
基于流感发病机制的药物研究进展
2021-05-07 00:54陈金凤
中国药理学通报 2021年5期
陈金凤,何 军,徐 涛
(1.安徽医科大学药学院,安徽 合肥 230032;2.安徽省疾病预防控制中心,安徽 合肥 230601)
流感是由流感病毒引起的急性呼吸道传染病,每年影响5%-15%的人口并造成30万-65万人死亡[1]。流感病毒耐寒不耐热,通过吸入打喷嚏、咳嗽在空气中形成的气溶胶或者接触携带病毒的物品导致感染。常见一些呼吸道症状并伴随全身酸痛乏力、食欲减退等全身症状,少数人还会出现腹痛、腹泻等消化道症状,严重者可能出现肺炎、脑膜炎、急性呼吸窘迫综合症、休克等并发症,进而导致多脏器功能衰竭甚至死亡。
流感病毒通过攻击宿主上皮细胞完成复制进而感染更多细胞,触发免疫系统攻击和破坏整个呼吸系统受感染组织并导致免疫系统过度反应。因此抗流感不仅要抑制病毒复制,更要注意阻断相关炎症因子在免疫细胞间传递并缓解机体过度免疫反应,减轻修复免疫反应带来的机体损伤。除了常用的NA抑制剂和M2阻断剂外,目前也在病毒复制和机体免疫的其他靶点方向上开发新药。中药一直以来以其多靶点治疗闻名,研究发现中药中很多活性成分如醌类、类黄酮、生物碱等都可有效抵抗流感。此外,很多研究还发现中西药联合使用抗流感比单用治疗效果好。本文就近些年对抗流感研究有效果的药物进行归纳总结,为未来抗流感药物的开发提供依据。
1 流感病毒
流感病毒根据病原特点分为4种,甲型流感病毒易发生抗原性转换和漂移,其传染性最强、死亡率最高,并可大规模爆发和流行。乙型流感病毒发生变异较少,能引起季节性流行但很少引起大流行。丙型流感病毒抗原性高度稳定无再分亚型,通常只会引起不明显或者轻微的上呼吸道感染,很少造成流行。近年来才发现的丁型流感病毒结构最稳定且无再分亚型,其中人易感甲型和乙型。以下就流感病毒的发病机制展开叙述。
1.1 流感病毒的复制流感病毒结构由外到内依次是糖蛋白、包膜、基质蛋白、核心,最外层糖蛋白包括血凝素(hemagglutinin,HA)和神经氨酸酶(neuraminidase,NA),病毒核心包括ss-RNA、核蛋白(nucleoprotein,NP)以及RNA聚合酶。病毒进入呼吸道后HA水解为轻链和重链,重链与宿主上皮细胞上的唾液酸受体结合,轻链协助病毒包膜与宿主细胞膜融合内吞进入细胞。随后病毒基因组在宿主细胞内复制、编码并组建新的病毒子代。成熟的流感病毒子代以出芽的方式脱离宿主细胞时HA会与其表面的唾液酸受体结合而无法脱离,此时NA将唾液酸水解,切断病毒与宿主细胞最后的联系。流感病毒通过扩散感染造成细胞变性、坏死乃至脱落,进而导致呼吸道充血、水肿及分泌物增加,产生鼻塞流涕、咽喉肿痛、咳嗽等呼吸道症状。
1.2 机体非特异性免疫流感病毒进入机体内环境开始感染细胞,而感染死亡的细胞在凋亡前会释放相关的细胞因子并通过其浓度差召唤巨噬细胞(macrophage,Mφ)、嗜中性粒细胞(neutrophilic granulocyte,NE)等吞噬病毒和受感染细胞,NE根据组织液中信息分子浓度差到达感染处,并在其刺激下进入狂暴状态,大肆吞噬病毒、被感染细胞并释放强烈的毒素除掉周围大量的正常细胞防止感染蔓延,因此牵扯很多无辜细胞损伤。如果流感病毒顽强,战斗持续进行还会产生更多的炎症因子,中枢神经检测到炎症因子达到一定量级时会作用于下丘脑后部的COX-2合成PGE2等激素[2]并释放至全身,此时机体产热增加、散热减弱通过升高体温来抑制病毒生存和繁殖,即机体表现出发热的全身性症状。
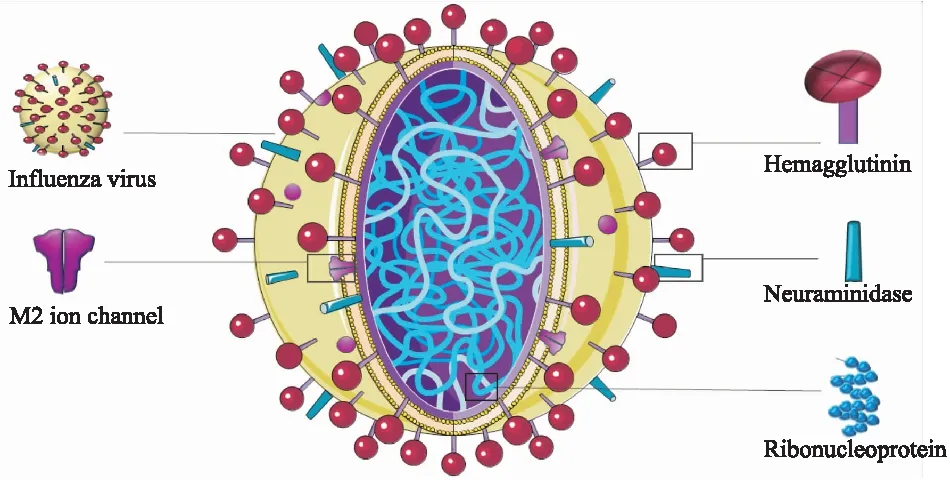
Fig 1 Structure diagram of influenza virus
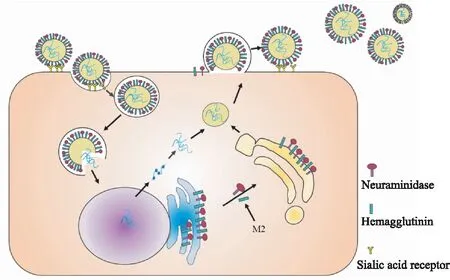
Fig 2 Replication process of influenza virus
1.3 机体特异性免疫若先天性免疫不可控,Mφ、树突状细胞(dendritic cell,DC)、杀伤T细胞(T cycocotic cell,Tc)、辅助T细胞(T helper cell,Th)、B细胞等在细胞因子层层传递下运作产生抗体。抗体具有多个位点可标记抗原通过一系列操作破坏病毒活性并通过类似于氢键作用直接将流感病毒向Tc吸附。但是无论是DC在上亿种T细胞中寻找能特异识别病毒的T细胞还是Th寻找B细胞都是需要时间的,免疫细胞根据感染处炎症因子浓度差来定位,若炎症反应持续,几轮内循环后炎症因子浓度差不再明显,机体因持续性抗击病毒失利释放更多的激素加速免疫细胞和体细胞释放炎症因子,试图让机体调动所有的免疫力量展开最后的攻击,直至形成炎症因子风暴。炎症因子因体循环遍布各大器官已经失去导向作用,导致NE开始攻击所有炎症因子出现的地方,在各个地方释放杀伤流感病毒的毒素、大面积收缩血管并破坏正常细胞的功能,机体出现器官衰竭、休克甚至死亡等情况。
2 抑制流感病毒表面蛋白NA的药物
蘑菇形四聚体糖蛋白NA的单体由固定在病毒膜上的茎结构域和催化活性中心球形头构成,其活性位点由8个功能性残基(R-118、D-151、R-152、R-224、E-276、R-292、R-371和Y-406)组成,被11个骨架残基环绕(I-222、E-119、R-156、E-277、W-178、S-179、D-198、E-227、H-274、N-294和E-425)。NA不仅能裂解HA与唾液酸之间的糖苷键来催化唾液酸水解,还通过降低呼吸道黏液层粘度使细胞表面受体暴露增强病毒吸附[3],此外还促进含病毒组织液散布至下呼吸道短期内使呼吸道受损。因此NA是目前开发及应用最多的靶标(Tab 1)。

Tab 1 NA inhibitors and mechanisms
2.1 化药及小分子化合物神经氨酸酶抑制剂(neuraminidase inhibitors,NAIs)是当前应用最广泛的抗病毒药物,通过与NA活性位点可逆性结合而与神经氨酸(底物)竞争,抑制NA的酶功能进而切断病毒的扩散链,阻止病毒子代从宿主细胞表面脱落。扎那米韦与NA活性位点E-119形成氢键,因生物利用度差被设计为吸入粉雾剂在呼吸道中局部浓缩用药。第一个口服的抗流感西药磷酸奥司他韦可结合NA活性位点H275Y,其中R292K、E119V和H274Y是该药常见的耐药突变点。帕拉米韦通过其羧基、胍基结构与NA活性位点(包含Asp151、Glu119和Glu227等)形成强烈的分子间作用抑制NA活性,R378Q,R378K和R378L是其耐药突变点,由于其生物利用度差,目前是经FDA批准的唯一通过静脉注射给药的抗流感药。辛酸拉尼米韦对磷酸奥司他韦的耐药株有较好的抑制作用,其最大优势是可在肺部停留较长时间保持长效抑制,每周只需吸入一次即可有效抗流感[4]。
2.2 中药活性成分及复方植物挥发油中提取的单环倍半萜吉马酮可剂量依赖性抑制流感病毒的复制转录及NA活性[5],与奥司他韦联用在体内外均表现出对病毒感染抑制的累加作用,有潜力单独或与其他药物联合开发用于流感的治疗。黑色接骨木莓中提取的黄酮类化合物矢车菊素-3-桑布双糖苷(cyanidin-3-sambubioside,C3S)可作为病毒H274Y突变的潜在抑制剂,与H274Y突变型流感病毒形成类似于氢键的作用力并显示出良好的结合亲和力[6],与耐H274Y的奥司他韦联用具有相当强的抗病毒活性。此外黄连解毒汤水提取物中黄连碱、小檗碱、栀子苷等以竞争性方式有效抑制NA-1[7]。
3 抑制流感病毒表面蛋白HA的药物HA是由球状头部和茎区域组成的穗状均三聚体,主要介导流感病毒导入宿主细胞,包括病毒-细胞结合和病毒-宿主膜融合。位于HA头部的受体结合位点与细胞表面的唾液酸结合,通过内吞作用使得病毒进入。病毒进入后在酸性环境下诱导HA茎区构象发生变化,从而导致病毒膜与宿主内体膜融合并释放病毒RNA基因组到细胞质中(Tab 2),HA介导病毒进入这一关键步骤目前已成为抗流感药物开发的潜在目标。

Tab 2 HA inhibitors and mechanisms
3.1 化药及小分子化合物已在俄罗斯、中国被批准用于甲、乙型流感治疗的广谱抗病毒药阿比朵尔通过发挥分子胶功能[8]与流感病毒的HA蛋白相互作用使其在低pH过渡到融合状态时稳定下来抑制HA介导的膜融合即低pH诱导的HA重折叠过程,还有效抑制病毒引起的氧化应激,此外研究还发现在阿比朵尔的苯硫酚部分添加间羟基以取代结合袋中的结构化水分子可显著增加对H3(1150倍)和H1(98倍)亚型的亲和力。化合物CBS1116同样能够干扰HA介导的膜融合,通过与HA茎区域结合并干扰低pH触发的HA构象变化。此外化合物CR6261也是通过类似CBS1116的作用方式结合HA的茎区域来发挥抗病毒作用。
3.2 中药活性成分枳的种子提取物在流感病毒复制早期阶段可阻止病毒附着、酸化、HA介导的细胞-细胞融合和脱壳[9],橄榄主要的萜类活性成分橄榄苦苷可靶向HA2蛋白[10],二者都能有效阻止流感病毒与细胞结合。
4 抑制流感病毒M2离子通道的药物
M2离子通道是流感病毒基因组解包和病毒解壳过程中RNA释放的基础,在质子作用下还可被病毒识别进出宿主细胞。此外还能阻断细胞自噬体与溶酶体融合来抑制细胞自噬降解。虽然当前相关的临床药物几乎完全耐药,但在促进感染细胞自噬方面仍有开发新药的潜力(Tab 3)。

Tab 3 M2 ion channel inhibitors and mechanisms
第一代抗甲流药物即M2离子通道抑制剂金刚烷胺和金刚乙胺,其在低浓度下可特异性抑制M2离子通道活性,高浓度下可非特异性提高宿主细胞内pH从而抑制或延迟酸诱导的病毒HA构象变化。大多数正在传播的人类流感病毒在M2跨膜孔结构域中携带抗药性突变(例如S31N,V27A和L26F)已导致两药近100%耐药,且因能透过血脑屏障具有潜在的中枢神经系统毒性[11],疾病控制与预防中心不再推荐二者用于抗流感。金刚烷胺衍生物金刚烷溴噻吩通过靶向S31-M2和N31-M2通道孔,抑制病毒进出宿主细胞来抗流感。化合物J10688是一种宿主cdc2样激酶1(cdc2 like kinase 1,CLK1)抑制剂,CLK1负责在流感病毒感染和复制过程中选择性剪接M2基因,而J10688显著下调剪接因子SF2/ASF和SC35的磷酸化进而调节病毒M2基因的选择性剪接,在体内外均表现出抗流感病毒活性。化合物TCN-032通过特异性结合M2保守的胞外域,阻止病毒出芽脱离以缓解流感症状[11]。
5 抑制流感病毒NP的药物
流感病毒结构蛋白NP在病毒复制中起着核心作用,NP是vRNP 的骨架结构,功能涉及RNA堆积、核运输和病毒RNA转录与复制等。在四型流感病毒株中NP的折叠基本是保守的,这意味着病毒不太容易对NP抑制剂产生抗性,故具有很不错的开发前景(Tab 4)。

Tab 4 NP inhibitors and mechanisms
F66是第一个被报道靶向NP的RNA结合沟抑制剂,其可能是与R174-K184表位区域的RNA结合槽结合而抑制病毒的复制转录。萘普生也是靶向NP的RNA结合沟抑制剂,不仅能直接抑制流感病毒活性,还可抑制病毒触发的炎症反应。微型基因组测定法发现RK424通过抑制流感病毒vRNP活性进而破坏病毒RNA诱导的NP寡聚,还与单体NP结合破坏NP-RNA和NP-NP的相互作用[12]。小分子化合物Nucleozin通过Y289/N309和Y52/Y313口袋从下至下桥接两个NP分子,阻止NP导入细胞核并引起细胞质内异常聚集来抑制流感病毒复制。化合物S119-8通过改变NP蛋白的寡聚状态来抑制流感病毒复制,与奥司他韦联用具有协同作用并降低耐药性[13]。
6 抑制流感病毒RNA聚合酶的药物
流感病毒RNA聚合酶是由PA、PB1和PB2亚基构成的蛋白质复合物,PA具有内切核酸酶活性并可诱导蛋白水解,PB1具有RNA聚合活性,PB2识别并结合宿主5'mRNA帽结构在基因组复制转录中发挥着重要作用。当前临床上RNA聚合酶抑制剂突变频率低且不良反应较轻,其已成为抗流感药物研究的重要靶标(Tab 5)。

Tab 5 RNA polymerase inhibitors and mechanisms
6.1 化药及小分子化合物鸟苷核糖核苷类似物利巴韦林主要靶标为肌苷5'-单磷酸脱氢酶(inosine monophosphate dehydrogenase,IMPDH),通过降低细胞内GTP浓度和阻止病毒蛋白质合成来抑制病毒RNA复制且可引起RNA复制突变,与甘草酸联用可显著降低炎症因子IL-6、TNF-α和IL-1β的表达水平。已进入Ⅲ期临床试验的匹莫地韦靶向RNA聚合酶的PB2亚基并通过阻止聚合酶结合宿主前体mRNA上的7-甲基GTP帽结构抑制病毒基因的表达,对多种甲型流感病毒及NAIs敏感性低的毒株具有活性。化合物D715-2441通过用作帽结合蛋白结合竞争者与RNA聚合酶中PB2蛋白特异性结合并影响其位置进而抑制病毒复制周期的早期步骤,对多种亚型(H1N1、H5N1、H7N9、H3N2)具有抗病毒活性。当前已在美国和日本上市的巴洛沙韦靶向病毒PA亚基的核酸内切酶活性,一旦它被PB2.8结合就可以阻止聚合酶切割前体mRNA,对甲、乙型流感病毒甚至耐奥司他韦的病毒均有效。嘌呤核苷类似物法匹拉韦几乎对所有流感病毒均具有活性,其在细胞内被核糖基化和磷酸化后形成药物活性形式核苷三磷酸化物法匹拉韦RTP,这种形式被病毒RNA聚合酶错误识别作为嘌呤误掺入新生RNA中并在RNA合成过程中导致链终止[14],此外还通过诱发致命性的突变来发挥抗病毒作用,由于新冠肺炎疫情等原因该药已在我国上市。
6.2 中药活性成分异槲皮苷可阻断RNA聚合酶中PB2运作来阻止病毒复制,还能减少甲流感染产生的活性甲氧基。水飞蓟种子中提取的水飞蓟素主要成分水飞蓟宾、水飞蓟宁等可阻止病毒进入宿主细胞并调节自噬抑制氧化应激物的形成[15],还可触发细胞外信号调节激酶(extracellular signal regulated kinase,ERK)/p38丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和IκB激酶(inhibitor of nuclear factor kappa-B kinase,IKK)级联,由于水飞蓟素溶解性差,目前探索了β-环糊精包合、成盐、糖苷衍生物、脂质体递送等方法来提高其生物利用度。川楝子的主要活性成分川楝素与PA的结合亲和力可能强于已知PA抑制剂的结合亲和力,其通过改变PA蛋白的核定位破坏核转运导致PA的胞质积累并抑制流感病毒mRNA早期复制,此外还通过抑制蛋白激酶B活性来干扰病毒进入宿主细胞[16]。
7 缓解机体免疫过度的药物
抗炎免疫是机体对抗流感病毒侵袭必不可少的一步,炎症因子作为免疫的信号分子通过NF-κB、Nrf2、JAK-STAT、RLRs、AMPK等信号通路释放进行机体自我防卫。但流感症状几乎也都是由炎症因子造成的,如何在免疫前期促进信号分子传导积极辅助抗病毒药物治疗,在免疫后期特别是发展成重症流感时抑制炎症反应,需要科学的判断并合理用药。当前主攻方向除了抑制流感病毒复制和扩散,还应重点研究如何阻断炎症因子在免疫细胞间传递来抵御人体过度免疫(Tab 6)。

Tab 6 Anti-inflammatory drugs and mechanisms in anti-flu course
7.1 化药及小分子化合物静脉免疫球蛋白(intravenous immunoglobulin,IVIG)含有IgG抗体,具有免疫替代和免疫调节双重治疗效果[2,11],可降低重症流感相关的死亡率和发病率,在流感重症监护环境中常作为第一阶段干预措施。过氧化物酶体增殖物激活受体γ(peroxisome proliferators-activated receptors γ,PPARγ)具有增强脂质和葡萄糖的代谢以及细胞分化和抑制炎症等功能,在机体免疫过度时可减少NE和Mφ数量并减少炎症因子的产生[2]。N-乙酰半胱氨酸可抑制髓过氧化物酶活性,减少NE、Mφ数量及促炎因子水平来减少肺部炎症和肺水肿[17]。
7.2 中药活性成分、复方及中成药瓜子金皂苷可有效降低肺组织中 IL-1β、TNF-α、IL-4、IFN-γ、TXA2和PGE2的水平,通过减轻肺部炎症反应来增强抗甲流感染的能力。穿心莲内酯被誉为天然抗生素药物,其抗流感病毒活性是通过参与NF-κB、Nrf2、JAK-STAT和RLRs信号通路的激活降低病毒载量和感染引起的炎性细胞因子表达并最终减少流感病毒诱导的细胞死亡,此外由穿心莲内酯作为主要成分的喜炎平注射液联合奥司他韦用药普遍具有良好的临床疗效。桔梗皂苷通过抑制NF-κB信号通路来抑制Cox-2和一氧化碳合酶的表达并提高NK细胞的杀伤活性[18],此外还通过抑制细菌生长缓解流感并发症。中药甘草活性成分包括甘草甜素、甘草次酸、异甘草素等,甘草甜素在IL-1信号通路中作用于皮质类固醇11-β-脱氢酶同工酶1进入宿主细胞,阻碍单核细胞募集并降低促炎分子(CXCL10、IL-6、CCL2和 CCL5)水平,激活IFN-γ并降低 NF-κBp38和JNK的表达[19],具有抗氧化活性可抑制流感病毒诱导的活性氧(ROS)的形成,此外还可以用作制备抗流感疫苗的佐剂;异甘草素在PPAR途径的激活下抑制炎症因子基因表达和炎症细胞募集来缓解肺部炎症。柴胡皂苷对许多炎症过程(毛细血管通透性增加、炎症介质释放、白细胞迁移和结缔组织增生)都有影响,柴胡皂苷A可显著抑制三磷酸腺苷诱导的血小板凝集缓解流感引起的出血现象[10]。经典名方葛根汤通过促进 IL-12 及 IFN-γ的产生来诱导 Th1 系统免疫应答,降低Toll样受体7(Toll like receptor 7,TLR-7)信号通路和TNF-α的表达并改善Th1/Th2免疫平衡从而减轻过度免疫反应[20]。参附注射液通过抑制高迁移率族蛋白B1(high mobility group protein B1,HMGB1)的转录、表达、易位和分泌,增加TLR-4信号的表达和IκB-α产生的同时降低肺中p65、p50、TNF-α和IL-1β的表达水平从而有效预防了MAP的下降并减轻了肺组织损伤,即通过抑制HMGB1-NF-κB途径预防炎症因子风暴发生[21]。
8 多靶点抗流感的药物
部分抗流感药物通过多成分、多靶点和多通路作用,达到抑制流感病毒复制和抗炎免疫的双重药效作用,当前此类药物多集中在一些待开发的中药活性成分,其在抗病毒药物研发上提供了很多新的思路和方法(Tab 7)。

Tab 7 Multi-target drugs and mechanisms
8.1 化药及小分子化合物盐霉素是甲、乙型流感病毒有效的杀伤剂,通过阻断NP的核迁移在病毒进入过程中特别是在核周内体中引起NP的胞质积累,还可阻止内体酸化并失活M2质子转运功能[22],此外与奥司他韦联用能显著提高疗效,可用于辅助治疗对现有药物耐药或敏感性较低的流感。硝唑尼特是一类口服抗寄生虫药但同时具有抗病毒和免疫调节作用,通过抑制HA的成熟与翻译进一步靶向流感病毒并上调IFN和各种IFN诱导型基因发挥抗炎作用,与奥司他韦组合也具有协同增效的作用[23]。
8.2 中药活性成分及中成药儿茶素类化合物在流感病毒结构保守的邻近残基430空腔附近[24](次唾液酸结合位点)与NA结合抑制病毒脱离,还以剂量依赖性方式显着降低自噬相关蛋白LC3B的含量并通过影响HA的构象发挥抗流感病毒的作用[10],天然儿茶素甚至可以用作制备灭活病毒疫苗的灭活剂。多酚类物质姜黄素干扰HA与宿主细胞受体结合且其衍生物与HA所有亚型都具有显著的结合亲和力,还通过中断Y148和RNA碱基之间的堆叠作用降低NP的RNA结合亲和力,还可明显抑制流感病毒诱导的氧化应激并通过激活Nrf2信号抑制流感病毒诱导的TLR2/4,NF-κBp38/JNK和MAPK信号的激活,这些过程可能依赖于姜黄素自身结构中两个烯酮基团[25];单乙酰姜黄素具有与姜黄素相同烯酮基团,除了对HA没有影响外其他功效均与姜黄素相同且能强烈抑制Akt磷酸化。
芦荟蒽醌类成分芦荟苷通过抑制流感病毒NA并阻止NA介导的TGF-β的活化来抑制由TGF-β介导的免疫反应,提高病毒HA特异性T细胞的免疫力,使得肺中HA特异性Th和Tc有更多的浸润并恢复NS1抑制的STAT1介导转染细胞中的抗病毒应答且增强了IFN-γ和TNF-α的产生;其黄酮类成分槲皮素、儿茶素和山奈酚等可显著抑制病毒mRNA中M2的相关节段合成并通过与关键功能位点(PHE47A和LEU43A)的结合来抑制M2蛋白的表达,此外还通过抑制流感病毒诱导的自噬对H1N1或H3N2产生抗病毒活性;芦荟多糖通过活化Mφ产生NO并分泌细胞因子(TNF-α、IL-6和IFN-γ)来介导免疫调节作用,同时也有抑制M2的作用[26]。黄芩的黄酮提取物包括黄芩素、汉黄芩素、木蝴蝶素及糖苷类等[27],黄芩苷可阻止H3N2抑制的mTOR信号传导途径诱导的自噬,还可诱导Th、Tc和NK细胞中的IFN-γ产生并激活JAK-STAT1信号通路;汉黄芩素可抑制流感诱导的AMPK磷酸化和激活,在I型和Ⅲ型IFN的诱导下通过IRF3的磷酸化和IL刺激基因(如MxA和OAS)的激活来增强抗病毒信号传导进而减轻炎症反应;黄芩的黄酮提取物不仅能抑制HA、NA活性,降低TNF-α、IL-6和 MCP-1的表达并增加肺组织中IFN-γ和IL-10的表达水平调节炎症反应,还能通过影响肠道菌群间接帮助身体恢复,在甲流诱导的肺损伤治疗中提取物比单一的黄芩素有更佳的治疗效果;此外黄芩参与的组方很多,如三物黄芩汤、双黄连,其中双黄连另两味中药金银花和连翘中的咖啡酸衍生物与抗甲型流感病毒密切相关。体外实验发现板蓝根水提物S-03具有抑制HA[28]和增加抗氧化酶活性的作用,例如SOD,CAT,GSH-Px和总抗氧化能力;板蓝根多糖提取物通过激活TLR-3信号通路削弱流感病毒诱导的促炎因子的上调并显着降低IL-6,IP-10,MIG和CCL5的表达,对人流感病毒(H1N1和H3N2)和禽流感病毒(H6N2和H9N2)具有抑制作用。
源于经典名方“桑菊饮”的夏桑菊颗粒可抑制NP的核输出,通过抑制NF-κB信号通路激活来抑制IKKα、IKKβ、NF-κBp50、NF-κBp65 的磷酸化并减少炎症因子的基因表达。体外实验发现复方一枝蒿颗粒对甲、乙型流感具有广谱抗病毒作用,其活性成分绿原酸和异甘草素具有抑制NA的活性,活性物质雪松酸、芥子苷、木犀草素和荭草苷等都对TNF-α有抑制作用,特别是木犀草素降低TNF-α含量作用最显著,可减少肺部浸润和肺损伤[29]。
9 结语与展望
流感一直是人类和动物生存面临的一个巨大威胁,由于优先使用NA抑制剂的治疗策略,现面临着层出不穷的耐药株,部分药物还具有严重的毒副作用如金刚烷胺类中枢神经系统毒性,更现实的问题在于在耐药株层出不穷的状况下许多有疗效的化合物还停留在筛查试验阶段,相关药物的临床试验及上市还需要很长一段时间观察。而中药的多靶点作用机制尚不明确,很多药物仅是体外实验有效果体内作用机制未知且用于流感治疗时依靠的只是长年累月的用药经验,部分中药成分还存在溶解性差、稳定性差、生物利用度低等问题。当前抗病毒研究主要分为两个方面,一是针对现有靶标来优化或设计新的药物,如针对已耐药的M2通道靶点,可考虑在其蛋白质二硫键结构上进行新的药物设计。二是寻找新的靶标,开发新的抗病毒药物。如由RNA聚合酶协同作用产生的独特“抓帽”机制;针对流感病毒突变产生耐药性等问题深入探究病毒相对保守的基因序列以及一些非编码RNA,如circRNA(参与抑制细胞自噬和促进病毒复制)和miRNA等,这些对病毒有影响的新靶标为设计新颖的抗病毒抑制剂提供了方向。此外,文章虽列举了部分针对炎症因子相关的药物,但其在体内更明确的作用机制还有待进一步探索。在接下来的抗流感药物研发上可充分利用网络拓扑学理论对生物信息和蛋白质相互作用节点分析进行药物作用靶点筛选[30],减轻筛查试验工作量。随着现代研究技术的不断进步和对流感的深入学习研究,未来定能构建一个完整的抗流感药物库有效应对流感大流行。
猜你喜欢
科学(2020年3期)2020-11-26
基层中医药(2020年4期)2020-09-11
基层中医药(2020年12期)2020-07-22
当代水产(2020年3期)2020-06-15
广东医科大学学报(2020年6期)2020-02-06
现代检验医学杂志(2016年3期)2016-11-15
现代检验医学杂志(2016年1期)2016-11-12
华南农业大学学报(2015年5期)2015-12-04
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
