
龙眼DCL4的亚细胞定位分析及其对外源ABA和SA的响应
2021-08-04 07:31张雪莹梁梓豪陈晓慧张舒婷赖钟雄林玉玲
热带作物学报 2021年6期
关键词:龙眼
张雪莹 梁梓豪 陈晓慧 张舒婷 赖钟雄 林玉玲

摘 要:为了研究DlDCL家族中DlDCL4基因在龙眼中的功能,本研究通过氨基酸序列比对、系统发育进化树的构建、亚细胞定位验证、实时荧光定量PCR(qPCR)等方法,对其进行初步探究。結果表明:遗传进化分析显示,DCL4氨基酸序列在所选的物种间相似度高于50%,其中龙眼与甜橙(Citrus sinensis)的亲缘关系最近。在激光共聚焦显微镜观察下,洋葱细胞核发出绿色荧光,其他部位不发出荧光,表明龙眼DCL4蛋白定位于细胞核中。在24 h内,100 μmol/L的外源脱落酸(ABA)对DlDCL4都有极显著的下调效果;100 μmol/L的水杨酸(SA)对其表达量有显著的上调效果,其中8 h时表达量最大,提示DlDCL4对不同激素的响应效果不同且不同处理时长下响应效果也会有影响。
关键词:龙眼;DCL4基因;亚细胞定位;激素处理
中图分类号:S667.2 文献标识码:A
Subcellular Localization Analysis and Responses to Exogenous ABA and SA in Longan Callus of DCL4
ZAHNG Xueying*, LIANG Zihao*, CHEN Xiaohui, ZHANG Shuting, LAI Zhongxiong**, LIN Yuling*
Institute of Horticultural Biotechnology, Fujian Agricultural and Forestry University, Fuzhou, Fujian 350002, China
Abstract: This study carried out amino acid sequence alignment, phylogenetic tree analysis, subcellular localization verification and real-time qPCR analysis to study the function of DlDCL4 gene of DlDCL family in longan. It was found that the similarity of the amino acid sequence of DCL4 was more than 50% in the selected species. In the evolutionary relationship, DlDCL4 and navel orange (Citrus sinensis) were closely related. Under the observation of laser confocal microscopy, the onion cell nucleus emitted green fluorescence, and other parts did not emit fluorescence, so the longan DCL4 protein was localized in the nucleus. In 24 h, 100 μmol/L exogenous ABA hormone had a significant down-regulation effect on DlDCL4. SA had a significant up-regulation effect on its expression, and the effect was higher than other time treatment at 8 h, which suggested that DlDCL4 had different responses to different hormones and this effect would also be affected by different treatment durations.
Keywords: longan; DCL4 gene; subcellular localization; hormone treatment
DOI: 10.3969/j.issn.1000-2561.2021.06.002
Dicer是一种高度保守的核糖核酸酶,在植物中称为Dicer-like(DCL)蛋白,属于RNase Ⅲ家族,是RNA沉默途径中至关重要的组分,其通常包括DEAD box、helicase-C、DUF283、PAZ、RNase Ⅲ和dsRBD 6个结构域[1]。DCL蛋白是加工dsRNA前体生成小分子RNA的关键酶,可通过剪切dsRNA产生siRNA,启动RNAi途径的起始阶段,在植物RNA沉默机制中发挥了关键作用[2]。由于DCL结构域的多样性,不同的DCLs在不同的生物中对RNA的剪切功能存在差异[3]。每个DCL蛋白都有其各自的功能,彼此通过功能互补和自我平衡的方式发挥作用[4-6]。研究发现,在拟南芥中,AtDCL4主要在转录后的基因沉默发生作用,通过特异性识别大于100 nt的长链dsRNA,将其剪切形成20 nt的反式作用siRNA;在水稻中,OsDCL4剪切产生21 nt的反式siRNAs和phasiRNAs4[7-8]。研究表明DCL4在调控植物生长发育和适应性方面也发挥了一定生物学作用。氮缺乏条件下,拟南芥突变体ski2 dcl4和ein5 dcl4中22 nt siRNAs被显著诱导,韧皮部发育相关基因CALS7和NEN4显著下调表达,土壤根系生长明显被抑制,幼苗发育迟缓[9];在棉花干旱胁迫和病毒侵染过程中,DCL4被诱导显著性上调表达[10];撇除水稻OsDCL4基因导致植株生长矮小、结穗率低,植物内源dsRNA含量减少而使植株易受病毒侵染[11]。
外源脱落酸(ABA)和水杨酸(SA)都是植物内源性激素,对植物的生长调节有重要作用。ABA在植物的生长发育调节和对非生物胁迫的应答方面发挥了显著的作用[12]。研究表明,ABA对植物生长发育的调节主要体现在促进器官脱落和休眠、抑制植物生长、促进种子休眠等方面[13],同时还可以提高植物对于干旱、低温和高盐抗性[14]。在龙眼的相关研究中发现ABA可以提高RNA沉默相关基因AGO10的表达量[15],但DlDCLs下调表达[16]。SA在植物中主要是作为防卫激素发挥作用[17],目前在多个物种中发现SA可以增强对生物性病原的抵抗作用,而且时常被应用于种子萌发前的催种[18]。研究表明,SA信号转导途径在植物对抗逆境胁迫,尤其是在对抗病害的胁迫过程中起着关键性作用[19-21]。同时,研究发现50、75 μmol/L浓度的SA可以使龙眼Mn-SOD的表达量显著上调,提高龙眼的抗氧化能力[22],同时对龙眼抗病基因的表达有调控作用[23]。
龙眼(Dimocarpus longan Lour.)作为一种热带亚热带水果,具有非常高的经济价值,同时龙眼干还有着很高的药用价值,所以龙眼果实的品质调控一直都是研究的热点。ABA和SA在植物的生长和发育中有重要的作用,而DCLs作为miRNA作用过程中的关键酶也越来越受到重视,探究它们之间的作用机制对提高龙眼果实品质具有重要意义。近年来,研究发现DlDCLs对龙眼的叶子和花器官发育具有重要的调控作用,并且能响应干旱、低温和ABA、SA等非生物胁迫[16]。研究表明,在不同种类外源激素处理下,DlDCL4的表达存在显著性差异,但是关于短期内DlDCL4对外源激素的响应机制还没有相关研究成果。因此,本试验通过比较不同物种DCL4氨基酸序列的保守性,并构建进化树;通过构建表达载体转化洋葱表皮研究龙眼DCL4的亚细胞定位;并通过设置0、4、8、12、24 h的时间梯度,探究龙眼胚性愈伤组织中DlDCL4是否参与对ABA和SA外源激素的应激反应。
1 材料与方法
1.1 试验材料
本试验所用的‘红核子龙眼胚性愈伤组织(EC)来自于福建农林大学园艺植物生物工程研究所。选择固体培养基中生长状态良好的龙眼胚性愈伤组织继代于液体培养基中,并且在2次液体培养基继代以后,选择生长状况最好的愈伤组织,在滤纸上滤干后混样。将0.3 g样品加入分别含有100 μmol/L脱落酸(ABA)和100 μmol/L水杨酸(SA)的MS液体培养基中,置于25 ℃ 120 r/min的摇床,分别处理0、4、8、12、24 h,每个处理做3次生物学重复,收样时将3组生物学重复处理的样品混样后用滤纸吸干水分,液氮急冻处理后保存于?80 ℃冰箱中备用。
1.2 方法
1.2.1 DCL4进化树的构建 使用www.NCBI. com和https://phytozome.jgi.doe.gov/pz/portal.html选择1821种和龙眼DCL4氨基酸序列相似度较高的物种,下载DCL4氨基酸序列,用MEGA.X软件进行氨基酸序列比对和构建,且对构建的进化树进行自检(boot-strap)分析,设置1000次重复检验,其他参数为默认值,并利用在线工具iTOL(https://itol. embl. de/up-load. Cgi)对进化树进行美化。
1.2.2 目的基因序列的获得和扩增 参考陈晓慧[16]的方法获得DlDCL4基因的序列,结合1302载体的酶切位点,设计上下游引物并分别命名为DCL4-1302.F和DCL4-1302.R(表1),进行PCR扩增。将扩增产物进行琼脂糖凝胶电泳验证后用Thermo scientificScientific生物公司的试剂盒对目的条带进行回收,将回收产物和B-zero载体连接后转化DH5-α大肠杆菌,随后将含有重组质粒的大肠杆菌涂板至含有氨苄霉素的LB培养基中过夜培养,挑取阳性克隆进行菌液PCR验证,将验证成功的菌液送至上海博尚生物公司测序。
1.2.3 线性载体的获得 将带有1302质粒的大肠杆菌进行扩繁培养,使用TIANGEN公司的TIANGEN试剂盒提取质粒。利用NcoⅠ以及BcuⅠ2种核苷酸限制性内切酶对上一步骤获得的质粒进行切割得到线性的质粒,并且DCL4基因两端有着可配对序列的线性DNA片段,通过凝胶电泳检测获得的DNA片段是否为目的线性DNA片段。
1.2.4 龙眼DCL4的亚细胞定位 将线性载体和目的基因连接以后将含有重组质粒菌液涂板至含有抗生素的培养基中进行筛选,同样地挑菌后进行菌液PCR,将含有重组质粒的大肠杆菌使用2种内切酶进行酶切验证,验证成功后的大肠杆菌可以用于转化农杆菌。随后将含有重组质粒的农杆菌侵染洋葱内表皮细胞后使用共聚焦显微镜观察。
1.2.5 实时荧光定量PCR 使用Trans geneGene生物公司的TransZol Up試剂盒提取处理后样品的总RNA,然后用TakaraTaKaRa生物公司PrimeScriptTM IV 1st Strand cDNA Syn-thesis Mix试剂盒进行cDNA
合成,具体步骤参考试剂盒说明书。用DNAMAN设计用于qPCR的引物后送至尚亚生物公司合成(表1),以EF-1a为内参基因,以上一步反转录的cDNA为模板进行实时荧光定量PCR,每个模板进行3个重复实验,反应体系为20 μL,反应设置为40个循环。DCL4成员的相对表达量用2?CtCT法计算。
1.3 数据处理
使用Excel软件对实时荧光定量PCR数据进行统计和计算,使用SPSS软件对结果进行差异性显著性分析,并且使用GraphPad Prism 6.01软件绘制统计图。
2 结果与分析
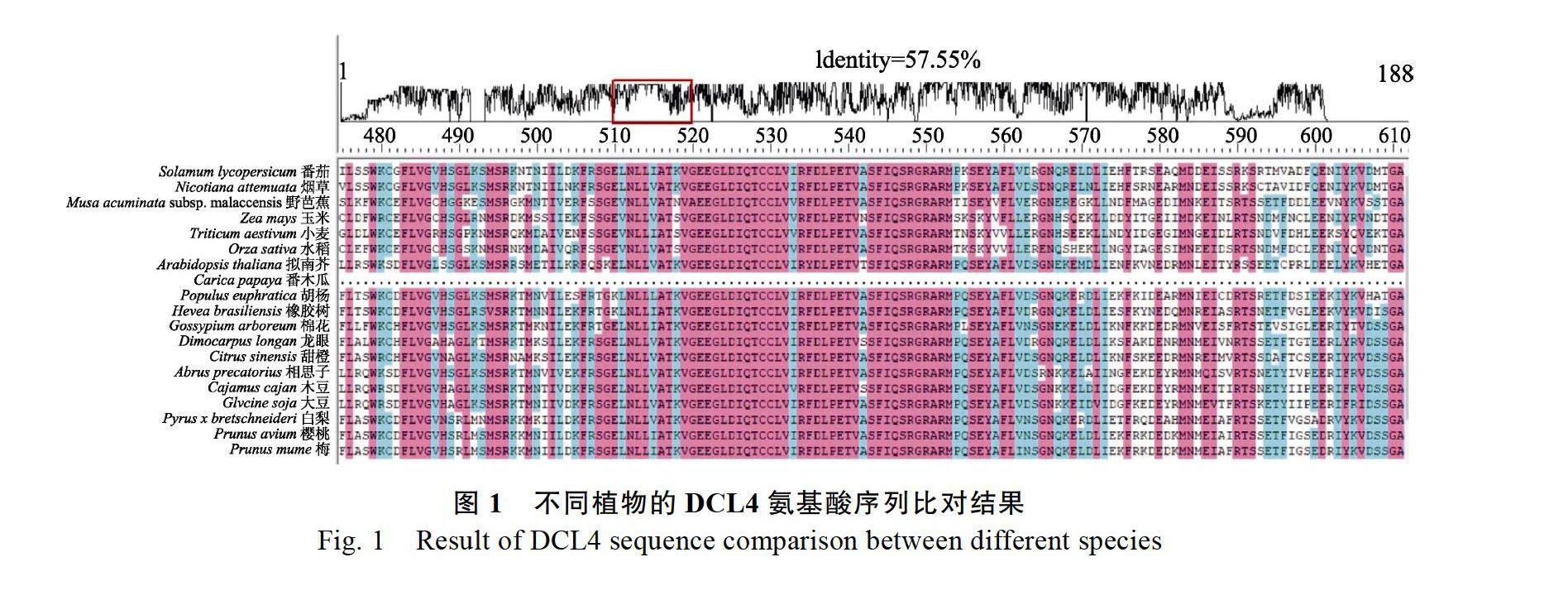
2.1 龙眼DCL4氨基酸序列比对和进化分析
使用MEGA.X软件分析龙眼和其他1820个物种的DCL4蛋白的氨基酸序列比对和进化树分析后发现,DCL4氨基酸序列在物种间比较保守,氨基酸序列的相似度为57.55%(图1);以DCL4的氨基酸序列作为进化依据,柑橘类的甜橙(Citrus sinensis)和龙眼具有最近的亲缘关系(图2)。同时DCL4的氨基酸序列在不同物种间的相似性较高,体现了该基因在不同物种之中相对保守。
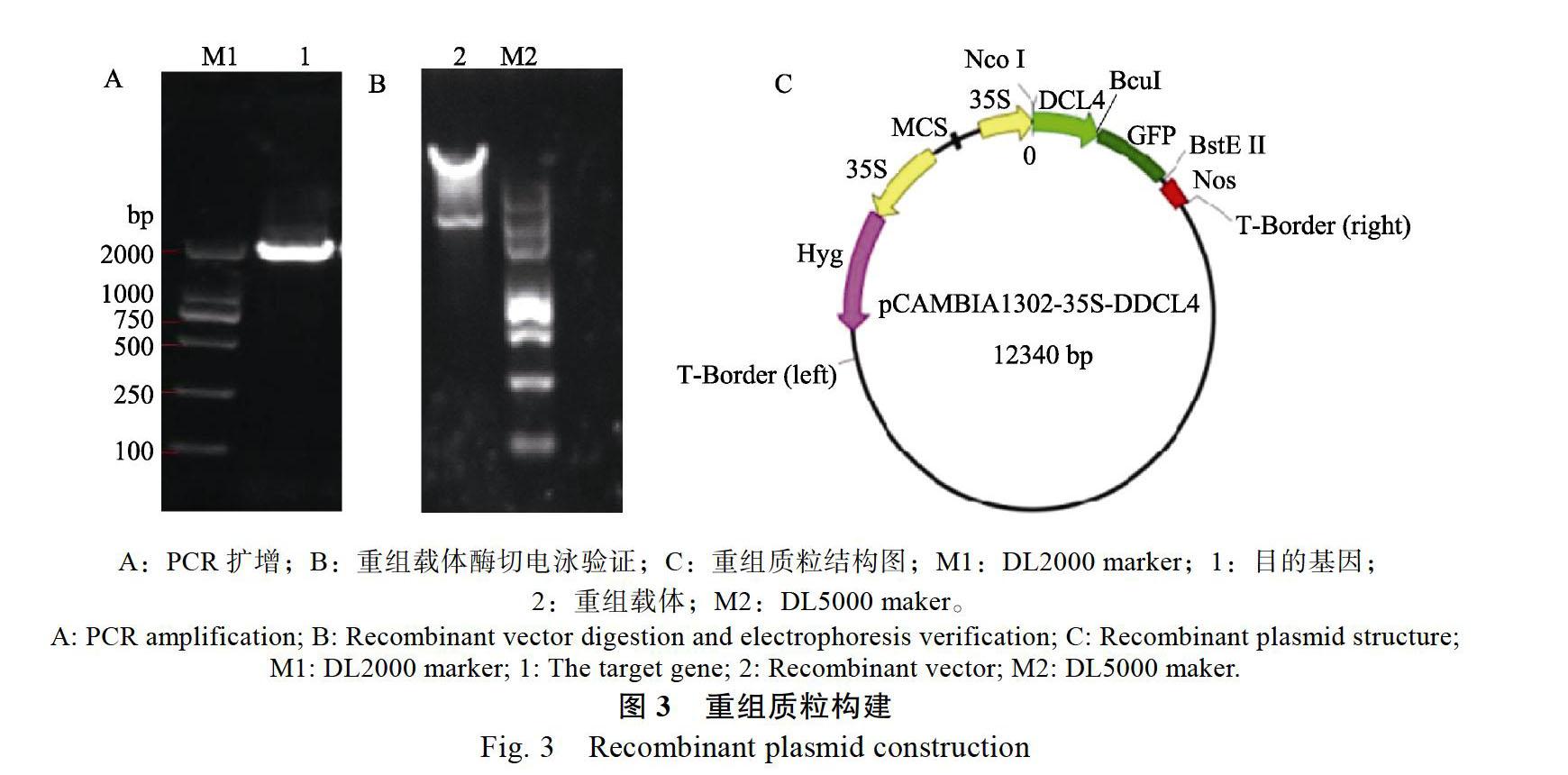
2.2 重组质粒构建
通过PCR扩增获得带有限制性内切酶NcoⅠ和BcuⅠ酶切位点的龙眼DCL4目的片段后(图3A),然后进行菌液验证和送至生物公司测序,将测序成功的DCL4目的片段和酶切获得的线性1302质粒连接,挑取在含有卡纳霉素的LB培养基中的阳性克隆子,再进行酶切验证(图3B),酶切验证成功的载体即为可用于转化至农杆菌中的重组载体,构建完成后的载体可用于亚细胞定位(图3C)。
2.3 DlDCL4亚细胞定位分析
采用农杆菌介导法将DlDCL4:GFP瞬时表达于洋葱鳞叶表皮细胞,用激光扫描共聚焦显微镜观察DlDCL4的亚细胞定位(图4)。实验组的洋葱细胞在暗场下表现为细胞核散发绿色荧光,但是其他位置不发出绿色荧光,空白对照组表现为整个细胞都发出绿色荧光,说明龙眼DlDCL4定位于细胞核中。
2.4 龙眼DlDCL4在外源ABA和SA处理下表达模式
从图5可知,浓度为100 μmol/L外源ABA处理能显著下调处理可显著下调DlDCL4的表达。而在100 μmol/L的外源SA处理下,DlDCL4的表达量明显上调,处理8、24 h时,其表达量显著上调。
3 讨论
DCL属于RNase Ⅲ家族,在进化上高度保守。序列分析表明,各物种的DCL都具有相似的结构域。由于DCL4的功能是剪切dsRNA产生siRNA,所以不难理解其作用于细胞核中[3]。本研究的亚细胞定位结果表明,龙眼DCL4蛋白定位于龙眼细胞核中,本研究结果与高粱[24]以及拟南芥[25]的DCL4亚细胞定位结果相同,推测DCL4在这几个物种间有着相似的功能。
研究表明,在一定浓度范围下,外源SA激素处理可以诱导烟草对某些病毒的获得性抗性[23]。研究发现,经过诱导剂处理后,柑橘对黄龙病的抗性有了极显著的提高,同时SA代谢通路中的关键基因上调表达4~10倍[26]。在龙眼中研究发现,SA处理可以使抗病原相关基因的显著性上调表达[27]。除此之外,在葡萄抗霜霉病[28]、番茄抗叶霉病[29]、苹果抗轮纹病[30]等植物对生物性病原的抗性研究中均有类似的发现。以上研究结果表明,SA信号传导途径在植物響应非生物胁迫过程中发挥了重要作用。同时也有研究发现,SA可以通过上调21 nt的siRNA含量导致拟南芥染色体着丝粒区域的兆碱基规模的甲基化这一途径提高其对植物病原体的抗性[31],DCL4蛋白在大部分生物中的主要功能是特异性切割长链dsRNA产生siRNA作用于相关靶基因,沉默其表达进而调控植株的生长发育和抗性反应[32]。研究发现SA可以沉默马铃薯梅豆病毒的RNA的转录[33]。在本试验中,外源SA激素处理后,龙眼胚性愈伤组织的DCL4显著性上调表达,由此我们推测,外源SA通过诱导DCL4基因的上调表达响应病害胁迫。
在本研究中分别用含有100 μmol/L的ABA和100 μmol/L的SA处理龙眼胚性愈伤组织,结果表现出ABA抑制DlDCL4表达甚至沉默,而SA明显促进DlDCL4表达。已有研究表明,体胚细胞发生过程中,单独加入ABA会抑制其发生[34],在龙眼胚性愈伤组织中的试验也证明了在外源ABA存在的情况下,DlDCL4的表达受到明显的抑制[16],但是本研究对其进行时间梯度处理下DlDCL4的表达量总体没有表现出变化趋势,这可能和ABA处理的浓度有关,外源ABA浓度太高导致时间梯度处理对DlDCL4的表达量不产生影响;在对抗棉花抗枯萎病相关基因WRKY的研究中发现,在使用外源SA处理3 h后WRKY的表达量出现最大值[35],并且推测SA可以刺激棉花在短时间内对枯萎病毒产生应激反应。在本研究中DlDCL4在SA处理8 h时出现最大表达量,和其结果相类似,因此,猜测8 h是DlDCL4响应外源SA刺激时的关键时间。DlDCL4在龙眼体愈伤组织阶段响应激素应答和抵抗非生物胁迫的具体机制还需要进一步深入研究。
参考文献
[30] Margis R, Fusaro A F, Smith N A, et al. The evolution and diversification of Dicers in plants[J]. FEBS Lett, 2006, 580(10): 2442-2450.
[31] Liu Q, Feng Y, Zhu Z. Dicer-like (DCL) proteins in plants[J]. Functional Integrative Genomics, 2009, 9(3): 277-286.
[32] 李 蕊, 王 恬, 文莉薇, 等. 植物Dicer-like功能的研究进展[J]. 中国农学通报, 2015, 31(30): 210-214.
[33] Liu Q P, Feng Y, Zhu Z J, Dicer-like (DCL) proteins in plants[J]. Funct. Iintegr Genomics, 2009, 9(3): 277-286.
[34] Dong Z, Han M H, Fedoroff N. The RNA-binding proteins HYL1 and SE promoteaccurate in vitro processing of pri-miRNA by DCL1[J]. Proceedings of the National Acad-emy of Sciences of the United States of America, 2008, 105(29): 9970-9975.
[35] Gasciolli V, Mallory A C, Bartel D P, et al., Partially redun-dant functions of Arabidopsis DICER-like enzymes and a role for DCL4 in producing trans-acting siRNAs[J]. Current biology, 2005, 15(16): 1494-1500.
[36] Ramachandran V, Chen X. Small RNA metabolism in Arabi?dopsis[J]. Trends in Plant Science, 2008, 13(7): 368-374.
[37] Song X W, Li P C, Zhai J X, et al. Roles of DCL4 and DCL3b in rice phased small RNA biogenesis[J]. The Plant Journal. 2012, 69(3): 462-474.
[38] Wu Y Y, Hou B H, Lee W C, et al. DCL2- and RDR6-dep-endent transitive silencing of SMXL4 and SMXL5 in Arabidopsis dcl4 mutants causesdefective phloem transport and carbohydrate over-accumulation[J]. Plant Journal, 2017, 90(6): 1064-1078.
[39] Moura M O, Fausto A K S, Fanelli A, et al. Genome-wide identification of the Dicer-like family in cotton and analysis of the DCL expression modulation in response to biotic stress in two contrasting commercial cultivars[J]. BMC Plant Biology, 2019, 19(1): 503.
[40] Cui X, Zhang H Y, Chen Z Y. et al.. 2007. Oryza sativa dicer-like4 reveals a key role for small interfering RNA silencing in plant development[J]. The Plant Cell, 2007, 19(9): 2705-2718.
[41] 郭文雅, 趙京献, 郭伟珍. 脱落酸(ABA)生物学作用研究进展[J]. 中国农学通报, 2014, 30(21): 205-210.
[42] 吴 寒. 植物生长发育中脱落酸的生理作用及其在逆境胁迫作用的研究进展[J]. 花卉, 2018(8): 149.
[43] 陈思奕, 王佩茹. 脱落酸对植物耐旱性的影响研究概述[J]. 生物学教学, 2015(5): 2-3.
[44] 白 玉. 龙眼体胚发生过程中DlDRMs的克隆与功能分析[D]. 福州: 福建农林大学, 2018.
[45] 陈晓慧. 龙眼体胚发生过程中DlDCLs的克隆与功能分析[D]. 福建: 福建农林大学, 2018.
[46] 汪 尚, 徐鹭芹, 张亚仙, 等. 水杨酸介导植物抗病的研究进展[J]. 植物生理学报, 2016, 52(5): 581-590.
[47] 郝 转. 水杨酸对盐胁迫黑果枸杞种子萌发的影响[J]. 贵州农业科学, 2019, 47(3): 101-104.
[48] Zhang L, Ma X, Zhou B, et al. EDS1-mediated basal defense and SA- signaling contribute topost-invasion resistance against tobacco powdery mildew in Arabidopsis[J]. Physiological and Molecular Plant Pathology, 2015, 91: 120-130.
[49] Vivancos J, Labbe C, Menzics J G, et al. Silicon-mediated resistance of Arabidopsis againstpowdery mildew involves mechanisms other than the salicylic acid (SA)-dependent defencepathway[J]. Molecular Plant Pathology, 2015, 16(6): 572-582.
[50] Li L, Li T, Jiang G, et al. Synergistion mechanism of exogenous Ca2+ to SA inducedresistance to Botrytis cinerea in tomato[J]. The Journal of Applicd Ecology, 2015, 26(11): 3497-3502.
[51] 陈晓慧, 曾丽兰, 徐小萍, 等. 龙眼Mn-SOD基因的表达及其启动子功能分析[J]. 热带作物学报, 2018, 39(5): 913- 919.
[52] 黎 飞. 植物GST、DCL基因和泛素-蛋白酶体途径对植物抗性的调控功能分析[D]. 杭州: 浙江大学, 2010.
[53] 張 腾, 尹梦娇, 郭红媛, 等. 高粱基因组DCL家族的系统进化与表达分析[J]. 山西农业科学, 2019, 47(6): 950- 956.
[54] Hiraguri A, Itoh R, Kondo N, et al. Specific interactions between Dicer-like proteins and HYL1/DRB-family dsRNA- binding proteins in Arabidopsis thaliana[J]. Plant Molecular Biology, 2005, 57(2): 173-188.
[55] 张 易. 植物免疫诱导剂诱导柑橘对黄龙病抗性的作用机制[D]. 北京: 中国农业科学院, 2018.
[56] 叶 炜. 龙眼胚性培养物抗性相关基因的克隆与功能鉴定[D]. 福州: 福建农林大学, 2015.
[57] 刘少莉. 葡萄抗霜霉病相关基因VaHAESA和VaNPR1筛选及抗性机理研究[D]. 北京: 中国农业大学, 2018.
[58] 薛东齐. 番茄抗叶霉病Cf-12候选基因的筛选及抗性应答机制分析[D]. 哈尔滨: 东北农业大学, 2017.
[59] 侯 珲. 苹果抗轮纹病品种鉴定及SA诱导后抗病差异表达基因的RNA-Seq分析[D]. 兰州: 甘肃农业大学, 2017.
[60] Yoshikawa M. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis[J]. Genes Development, 2005, 19: 2164-2175.
[61] Blevins T, Rajeswaran R, Shivaprasad P V, et al. Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing[J]. Nucleic Acids Research, 2006, 34(21): 6233-6246.
[62] Alamillo J M, Saénz P, García J A. Salicylic acid mediated and RNA-silencing defense mechanisms cooperate in the restriction of systemic spread of plum pox virus in tobacco[J]. Plant Journal, 2006, 48(2): 217-227.
[63] Akula A, Akula C, Bateson M. Betaine a novel candidate for rapid induction of somatic embryogenesis in tea [Camellia sinensis (L.) O. Kuntze][J]. Plant Growth Regulation, 2000, 30(3): 241-246.
[64] 李会会, 赵曾强, 韩泽刚, 等. 棉花GhWRKY106-1的克隆与表达分析[J]. 分子植物育种, 2015, 13(11): 2461-2468.
责任编辑:黄东杰
猜你喜欢
作文大王·低年级(2019年4期)2019-05-13
世界热带农业信息(2018年5期)2018-11-09
世界热带农业信息(2018年4期)2018-10-25
世界热带农业信息(2018年3期)2018-09-26
世界热带农业信息(2018年6期)2018-03-05
世界热带农业信息(2018年7期)2018-01-19
阅读与作文(小学低年级版)(2016年10期)2016-10-12
学苑创造·A版(2016年9期)2016-10-10
小火炬·阅读作文(2016年7期)2016-08-24
饮食科学(2016年7期)2016-07-27
