
西农萨能奶山羊瘤胃产蛋白酶酵母菌的筛选研究
2021-04-14 01:01魏满红张佳运陈玉林杨雨鑫
饲料工业 2021年5期
■魏满红 张佳运 陈玉林 杨雨鑫
(西北农林科技大学动物科技学院,陕西杨凌712100)
幼龄动物的消化器官未发育完全,其所分泌的蛋白酶不能将饲料中的蛋白质完全消化,将蛋白酶添加在饲料中可以促进幼龄动物对蛋白质消化吸收,同时蛋白酶也可以促进饲料中抗营养因子的降解,提高动物机体的免疫力。与瘤胃微生物相比,添加外源菌可能会与瘤胃微生物产生拮抗等不良反应。从瘤胃中筛选出的产蛋白酶菌更易于在瘤胃中定植[1]。在饲料行业内根据“最适作用pH 值”为标准,将蛋白酶分为酸性蛋白酶、中性蛋白酶以及碱性蛋白酶[2]。碱性蛋白酶的最适作用pH 值为9~11,最先在猪的胰腺中发现,主要在小肠中发挥作用,碱性环境下可以把蛋白质水解成小分子肽链[3];从酶的活力pH值曲线分析可知,酸性蛋白酶的最适作用pH 值为2~4,能够水解蛋白质肽键,酸性蛋白酶的活性部位中含有一个或更多的羧基;中性蛋白酶是一种可用于蛋白质水解的内切酶,目前也已在饲料行业中广泛应用[4]。利用微生物生产蛋白酶具有成本低、产出率高、周期短、易控制等优点,为当下规模化生产蛋白酶主要来源[5]。产蛋白酶微生物主要有乳酸杆菌和芽孢杆菌,但产酶菌种较为单一,难以满足工业化对各类蛋白酶的生产需求。有研究表明,酵母菌具有高生产率、发酵快[6]等优势,但是关于酵母菌产蛋白酶报道较少。本试验对山羊瘤胃内容物中产蛋白酶酵母菌进行初步分离和鉴定,并对其酶学特性进行初步研究。
1 材料与方法
1.1 样品的来源、培养基、试剂
1.1.1 样品的来源
从西北农林科技大学畜牧站内抽取西农萨能奶山羊瘤胃液,立即装入无菌管,带回实验室保存在-80 ℃冰箱中。
1.1.2 培养基
蛋白酶培养基:葡萄糖50 g、蛋白胨10 g、KH2PO42 g、MgSO4·7H2O 0.4 g、用无菌水定容至1 000 mL,pH值调至7,121 ℃灭菌15 min。
植酸酶培养基:氯化钾0.5 g、植酸钠2 g、硫酸锰0.03 g、葡萄糖30 g、硫酸镁0.5 g、硫酸亚铁0.03 g、硝酸铵5 g,用无菌水定容至1 000 mL,pH 值调至5.5,121 ℃灭菌15 min。
纤维酶培养基:羧甲基纤维素钠10 g、蛋白胨2 g、酵母膏0.5 g、K2HPO41.5 g、Na2SO42.5 g、琼脂20 g,用无菌水定容至1 000 mL,pH 值调至7.0,121 ℃灭菌15 min。
果胶酶培养基:果胶10 g、酵母提取物10 g、蛋白胨20 g、葡萄糖20 g、琼脂20 g,用无菌水定容至1 000 mL,pH值调至7.0,121 ℃灭菌15 min。
YPD 培养基:葡萄糖20 g、酵母粉5 g、蛋白胨10 g,用无菌水定容至1 000 mL,121 ℃灭菌15 min。
1.1.3 试剂
羧甲基纤维素钠(CMC-Na)、3,5-二硝基水杨酸、葡萄糖、植酸钠、果胶、牛肉膏、柠檬酸三铵、乙酸钠、蔗糖、琼脂、酪蛋白、干酪素、酪氨酸、三氯乙酸、脱脂乳奶粉、HCl、PDA、KH2PO4、KCl、MgSO4、FeSO4、MnSO4、吐温80、NaCl、NaOH、H2SO4、Na2CO3、NaH2PO4·2H2O、Na2HPO4·12H2O,购自杨凌晓白化玻仪器公司;酵母提取物、蛋白胨,购自OXOID 公司;Folin-酚,购自北京东方顺科生物科技有限公司;2×Taq PCR Master⁃Mix,TIANampYeast DNA kit(离心柱型),gelRed 核染色剂,引物NL-1(5-GCATATCAATAA GCGGA GGA AAAG-3),NL-4(5-GGTCCGT GTTTCAAGACG-3)购自北京擎科生物技术有限公司。
1.2 试验方法
1.2.1 产蛋白酶酵母菌株的筛选
将瘤胃液10 倍梯度稀释于灭菌水中,在摇床上振荡培养30 min。取20 μL 稀释液涂布于PDA 固体培养基,37 ℃恒温培养24 h,挑取符合酵母菌菌落形态的菌株进行纯化。将纯化得到的酵母菌株,接种在脱脂乳培养基中进行初筛,置于30 ℃恒温培养箱中培养5 d[7],筛选出水解圈较大的菌株进行活化扩繁,将菌株培养液以1∶1 与30%甘油混合保存于2 mL EP管中,放置-80 ℃冰箱内进行保存。
1.2.2 菌株的分子生物学鉴定
将2 mL左右的种子液加入离心管中,10 000 r/min离心2 min,弃上清液富集菌体,使用酵母菌基因组DNA 提取试剂盒提取目标菌株基因组DNA。以基因组DNA 为模板,用酵母菌通用引物NL-1 和NL-4 对26S rDNA D1/D2 区基因片段进行PCR 扩增。扩增条件:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火1 min,72 ℃延伸45 s,共35 个循环;最后72 ℃延伸10 min。1%琼脂糖凝胶电泳确认扩增效果和片段大小。测序结果应用NCBI Blast 在线比对工具对测序结果进行同源性比对。
1.3 产蛋白酶酵母菌产酶条件优化
将筛选出来的菌株从-80 ℃冰箱拿出,立即放置到冰盒中进行冰浴融化,再接种到液体发酵培养基中,于不同温度、不同pH值、不同接种量浓度、100 r/min条件下摇床培养,采用福林法[8]测定粗酶液中蛋白酶活力。
1.3.1 培养时间对菌株产蛋白酶影响
将初始菌液按体积分数3%接种量,在30 ℃、pH为7、100 r/min 条件下摇床培养,每隔12 h 取样,5 000 r/min条件下离心10 min,取上清液测蛋白酶活力,绘制酶活统计图,确定最适产酶时间。
1.3.2 接种量对菌株产蛋白酶的影响
将初始菌液按体积分数1%、2%、3%、4%、5%接种量在30 ℃、pH 值为7、100 r/min 条件下摇床培养,培养时间为1.3.1 得到的各菌株最佳产酶时间,5 000 r/min 离心10 min,取上清液测蛋白酶活力,绘制酶活统计图,确定最适接种量。
1.3.3 培养温度对菌株产蛋白酶的影响
将初始菌液按1.3.2得到的各菌株最佳接种量,在温度分别为18、25、30、37、50 ℃、pH 值为7、100 r/min条件下摇床培养,培养时间为1.3.1 得到的各菌株最佳产酶时间,5 000 r/min 离心10 min,取上清液测蛋白酶活力,绘制酶活统计图,确定最适培养温度。
1.3.4 初始培养基pH值对菌株产蛋白酶的影响
将培养基的初始pH 值分别调节为5、6、7、8、9、10,各菌株按1.3.2 最佳接种量及1.3.3 最佳温度、100 r/min 条件下摇床培养,培养时间为1.3.1 各菌株最佳产酶时间,5 000 r/min 离心10 min,取上清液测蛋白酶活力,绘制酶活统计图,确定最适产酶pH值。
1.4 产植酸酶、果胶酶、纤维素酶三种水解酶效果的研究
将初始菌液按体积分数3%接种量接入产纤维素酶培养基、产果胶酶培养基、产植酸酶培养基,30 ℃培养48 h 后,5 000 r/min 离心10 min,分别取上清液采用DNS 法测定纤维素酶活力、果胶酶活力、植酸酶活力,绘制酶活统计图。
2 结果
2.1 产蛋白酶酵母菌株的筛选结果
通过比对《酵母菌的特征与鉴定手册》与平板上的菌株外观形态,结合相关试验分析进行初筛,筛选出Y1、Y5、Y12、Y16、Y19、Y20、Y21、Y22、Y27、Y28、Y29、Y30等12株酵母菌。将这12株菌点涂到脱脂乳培养基上30 ℃恒温培养5 d,Y1、Y5、Y16、Y20、Y21五株菌均出现明显的水解圈(见图1)。

图1 具有明显水解圈菌落
2.2 分子生物学鉴定
将分离出的五株酵母菌基因组进行26S rDNA区PCR 扩增,用1%的琼脂糖凝胶对扩增结果检测,电泳顺序为2 000 bp Marker、Y1、Y5、Y16、Y20、Y21,扩增后的结果如图2所示。

图2 菌株26S rDNA PCR扩增产物电泳图
5 株菌扩增出的片段长度均在500~750 bp,条带清晰可见,无杂带,可用于菌株26S rDNA 测序。将PCR 产物送北京擎科生物科技有限公司武汉分公司测序,利用QuickStart 软件对测序结果进行拼接后通过NCBI Blast 在线比对工具对测序结果进行同源性比对。Y1为库德毕赤酵母、Y5为东方伊萨酵母、Y16为马克斯克鲁维酵母、Y20为近平滑假丝酵母、Y21为单孢酿酒酵母。
2.3 产蛋白酶酵母菌产酶条件优化结果
2.3.1 培养时间对菌株产蛋白酶的影响
3%接种量在培养基pH 值为7、30 ℃、100 r/min条件下摇床培养,培养时间对菌种产蛋白酶的影响如图3所示。随着时间的增加,培养液中菌株产蛋白酶活力也逐渐增加,在48 h 时产蛋白酶活力达到了最高;培养超过48 h 后,蛋白酶活力逐渐减少。因此筛选出的产蛋白酶酵母菌株的最适产酶时间为48 h。
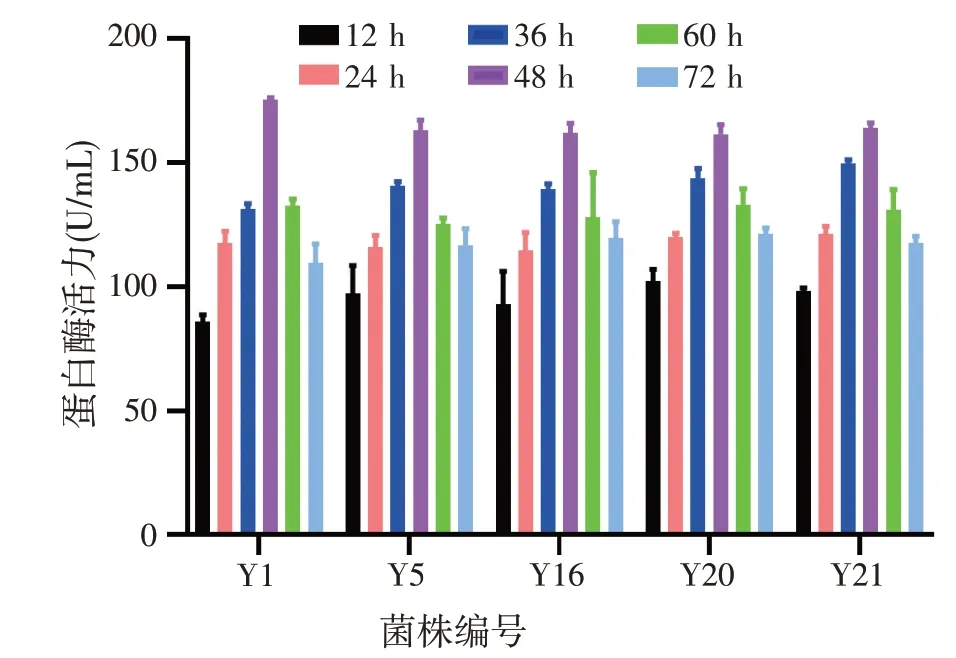
图3 培养时间对菌株产蛋白酶的影响
2.3.2 菌液接种量对菌株产蛋白酶的影响
将初始菌液按体积分数1%、2%、3%、4%、5%的接种量,在培养基pH 值为7、30 ℃、100 r/min 条件下摇床培养48 h,测定蛋白酶活力,不同接种量对菌株产蛋白酶的影响如图4 所示。在接种量1%时,产蛋白酶活力较低;随着初始菌液接种量增加,产蛋白酶活力逐渐上升,其中菌株库德毕赤酵母Y1、马克斯克鲁维酵母Y16在3%时酶活最大,东方伊萨酵母Y5、近平滑假丝酵母Y20、单孢酿酒酵母Y21在4%时酶活最大。

图4 接种量对菌株产蛋白酶的影响
2.3.3 培养温度对菌株产蛋白酶的影响
各菌株按2.3.2得到的最佳接种量、培养基pH为7、100 r/min条件下摇床培养48 h,不同培养温度对菌株产蛋白酶的影响如图5 所示。菌株库德毕赤酵母Y1、单孢酿酒酵母Y21产蛋白酶的最佳温度为30 ℃,菌株东方伊萨酵母Y5、马克斯克鲁维酵母Y16、近平滑假丝酵母Y20产蛋白酶的最佳温度为37 ℃。
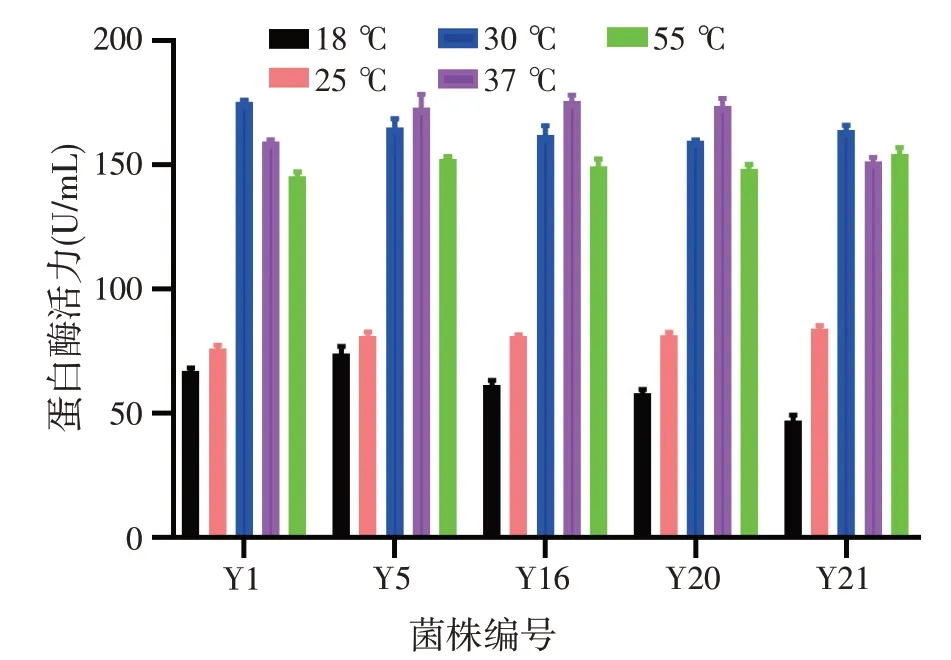
图5 培养温度对菌株产蛋白酶的影响
2.3.4 初始培养基pH对菌株产蛋白酶的影响
各菌株按2.3.2 得到的最佳接种量,2.3.3 得到的最培养温度、100 r/min、不同培养基pH 值(5、6、7、8、9、10)条件下摇床培养48 h,不同培养基pH值对菌种产蛋白酶的影响如图6 所示。随着pH 值的升高,各培养液中蛋白酶的活性也逐渐升高,在pH值为9时5株菌的培养液中蛋白酶的活性均最高。

图6 初始培养基pH对菌株产蛋白酶的影响
2.4 产植酸酶、果胶酶、纤维素酶三种水解酶效果的研究
2.4.1 产植酸酶效果的研究
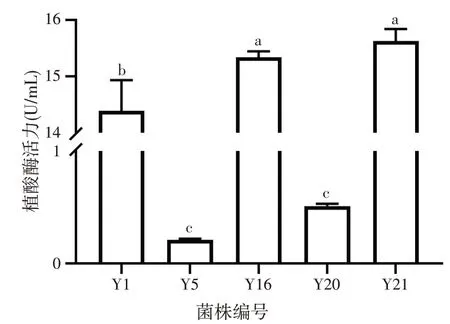
图7 产植酸酶效果的研究
由图7可知,各菌株以3%的添加量,100 r/min条件下摇床培养48 h后,库德毕赤酵母Y1、马克斯克鲁维酵母Y16、单孢酿酒酵母Y21具有较高的植酸酶活性,酶活力分别达到14.38 U/mL、15.34 U/mL、15.63 U/mL。东方伊萨酵母Y5和近平滑假丝酵母Y20产生的植酸酶较少,低于1.00 U/mL。
2.4.2 产果胶酶效果的研究
由图8可知,各菌株以3%的添加量,100 r/min条件下摇床培养48 h后,五株菌株均表现出较高的果胶酶活性。其中单孢酿酒酵母Y21 酶活力显著高于其他菌株(P<0.05),达到15.18 U/mL。
2.4.3 产纤维素酶效果的研究

图8 产果胶酶效果的研究

图9 产纤维素酶效果的研究
由图9 可知,各菌株以3%的添加量,100 r/min条件下摇床培养48 h 后,五株菌株均表现出较高的纤维素酶活性,均超过180 U/mL,其中近平滑假丝酵母Y20 酶活力显著高于其他菌株(P<0.05),达到199.08 U/mL。
3 讨论
3.1 发酵条件对微生物蛋白酶活力的影响
随着工业用酶用量的不断增加,已有的蛋白酶无法满足日益增长的需求。筛选出新的高产产酶菌株和对已有的菌株产酶条件进行优化以提高蛋白酶的产量和品质,是两种最主要的解决方案[9]。从土壤、污泥、海水及动物肠道中已经分离出可产生中性、碱性或耐盐性蛋白酶的微生物菌株,同时也从不同来源的植物中筛选出了产蛋白酶的内生真菌、细菌和放线菌[10],极大地拓宽了产蛋白酶菌的菌株库。菌株在液体培养基中发酵,其产酶能力也会受到培养基发酵条件的影响。温度、pH 值、发酵的时间等条件会对菌株的生产繁殖产生影响,从而导致最终的蛋白酶活性不同[11]。
本研究通过点涂于脱脂乳平板来初筛具有降解蛋白酶的菌,乔传丽等通过计算HE 值,比对HE 大小初步判断产蛋白酶活力,但是实际上培养出来的菌落并不是完整的圆形,通过测量水解圈直径、菌落直径等数据,误差较大,缺乏科学严谨性。据报道,HE值和蛋白酶活力的高低并非显著的正相关[12]。因此结合相关实际状况,通过肉眼观察,找出水解圈相对而言比较明显的五株菌,进行后续产蛋白酶条件优化的试验。
将筛选出的菌株以3%接种量在培养基pH 为7、温度30 ℃、100 r/min条件下培养,12 h时菌株还未开始增殖,数量较少,产蛋白酶能力有限,故产蛋白酶活力较低;随着时间的增加,酵母菌数量增多,培养液中菌株产蛋白酶活力也逐渐增加,在48 h时产蛋白酶活力达到了最高;这与Zhou等[13]的研究结果一致。但随着时间的延长,培养液中营养物质减少蛋白酶活力逐渐降低。刘静等[14]发现产蛋白酶菌在30 h 蛋白酶活力达到最高。在接种量1%时,培养48 h 产蛋白酶活力较低,随着初始菌液接种量增加,产蛋白酶活力逐渐上升,其中菌株库德毕赤酵母Y1、东方伊萨酵母Y5、马克斯克鲁维酵母Y16在3%时酶活力最大,近平滑假丝酵母Y20、单孢酿酒酵母Y21在4%时酶活力最大。这同曹慧等[15]的研究结果一致,可能是当接种量较大时,菌体前期生长迅速,培养液中营养物质被迅速消耗,同时产生大量代谢废物,抑制中后期菌体的生长。同时酵母菌在生长过程中会产生大量的乙醇,随着乙醇含量的升高,培养液中pH值降低也会抑制菌株的增殖使蛋白酶的活性降低。将五株菌株加入到不同pH的培养液中,随pH值的升高蛋白酶活也逐渐增加,五株菌均在pH值9的培养基中最高,达到200 U/mL以上,结合先前的报道可知,五株菌产生的均为碱性蛋白酶。在30~55 ℃蛋白酶的活性较高,筛选出的产蛋白酶酵母菌株库德毕赤酵母Y1、单孢酿酒酵母Y21的最佳培养温度为30 ℃,菌株东方伊萨酵母Y5、马克斯克鲁维酵母Y16、近平滑假丝酵母Y20 最佳培养温度为37 ℃,温度对产蛋白酶的活性也具有显著的影响。
3.2 微生物产复合酶的研究
微生物由于繁殖速度快、发酵简单、易于大规模生产等优点,目前成为蛋白酶、纤维素酶、植酸酶、果胶酶等生物酶类的重要来源。植酸盐是谷类、豆类和油料中磷酸盐和肌醇的主要储存形式。磷以肌醇六磷酸的形式存在不能被人类、家禽或其他单胃动物利用。为了提高膳食植酸磷的利用率,不同来源的植酸酶一直是研究的热点。Li 等[16]从鱼类的肠道中分离出可以分泌大量植酸酶的海洋酵母菌株,该酵母的最佳产酶条件为pH5.0 和28 ℃。崔明玉[17]从酵子和全麦面团中筛选分离出具有较高植酸降解能力的一株异常威克汉逊姆酵母和一株酿酒酵母,其植酸酶活性分别为18.52 U/mL 和19.74 U/mL。本研究中筛选分离出的单孢酿酒酵母Y21植酸酶活性为15.63 U/mL,可能跟菌液的起始浓度等因素有关。
果胶酶属于多糖家族,可以降解果胶物质。果胶酶在工业生产中占据领先地位,具有广泛的应用前景,如植物纤维的脱胶、榨油,果汁和葡萄酒的澄清,生物燃料的生产,动物饲料的生产等。但像许多其他工业酶一样,果胶酶在其经济生产中也面临着低产量和高生产率的限制[18]。据报道,果胶酶的50%来源是真菌和酵母菌,35%来源是细菌。近年来,已经筛选分离出了许多具有果胶酶活性的微生物,如可以生产果胶酶的黑曲霉菌和曼赫毛菌。此外,从土壤和牛粪便中筛选出的芽孢杆菌也能高效的生产果胶酶[19]。Karabi 等[20]从土壤中筛选出了产果胶酶的细菌在37 ℃和pH 7.5 的条件下产生最大的果胶酶活性0.671 U/mL。本研究中果胶酶活性达到15.18 U/mL,具有较高的果胶酶活性。果胶酶的活性高低可能和菌属有关,本试验筛选分离得到的菌株为酵母菌。
纤维素酶可以通过酶的协同作用将纤维素有效地水解为葡萄糖单位,将植物细胞中平常不消化以及难以消化的大分子物质,如大分子多糖、蛋白质和脂类等物质分解成小分子容易被动物吸收的物质。这些酶被称为内切β-1-4-葡聚糖酶、纤维二糖水解酶和β-D-葡萄糖苷酶[21]。有研究表明,几乎所有酵母菌种均具有胞外纤维素酶活性,大多数酵母中纤维素酶活性在30 ℃时最高[22]。因此本研究对菌株在30 ℃产蛋白酶活性进行了探讨,五株菌株均表现出较高的纤维素酶活性,均超过180 U/mL,其中近平滑假丝酵母Y20 酶活力显著高于其他菌株(P<0.05),达到199.08 U/mL。
具有产蛋白酶、果胶酶、植酸酶、纤维素酶的菌株对菜籽粕等进行发酵,不仅能够为动物提供所需的营养物质,也是含有多种酶的复合酶制剂。复合的酶制剂具有互补作用,在多种酶的共同作用下,饲料中的一些抗营养因子会被破坏,可显著促进动物生长,提高机体免疫力。据报道,将复合酶添加到不同动物的基础日粮中,动物生产性能均显著提高[23]。本文研究了从山羊瘤胃液中筛选出产蛋白酶的酵母菌并分析了影响产酶因素以及纤维素酶、植酸酶、果胶酶的活性,其结果将为杂粕类发酵饲料提供科学依据。
4 结论
①本试验筛选出的5 株产蛋白酶菌经形态学分析及分子生物学(26S rDNA序列分析)鉴定菌株Y1为库德毕赤酵母,Y5为东方伊萨酵母,Y16为马克斯克鲁维酵母,Y20为近平滑假丝酵母,Y21为单孢酿酒酵母。
②对各菌株产酶条件进行优化,在最佳产酶条件下,菌株库德毕赤酵母Y1(时间48 h、接菌量3%、温度30 ℃、pH 值9)酶活力为229.89 U/mL,东方伊萨酵母Y5(时间48 h、接菌量4%、温度37 ℃、pH值9)酶活力为212.63 U/mL,马克斯克鲁维酵母Y16(时间48 h、接菌量3%、温度37 ℃、pH值9)酶活力为208.44 U/mL,近平滑假丝酵母Y20(时间48 h、接菌量4%、温度37 ℃、pH 值9)酶活力为211.74 U/mL,单孢酿酒酵母Y21(时间48 h、接菌量4%、温度30 ℃、pH 值9)酶活力为209.96 U/mL。对复合酶进行测定,库德毕赤酵母Y1、马克斯克鲁维酵母Y16、单孢酿酒酵母Y21 具有较高的植酸酶活性。五株菌株均表现出较高的果胶酶活性、纤维素酶活性。
猜你喜欢
食品工业(2022年3期)2022-11-16
新农业(2022年13期)2022-07-18
中国饲料(2022年13期)2022-07-07
食品工业(2022年2期)2022-03-09
食品安全导刊(2021年29期)2021-11-10
食品与机械(2020年8期)2020-09-18
中州大学学报(2020年2期)2020-05-31
中国畜牧杂志(2019年12期)2019-12-26
新农业(2019年16期)2019-09-05
湖南饲料(2019年3期)2019-06-12
