因,18C5 携带orf138不育基因,18C6 携带orf138和orf222不育基因,具有细胞质多样性特点。用大白菜基因组已知的28 对核心SSR 引物对18C5、18C6 及其保持系进行遗传多样性分析,18C5 和18C6 与保持系的平均遗传相似系数分别为91.11%和93.76%,两者差异不大,但均低于理论相似性96.875%,这可能是由于回交早期没有进行植物学性状选择造成的。
对植株生育期性状的观测结果表明,2 个同核异质不育系均花丝短缩、花药干瘪无花粉,不育性100%,雌蕊、蜜腺发育正常,能正常结籽,可以用于杂交制种。值得注意的是:莲座期18C5 与保持系15S1094 的相似度更高;而在成熟期18C6 与保持系15S1094 的相似度更高,即2 个不育系不同发育时期与保持系相似度不同,表明Ogu 型萝卜细胞质雄性不育和Nap 型甘蓝型油菜细胞质雄性不育对植物学性状有不同的影响,同时发现2 个不育系的植物学性状普遍较保持系有明显优势,这可能是不育系核质杂种优势的表现,也可能是不育系的回交代数低或纯合程度不够所显示的异核结合的杂交优势还在起作用。该杂种优势是否能遗传到杂交种中尚不清楚;另外,Ogu 型萝卜细胞质雄性不育和Nap 型甘蓝型油菜细胞质雄性不育对杂种优势是否有影响,也有待于进一步研究。
猜你喜欢
广西植物(2022年1期)2022-03-17
中国农学通报(2022年4期)2022-03-02
课外生活·趣知识(2022年2期)2022-02-08
家庭教育报·教师论坛(2021年42期)2021-12-23
河北画报(2021年2期)2021-05-25
北方论丛(2021年2期)2021-05-22
高考·中(2019年6期)2019-09-10
分析化学(2019年3期)2019-03-30
现代农业科技(2018年21期)2018-01-12
科学中国人(2016年30期)2016-07-14
