
高表达FcεR1α和共表达NPY-EGFP的RBL细胞系构建及生物学功能检测
2021-04-01 01:12黄伦辉张嘉懿李柳栩张蓓刘运德李会强
现代免疫学 2021年2期
黄伦辉,张嘉懿,李柳栩,张蓓,刘运德,李会强
[1.天津医科大学 医学检验学院,天津 300203;2.中国医学科学院血液病医院(中国医学科学院血液学研究所) 实验血液学国家重点实验室, 国家血液系统疾病临床医学研究中心, 天津 300020;3.天津市儿童医院 检验科,天津 300074]
特异性IgE(specific IgE, sIgE)抗体在变态反应性疾病中处于核心地位,其生物功能是通过结合四聚体高亲和力IgE受体而介导的。目前,sIgE的检测是含量测定而非功能测定[1],与临床症状相关性不高,因为sIgE水平仅仅表示IgE结合过敏原的强度,并不能反映结合过敏原引起的致敏细胞脱颗粒效应。
高亲和力IgE受体α链(high affinity IgE receptor alpha chain,FcεR1α)在RBL细胞上的表达可用于检测sIgE与过敏原结合所导致的组胺、5-羟色胺等炎性介质的胞吐[2]。目前,RBL报告系统有RS-ATL8和NFAT-DsRed,用于研究纯化的花生过敏原诱导脱颗粒的能力或检测食物过敏原[3-4]。这2种系统可实现高通量测量,后者也可以与过敏原阵列配合使用[5],但均不能直接反映脱颗粒效应[6]。
Lang等[7]的研究发现,神经肽Y(neuropeptide Y,NPY)与荧光蛋白融合表达可用于标记神经元细胞系PC12中的颗粒。Azouz等[8]的研究证明,NPY-mRFP也可以用来标记非神经元RBL细胞中的颗粒,用于定量细胞胞吐。本研究假设稳定转染NPY-增强型绿色荧光蛋白(enhanced green fluorescent protein, EGFP)和FcεR1α的RBL细胞可以用于人类血清中致敏作用的检测,通过定量荧光释放率来评估致敏效应。
1 材料与方法

1.2 方法
1.2.1 基因表达载体的构建 通过NCBI网址查询获得FcεR1α、NPY-EGFP的基因序列,由义翘神州生物公司合成并构建质粒pLV-Puro-FcεR1α-T2A-NPY-EGFP-TYK2,对其进行测序鉴定后无内毒素大量提取。
1.2.2 脂质体Lipo2000转染 取5×105个/孔指数增长期RBL-2H3细胞过夜培养,第2天换5%血清培养液。对照质粒和目的质粒各3 μg,分别加入250 μL无血清培养液,取Lipo2000 20 μL加入500 μL无血清培养液,室温静置5 min后分别混合。10 min 后逐滴加至细胞,混匀,37 ℃培养6 h,换完全培养液再继续培养48 h。
1.2.4 FcεR1α亚基蛋白表达水平的检测 分别收集一孔转染前和转染后的RBL-2H3细胞,用SDS裂解液对细胞进行裂解。采用湿转的方法将等量的蛋白质转移到聚偏二氟乙烯(polyvinylidene fluoride, PVDF)膜上。在室温下用5%脱脂奶粉封闭2 h后,于4 ℃过夜孵育鼠抗人FcεR1α抗体(1∶2 000),然后用0.05% Tween-20 TBST洗膜3次(10 min/次),继续室温孵育二抗羊抗鼠IgE抗体1 h,在1∶5 000的比例下室温持续孵育1 h。使用ChemiDoc XRS+成像系统检测信号。
计数转染前后细胞各1×105个,以300×g离心10 min。用100 μL FACS缓冲液[2 mmol/L EDTA,0.5%牛血清白蛋白(bovine serum albumin, BSA),500 mL PBS] 重悬细胞,加入1 μL PE标记的鼠抗人FcεR1α抗体,混合后避光置于4 ℃冰箱30 min。然后清洗去除未结合的抗体。最后使用BD FACSVerse FCM进行检测。
1.2.5 β-氨基己糖苷酶介质释放试验 用1∶10稀释血清对转染后RBL-2H3细胞于37 ℃孵育1 h进行致敏。过夜培养后,用缓冲液A(140 mmol/L NaCl、5 mmol/L KCl、0.6 mmol/L MgCl2、1.0 mmol/L CaCl2、5.5 mmol/L葡萄糖、0.1%BSA和10 mmol/L哌嗪-1, 4-二乙磺酸,pH7.4)冲洗细胞2次。在预热的缓冲液A中将稀释的BLG(1 μg/mL)或羊抗人IgE抗体添加到细胞中(50 μL/孔),37 ℃孵育30 min。然后收集上清液,用0.2% Triton X-100裂解细胞。以4-甲基伞形酰-N-乙酰基-β-D-葡糖胺(溶于100 mmol/L柠檬酸,pH4.5)为底物,在37 ℃孵育30 min后用0.25 mol/L甘氨酸缓冲液终止反应。在Biotek Synergy2上读取光密度[D(405 nm)]值,计算β-氨基己糖苷酶释放率。β-氨基己糖苷酶释放率=(S-B)/(T-B)×100%,其中B、S、T分别代表培养液、细胞上清液培养液和剩余细胞裂解后培养液的D值。
1.2.6 NPY-EGFP释放试验 用Tyrode缓冲液洗涤转染细胞3次。在96孔板,用稀释血清过夜致敏,用1 μg/mL BLG在37 °C下孵育30 min。将每孔上清液小心地移到新的96孔板,放在冰上,避光。使用荧光板阅读器在488 nm激发光和520 nm发射光下测量荧光信号值。NPY-EGFP释放率=(S-B)/(T-B)×100%,其中B、S、T分别代表培养液、细胞上清液培养液和剩余细胞裂解后培养液的荧光信号值。
2 结果


注:CMV,巨细胞病毒(cytomegalovirus);RRE,Rev反应元件(Rev response element);cPPT/CTS,中央聚嘌呤序列(central polypurine tract/central);mPGK,鼠磷酸甘油酸激酶(mouse phosphoglycerate kinase);Puro,嘌呤霉素(puromycin);WPRE,土拨鼠肝炎病毒转录后调控元件(woodchuck hepatitis post-transcriptional regulatory element);Amp,氨苄西林(ampicillin)。图1 表达载体pLV-Puro-FcεR1α-T2A-NPY-EGFP-TYK2结构示意
2.2 转染细胞中FcεR1αmRNA的表达用实时荧光定量PCR检测转染组和空载组FcεR1αmRNA的相对表达水平,用2-ΔΔCq方法分析数据,以转染空载质粒表达量作为“1”,转染组FcεR1α的表达量是空载组基因表达量的165.905 2倍(图2)。这提示目的基因高表达于RBL-2H3细胞上。

图2 实时荧光定量PCR 检测FcεR1α基因表达水平
2.3 受体蛋白的表达使用抗人 FcεR1α 特异性单克隆抗体检测空载组和目的组细胞中 FcεR1α的表达。目的组在相对分子质量为50 000 处检测到一条明显的蛋白条带,而空载组未能检测到该条带(图3)。
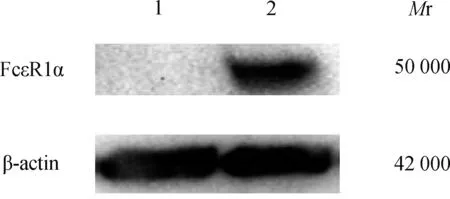
注:1为空载组; 2为目的组。图3 Western blotting 检测 FcεR1α 的蛋白表达
使用PE标记的抗人FcεR1α特异性单克隆抗体识别空载组和目的组细胞中FcεR1α的表达情况,目的组阳性细胞率达83.2%。这提示绝大多数受体表达在细胞膜上。(图4)
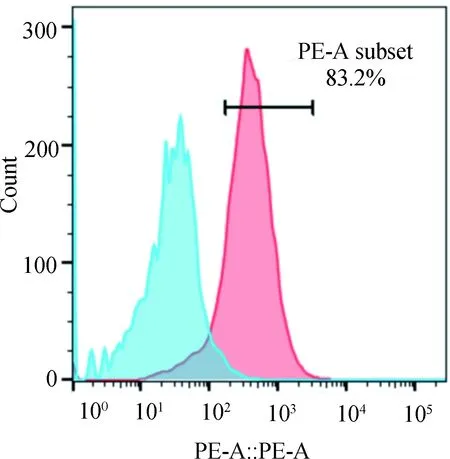
图4 FACS检测FcεR1α的细胞定位
2.4 受体蛋白活性试验使用24例过敏阳性血清和1例过敏阴性血清进行β-氨基己糖苷酶介质释放试验。过敏阳性血清β-氨基己糖苷酶释放率为20%~60%,过敏阴性血清在10%以下(图5)。β-氨基己糖苷酶介质释放试验证明了人血清sIgE导致了脱颗粒反应,FcεR1α具有结合血清sIgE的能力。
用相同血清样本,更换新型指标NPY-EGFP检测释放率。过敏阳性血清NPY-EGFP释放率为20%~80%,与过敏阴性血清可明显区分(图6),这为接下来的诊断指标研究奠定了物质基础。

注:样本1~24为过敏阳性血清; 样本25为过敏阴性血清。图5 转染后的RBL-2H3细胞β-氨基己糖苷酶介质释放试验

注:样本1~24为过敏阳性血清; 样本25为过敏阴性血清。图6 转染后的RBL-2H3细胞NPY-EGFP释放试验
3 讨论
检测血清sIgE的方法由物理化学法向细胞生物学法过渡是历史性的进步。细胞生物学法检测sIgE不仅能反映过敏原结合sIgE的能力,而且能同步反映过敏原的致敏情况,可进一步研究机体的过敏体征。RBL-2H3细胞系的发展基于一种FcεR1α的嵌合受体复合物,其中至少需要转染人源的α链,与大鼠肥大细胞内源性α链竞争结合大鼠内源性β、γ链[10]。经典RBL-2H3细胞系的一个缺点是某些人血清对大鼠细胞具有细胞毒性,需要将血清高度稀释方可减少此毒性,但这样做会导致产生不充分的致敏作用,造成假阴性结果。所以,新一代报告细胞系正在利用敏感报告基因(如荧光素酶或荧光蛋白)替代以前的人源化RBL细胞系,这样即使使用高达1∶100稀释度的血清,也可以检测过敏原sIgE。目前,国外使用2种这样的人源化RBL报告系统,分别是RS-ATL8和NFAT-DsRed。这2种系统可实现高通量测量,后者也可以与过敏原阵列配合使用。RS-ATL8非常敏感,但有2个主要的缺点:首先,其需要荧光素酶底物,常规使用相对昂贵;其次,RS-ATL8并不适合用于评估过敏原阵列上的活化,因为荧光素酶检测需要裂解细胞,这会使荧光信号与过敏原结合位点的位置出现错位[4,11]。为了克服这些缺点,一种类似于RS-ATL8的荧光报告系统NFAT-DsRed被开发出来,通过诱导红色荧光蛋白表达来报告IgE依赖性活化T细胞核因子核易位[5]。不过,这些报告系统均不能直接反映脱颗粒效应。本研究所构建的工具细胞自带EGFP,省去添加酶底物的繁琐步骤和额外花费,同时作为预先合成的新型标志物NPY直接反映了过敏反应中的脱颗粒效应,更好地体现了过敏原与高亲和受体表面sIgE的结合,有望实现高通量和高灵敏度检测。
本研究为开发一种稳定高表达FcεR1α和共表达且便宜的新型指标NPY-EGFP的细胞系奠定了坚实的基础。从基因水平、蛋白水平、膜蛋白表达水平及功能试验方面证明了该受体表达的可实现性和优越性,以及重要的新型指标NPY-EGFP开发工作的实现。FcεR1α从基因水平检测,其表达量特别高,蛋白表达量相对高,但NPY-EGFP荧光强弱不一导致检测的释放率不均质。事实上,正是由于瞬时表达基因定位的不确定性,才体现出建立稳定表达细胞系的重要性。通过后期筛选细胞有望得到高表达FcεR1α且具有均质荧光的NPY-EGFP的细胞系。
表达NPY-EGFP的人RBL细胞系的新型报告系统可用于分析与食物蛋白质生物学相关的交叉反应性,鉴定过敏原的生物学特性,诊断食物过敏[12],研究食物蛋白质的致敏潜力,比较食品加工前后的变应原性。在对受试者的过敏症状进行诊断时,须使用纯化的过敏原或过敏原提取物,而基于RBL细胞的免疫测定可提供关于全食物产品引起过敏反应的能力信息[13],筛选潜在的抗过敏食品化合物或功能表位,然后用于针对食物过敏的免疫治疗。
猜你喜欢
中成药(2018年5期)2018-06-06
中成药(2017年8期)2017-11-22
食品与机械(2017年5期)2017-07-05
邯郸职业技术学院学报(2016年2期)2016-02-27
山东医药(2015年14期)2016-01-12
中国医疗美容(2015年1期)2015-07-12
江苏大学学报(医学版)(2015年2期)2015-04-17
中国医药导报(2015年26期)2015-02-28
天然产物研究与开发(2014年3期)2014-04-27
茶叶通讯(2014年4期)2014-02-27
