
外源BR对盐碱胁迫下甜菜内源激素含量及保护酶活性的影响
2021-08-09 08:11陈婧婷刘新宇王玉波李彩凤
西北农林科技大学学报(自然科学版) 2021年7期
王 鑫,刘 丹,陈婧婷,刘新宇,张 贺,王玉波,李彩凤
(东北农业大学 农学院,黑龙江 哈尔滨 150000)
我国是盐碱地大国,也是全球受盐渍化危害最重的国家之一[1],国内盐碱化土壤分布具有明显的地域性,主要集中于干旱和半干早地区[2]。近年来我国土壤盐碱化程度日渐加重[3],耕地中50%以上都有盐碱化的趋势[4],盐渍土面积更是达到了20多万km2,约占国土总面积的2.1%[5],严重威胁着作物的品质和产量。盐碱胁迫抑制了作物对土壤中有效养分的吸收和利用[6],且主要是以渗透胁迫和离子毒害的方式对作物生理过程造成危害[7]。
甜菜(BetavulgarisL.)是我国两大糖料作物之一,经济价值高[8],抗逆性强,是人们所公认的耐盐碱、耐贫瘠、适应性强的作物。甜菜广泛种植于盐碱地,其产糖量约占我国食糖总产量的10%[9],仅次于甘蔗。有研究表明,低浓度Na+可促进甜菜生长,提高其产量和品质;高浓度Na+会使甜菜出苗率降低、植株生长迟缓且矮小、生物量减少、光合效率减弱、产量下降等,严重时甚至导致其死亡[10-12]。
油菜素内酯(BR)是生长调节物质,最先被发现于油菜花粉[13]上,属于甾醇内酯,与动物甾醇类激素类似[14],是公认的第六大植物激素[15]。BR生理活性极强,能增强作物抗逆性和抗氧化能力[16],提高作物品质和产量[17]。同时BR也具有促进作物生理生化代谢、调节酶活性[18]、调控作物生长发育以及激活氧化酶保护系统的功能[19]。近年来关于外源BR的研究多在大豆、玉米等作物上展开,如魏鑫等[20]研究发现,外源BR能够减轻干旱对大豆光合的抑制作用;马野等[21]研究表明,干旱胁迫下BR能提高玉米幼苗的光合效率,维持其正常生长。有关BR在蔬菜方面的研究报道也较多,如有研究表明,BR在蔬菜抗旱、耐低氧、抗盐碱、耐病害等方面有着重要作用[22]。而有关BR对甜菜的生理调控研究较少,例如菅彩媛[23]分析了不同生育时期喷施BR对甜菜生理指标和产量的影响,筛选出了提高甜菜生产性能的BR最佳浓度;刘丹等[24]探究了外源BR促进甜菜生长的最佳质量浓度。本试验通过研究盐碱胁迫下不同时期喷施BR对甜菜内源激素含量和保护酶活性的影响,以探究BR与其他植物激素间的共同调节作用,分析盐碱胁迫下BR对甜菜体内保护酶系统的调控,以期为明确外源BR对甜菜耐盐碱性影响的生理机制提供理论依据,也可为BR在甜菜上的应用提供参考。
1 材料与方法
1.1 试验材料
甜菜品种KWS 0143源自德国艾恩贝克,由卡韦埃斯种子欧洲股份有限公司提供。
1.2 试验设计
试验于2019年在东北农业大学盆栽场进行。供试土壤为东北地区黑钙土(pH=7.65),土壤基本养分含量为:速效磷83.30 mg/kg,速效钾153.20 mg/kg,有机质35.66 g/kg,碱解氮176.27 mg/kg。采用桶栽种植,桶高26.5 cm,桶顶直径30.0 cm,桶底直径24.0 cm。每桶装10 kg风干土壤。土壤中加入盐碱(无结晶水,V(NaCl)∶V(Na2SO4)∶V(NaHCO3)∶V(Na2CO3)=4∶2∶2∶1)使Na+质量分数达到风干土壤的3%,用来模拟大田盐碱土(pH=9.15)。此外土壤中施入基肥:N 180 kg/hm2,P2O590 kg/hm2,K2O 90 kg/hm2。
试验设9个处理:CK(对照,供试土壤与基肥混匀后装于桶中)、YJ(盐碱处理,供试土壤、基肥与盐碱混匀装于桶中),O、S、V(在YJ处理基础上,分别于叶丛形成期(前期)、块根膨大期(中期)和糖分积累期(后期)喷施0.15 mg/kg的BR)以及不同时期喷施BR的组合处理,即OS、SV、OV和OSV。于05-05播种,每个处理30桶。每桶播10粒种子。出苗率达70%左右时间苗,确保喷施BR时每桶只留1株健壮幼苗。
将0.15 mg BR溶于10 mL体积分数90%乙醇中,加入高温灭菌的蒸馏水定容到1 L,滴入1 mL 吐温-80做延展剂,混合均匀,配制成0.15 mg/kg的BR溶液备用。选择晴朗天气,于甜菜生长前期(06-30)、中期(08-06)和后期(09-11)在叶片正反两面均匀喷施BR溶液40 mL,使叶面布满液滴又不流下,保证叶片充分吸收,其他处理喷施等量蒸馏水。
于甜菜生长前期喷施BR后9 d开始采集叶片,每两次取样间隔18 d,共取样5次。选择晴天08:00-10:00进行取样,挑选叶龄一致且完全展开的叶片,取回后洗净,一部分去除叶脉并剪碎混匀,用液氮速冻后于-80 ℃ 冰箱保存;另一部分烘干后测定干质量,用于全氮含量测定。
1.3 测定指标及方法
1.3.1 内源激素含量 采用酶联免疫分析(ELISA)试剂盒(由上海酶联生物科技有限公司提供)。取0.5 g经液氮处理后叶片样品充分研磨,加入PBS,2 000~3 000 r/min离心20 min,收集上清液后分装,留1份待测,其余放4 ℃冰箱备用。首先用酶标仪在450 nm波长下测定吸光度(OD),并通过标准曲线计算甜菜叶片生长素(IAA)、细胞分裂素(CTK)、油菜素内酯(BR)、脱落酸(ABA)、赤霉素(GA)含量,之后计算IAA、CTK、BR、GA与ABA含量的比值,即IAA/ABA、CTK/ABA、BR/ABA、GA/ABA,以分析内源激素比例变化。
1.3.2 保护酶活性 参考王学奎[25]的方法,测定甜菜叶片的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性。
1.3.3 氮素积累量 用凯氏定氮法[26]测定叶片全氮含量,然后计算氮素积累量(g/株):氮素积累量=叶片全氮含量×叶片干质量。
1.3.4 块根产量、含糖率和产糖量 于10-10收获甜菜,每处理取5株,测定块根产量。 用便携式折光仪测定含糖率,并计算产糖量,产糖量=含糖率×块根产量。
1.4 数据分析
采用Excel 2010进行数据处理及制图,利用IBM SPSS Statistics 24进行差异显著性和相关性分析。
2 结果与分析
2.1 外源BR对盐碱胁迫下甜菜叶片内源激素的影响
2.1.1 BR含量 外源BR对甜菜叶片内源BR含量的影响如表1所示。
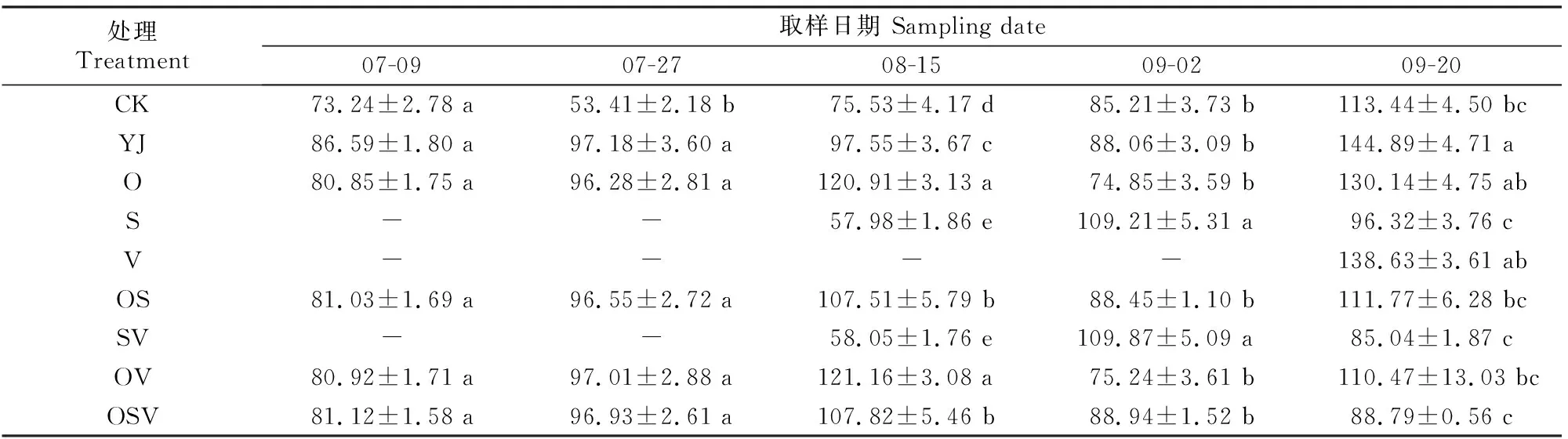
表1 外源BR对甜菜叶片内源BR含量的影响Table 1 Effects of exogenous BR on content of endogenous BR in sugar beet leaves ng/g
如表1所示,各时期YJ处理甜菜的内源BR含量均高于CK处理。07-09和07-27,O、OS、OV和OSV处理的内源BR含量低于YJ处理;08-15,S和SV处理的内源BR含量显著低于其他处理,O、OS、OV和OSV处理的内源BR含量显著高于YJ处理,其中OV处理最高;09-02,S和SV处理的内源BR含量显著高于其他处理,O和OV处理的内源BR含量低于YJ处理;09-20,YJ处理的内源BR含量显著高于除O和V处理之外的其他处理。随着生育期的推进,盐碱胁迫下,无论组合喷施与否,前期喷施BR后甜菜的内源BR含量总体呈先升高后降低再升高的变化趋势,并于生育后期保持在较高水平。
2.1.2 ABA含量 由表2可知,各时期YJ处理甜菜的ABA含量总体高于CK处理。O、OS、OV和OSV处理ABA含量在07-09显著低于CK和YJ处理,07-27和08-15表现为上升趋势,并明显高于YJ处理;S和SV处理的ABA含量在08-15显著低于其他处理(CK除外),之后随时间的推移呈上升趋势;09-20,除O处理外,YJ处理的ABA含量均高于其他处理。与CK相比,盐碱胁迫后甜菜ABA含量总体升高。无论组合喷施与否,前期喷施BR后ABA含量呈先升高后降低再升高的变化趋势,并于后期保持在较高水平,说明ABA与衰老密切相关。
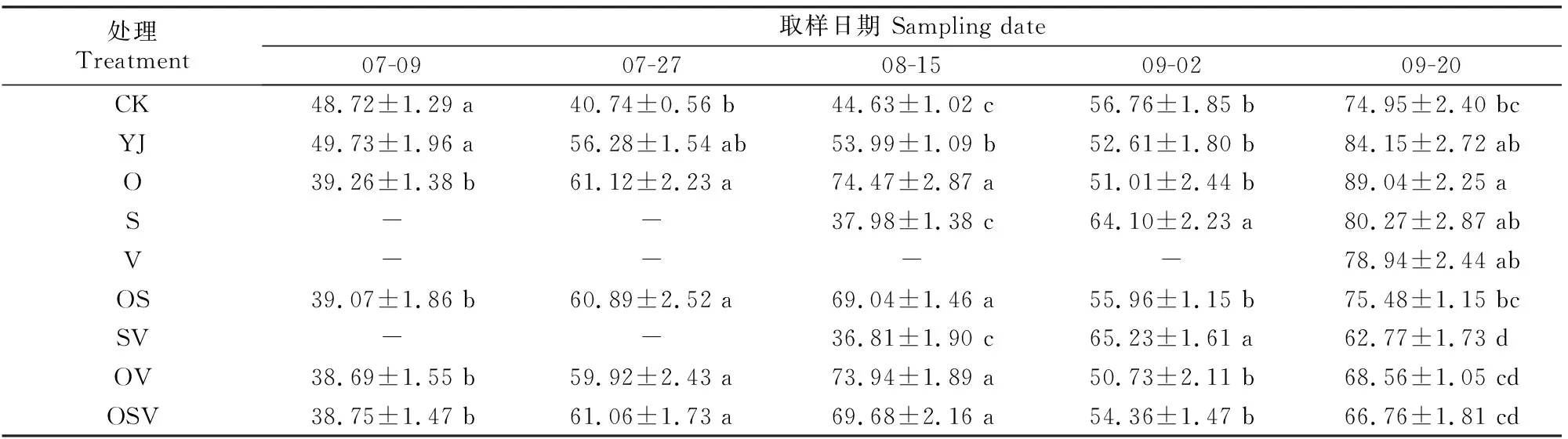
表2 外源BR对甜菜叶片内源ABA含量的影响Table 2 Effects of exogenous BR on content of endogenous ABA in sugar beet leaves ng/g
2.1.3 IAA含量 IAA可促进作物生长,主要参与细胞壁合成和核酸代谢。如表3所示,随着甜菜生育进程的推进,YJ处理的IAA含量逐渐高于CK处理,但其植株却生长缓慢,可能是因为部分IAA参与了细胞壁的重新合成。O、OS、OV和OSV处理的IAA含量在07-09显著低于CK和YJ处理,07-27和08-15表现为上升趋势,并显著高于CK和YJ处理。中后期甜菜老叶片脱落,新叶片生成,促使IAA含量升高,但09-02 S、SV、OS和OSV处理的IAA含量显著低于CK、YJ、O和OV处理,可能是因为中期喷施BR使老叶衰老减慢,新叶生成较少。09-20,CK处理的IAA含量最低,YJ处理的IAA含量低于除CK和O处理之外的其余处理。综上可知,盐碱胁迫下,喷施BR后生长后期甜菜叶片IAA含量整体呈升高的趋势。

表3 外源BR对甜菜叶片内源IAA含量的影响Table 3 Effects of exogenous BR on content of endogenous IAA in sugar beet leaves ng/g
2.1.4 GA含量 由表4可知,随着甜菜生育进程的推进,CK处理的GA含量先下降后升高;YJ处理的GA含量总体高于CK处理。O处理的GA含量与YJ处理无显著差异。08-15,S和SV处理的GA含量均较低;09-02,OS和OSV处理的GA含量均较高;09-20,O、S和V 处理与YJ 处理无显著差异,且高于其他处理,OS、OV、SV和OSV处理的GA含量显著低于O、S和V处理。随着生育期的推进,盐碱胁迫下,无论组合喷施与否,前期喷施BR后甜菜的GA含量呈先升高后降低再升高的变化趋势,且不同时期组合喷施BR对甜菜叶片GA含量影响较大。

表4 外源BR对甜菜叶片内源GA含量的影响Table 4 Effects of exogenous BR on content of endogenous GA in sugar beet leaves ng/g
2.1.5 CTK含量 如表5所示,随着生育期的推进,CK处理甜菜的CTK含量先下降后升高;YJ处理的CTK含量总体低于CK处理。07-09和07-27,除CK处理外,其余处理的CTK含量无显著差异;08-15,喷施BR处理的CTK含量显著高于CK和YJ处理;09-02,O和OV处理的CTK含量显著高于其他处理;09-20,除O处理之外,YJ处理的CTK含量均低于其他处理。综上可知,盐碱胁迫下,喷施BR总体上提高了甜菜生长后期的CTK含量。

表5 外源BR对甜菜叶片内源CTK含量的影响Table 5 Effects of exogenous BR on content of endogenous CTK in sugar beet leaves ng/g
2.1.6 内源激素含量的相关性 如表6所示,盐碱胁迫下甜菜内源激素含量间均呈正相关关系。内源激素BR、GA和ABA含量间相关性达到极显著水平,表明这3种内源激素在甜菜生长发育中起协同作用。内源激素CTK与IAA间相关性达显著水平,证明二者可共同促进甜菜的生长发育。

表6 盐碱胁迫下甜菜叶片内源激素含量的相关性Table 6 Correlation analysis of endogenous hormone contents in sugar beet leaves under saline-alkali stress
2.1.7 内源激素比例 由图1~图4可知,07-09,喷施BR处理的IAA/ABA、GA/ABA、BR/ABA和CTK/ABA值均显著高于YJ处理。07-27,喷施BR处理的IAA/ABA值显著高于YJ处理,GA/ABA、BR/ABA和CTK/ABA均低于YJ处理。08-15,除O和OV处理之外,其余处理IAA/ABA值均显著高于YJ处理;喷施BR处理的GA/ABA和BR/ABA值均显著低于YJ处理,而CTK/ABA值均显著高于YJ处理。09-02,OS和OSV处理的IAA/ABA和BR/ABA值显著低于其他处理;SV和V处理的GA/ABA和CTK/ABA值显著低于其他处理。09-20,除O处理外,YJ处理的IAA/ABA和CTK/ABA值显著低于其他处理;V处理的IAA/ABA、GA/ABA、BR/ABA和CTK/ABA值均高于YJ处理。综上可知,盐碱胁迫下,喷施BR后GA/ABA和BR/ABA值无明显变化规律;随着生育期的推进,无论组合喷施与否,中期喷施BR后甜菜的IAA/ABA值总体呈先下降后升高的趋势;前期喷施BR后甜菜的CTK/ABA值呈先下降后升高再下降的变化趋势。
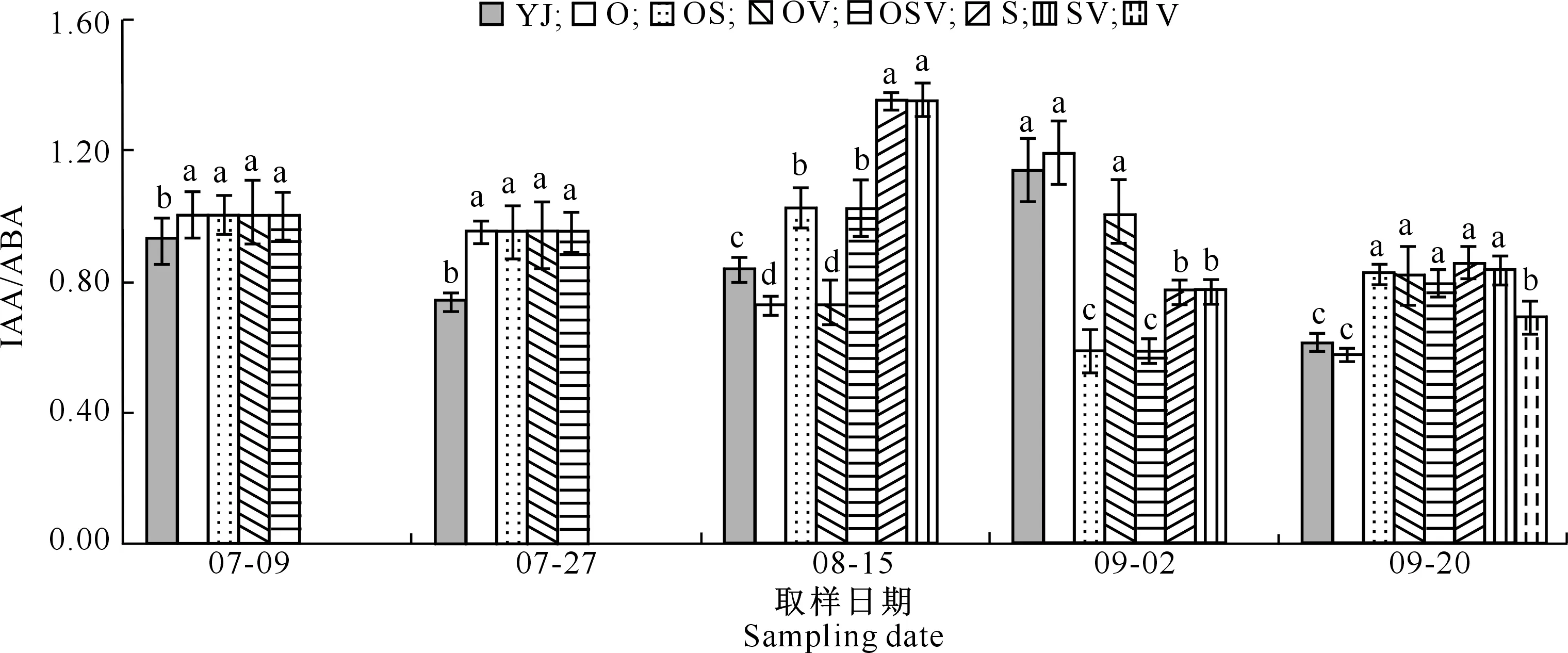
图柱上标不同小写字母表示同一日期下各处理间差异显著(P<0.05)。下图同Different lowercase letters indicate significant differences at P<0.05 level between treatments.The same below图1 盐碱胁迫下外源BR对甜菜叶片IAA/ABA的影响Fig.1 Effects of exogenous BR on IAA/ABA in sugar beet leaves under saline-alkali stress

图2 盐碱胁迫下外源BR对甜菜叶片GA/ABA的影响Fig.2 Effects of exogenous BR on GA/ABA in sugar beet leaves under saline-alkali stress

图3 盐碱胁迫下外源BR对甜菜叶片BR/ABA的影响Fig.3 Effects of exogenous BR on BR/ABA in sugar beet leaves under saline-alkali stress
2.2 外源BR对盐碱胁迫下甜菜叶片氮素积累量的影响
由图5可知,在整个生育期中,CK处理甜菜的氮素积累量先下降后升高,YJ处理的氮素积累量均显著低于其他处理,OSV、S和SV处理的氮素积累量先升高后下降,O、OS和OV处理则呈先上升后下降再上升的趋势。09-20,O和V处理的氮素积累量显著高于其他处理。综上可知,盐碱胁迫下,喷施外源BR总体上增加了甜菜叶片的氮素积累量,以中后期表现最为明显。

图5 盐碱胁迫下外源BR对甜菜叶片氮素积累量的影响Fig.5 Effects of exogenous BR on nitrogen accumulation in sugar beet leaves under saline-alkali stress
2.3 外源BR对盐碱胁迫下甜菜叶片保护酶活性的影响
2.3.1 SOD活性 如表7所示,各处理甜菜的SOD活性均表现为前期低而后期高。07-09和07-27,各处理的SOD活性基本相当;YJ处理的SOD活性在08-15最高,09-02和09-20显著低于其他处理。综上可知,外源BR可以明显提高生长中后期甜菜叶片SOD活性。
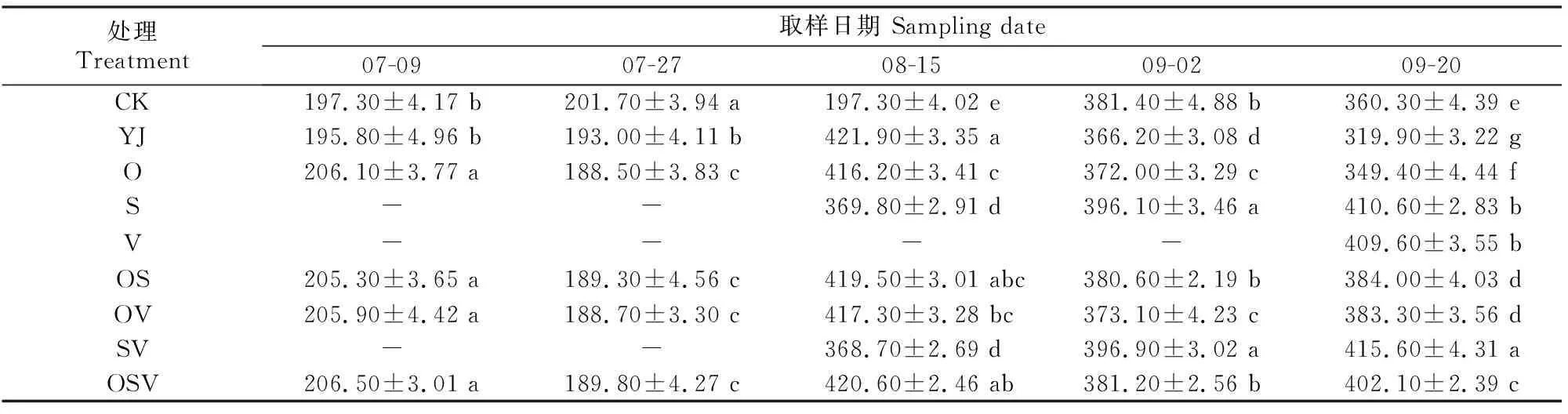
表7 外源BR对甜菜叶片超氧化物歧化酶(SOD)活性的影响Table 7 Effect of exogenous BR on SOD activity in sugar beet leaves U/g
2.3.2 POD活性 由表8可知,各时期CK处理甜菜的POD活性均高于YJ处理。O、OS、OV和OSV处理的POD活性在07-09显著低于YJ处理,07-27和08-15表现为上升趋势,并显著高于CK和YJ处理。08-15,S和SV处理的POD活性与YJ处理无显著差异;09-02,CK和YJ处理的POD活性显著高于其他处理;09-20,V处理的POD活性与YJ处理无显著差异。综上可知,盐碱胁迫下,喷施BR后甜菜的POD活性整体呈先上升后下降的变化趋势,且越早喷施BR,对甜菜叶片POD活性的影响越大。

表8 外源BR对甜菜叶片过氧化物酶(POD)活性的影响Table 8 Effect of exogenous BR on POD activity in sugar beet leaves U/(min·g)
2.3.3 CAT活性 如表9所示,各时期YJ处理甜菜的CAT活性基本高于CK处理。
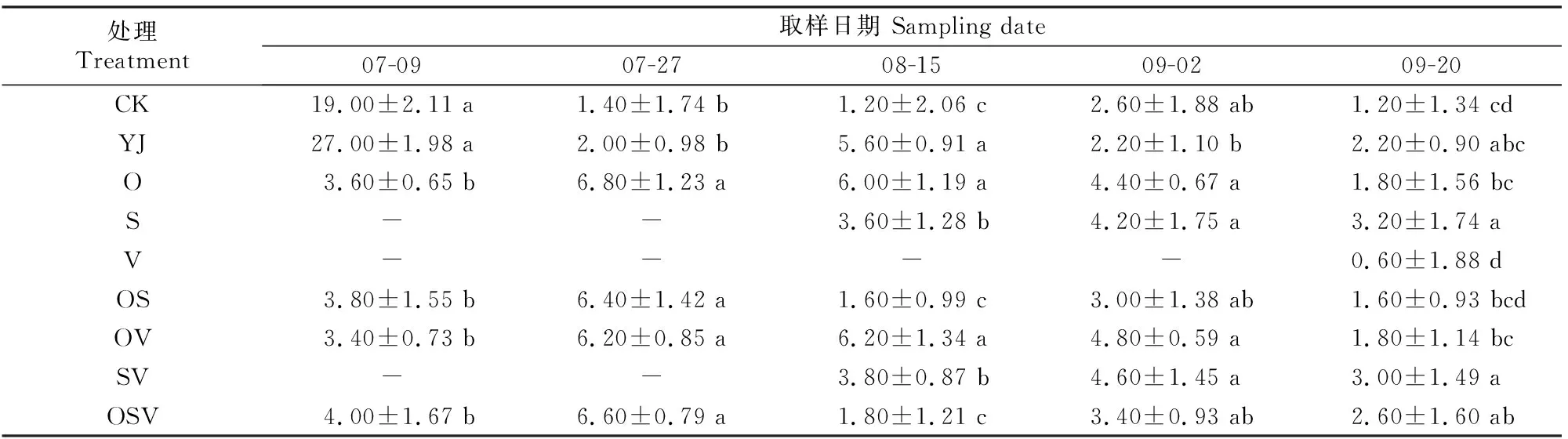
表9 外源BR对甜菜叶片过氧化氢酶(CAT)活性的影响Table 9 Effect of exogenous BR on CAT activity in sugar beet leaves U/(min·g)
O和OV处理的CAT活性在07-09显著低于YJ处理,在07-27-09-02均高于YJ处理。08-15,S、SV、OS和OSV处理的CAT活性显著低于YJ处理;09-02,YJ处理的CAT活性最低;09-20,V处理的CAT活性显著低于除CK和OS之外的其他处理。随着生育期的推进,盐碱胁迫下,喷施BR后甜菜的CAT活性整体呈先上升后下降的变化趋势(表9)。
2.4 外源BR对甜菜块根产量、含糖率和产糖量的影响
外源BR对甜菜块根产量、含糖率和产糖量的影响如表10所示。
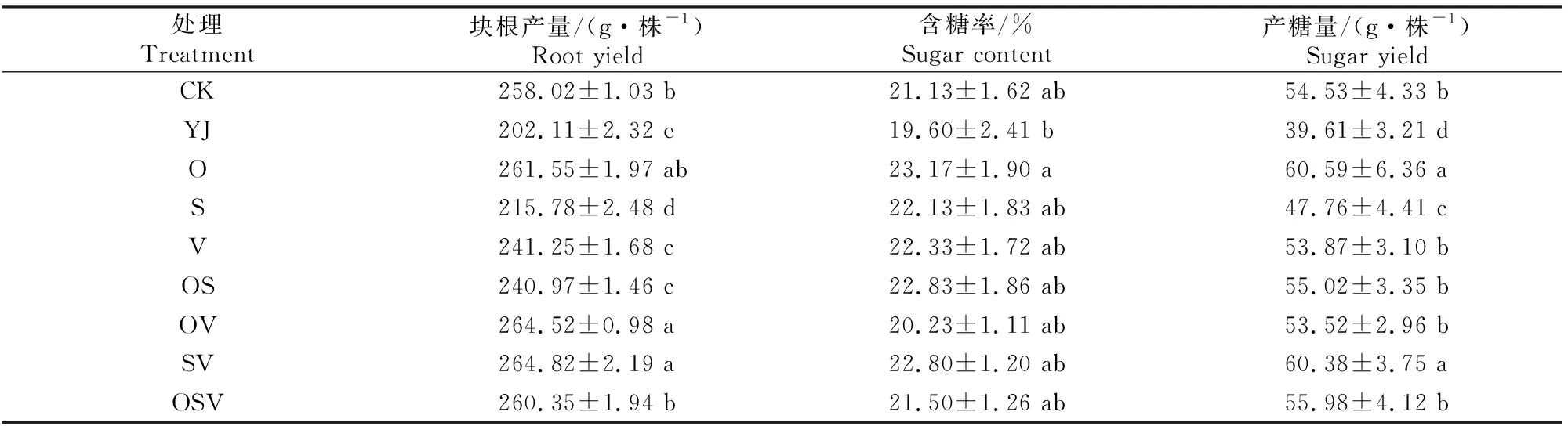
表10 外源BR对甜菜块根产量、含糖率和产糖量的影响Table 10 Effects of exogenous BR on yield,sugar content and sugar yield of beetroot
如表10所示,YJ处理的甜菜块根产量和产糖量显著低于其他处理,且含糖率最低。O、SV和OSV处理的块根产量、含糖率和产糖量高于CK处理。O和SV处理的产糖量显著高于其他处理;除YJ和O处理外,其余处理的含糖率无显著差异。综上可知,盐碱胁迫下,喷施BR有效提高了甜菜的块根产量、含糖率和产糖量;前期喷施BR甜菜的含糖率和产糖量均最大,块根产量也较高,因此为最佳选择。
3 讨 论
内源激素是信号传导物质,具有复杂多样的生理机制。ABA被认为是逆境激素,在作物对逆境的适应中起主要调控作用。GA调控衰老,参与作物对逆境胁迫的响应。CTK延缓衰老,提高作物抗逆性。此外,BR也是公认的逆境激素,生理活性强,本试验发现,喷施BR有效加强了甜菜耐盐碱和抗氧化的能力,与吴雪霞等[27]研究结论一致。Cohen等[28]研究表明,逆境条件下植物内源ABA含量明显上升。魏湜等[29]研究发现,盐碱胁迫使玉米幼苗ABA含量处于较高水平,喷施BR后ABA含量下降。李宁等[30]研究表明,弱光胁迫下喷施BR后番茄叶片IAA和GA含量上升,ABA含量显著下降。Pal等[31]研究表明,外源BR提高了萝卜IAA含量,降低了ABA含量。赵雪松等[32]研究表明,外源BR提高了水稻根系的CTK和GA含量。本试验结果显示,盐碱胁迫下,甜菜叶片ABA含量总体升高,外源BR提高了甜菜生长后期的CTK含量;不同时期喷施BR后,IAA含量整体呈升高的趋势,与上述研究结果一致。本研究发现,随着生育期的推进,盐碱胁迫下,无论组合喷施与否,前期喷施BR后甜菜的内源BR、ABA和GA含量总体呈先升高后降低再升高的变化趋势。
内源激素通过交互作用调控作物生育进程。相较于单一内源激素,不同激素间的平衡变化更能反映作物生育的本质规律。王廷芹等[33]发现,外源BR显著影响青花菜茎尖内源激素的平衡。本研究发现,盐碱胁迫下外源BR对甜菜GA/ABA、BR/ABA、IAA/ABA和CTK/ABA影响较大。
前人研究表明,盐碱胁迫使甜菜前期、中期生长受到抑制,吸收利用少量土壤氮素,因此在中期、后期甜菜老叶片无缺氮素症状,而未受胁迫的甜菜前期、中期生长快速,很快耗尽土壤氮素,并在中期、后期表现为老叶片枯死,新生叶片呈淡绿色[34]。李勇[35]研究表明,外源BR可以用于调控小麦的氮素吸收积累。本研究发现,盐碱胁迫下,喷施外源BR总体上提高了甜菜生长中期、后期叶片的氮素积累量。
SOD、POD和CAT是作物内在的保护酶,通过相互间的分工合作,对作物抗逆代谢起着重要的调控作用。SOD是作物抵御活性氧伤害的“第一道防线”,可将活性氧歧化成过氧化氢,过氧化氢的累积又会造成膜脂过氧化,导致细胞膜损伤,而POD与CAT共同作用,清除由SOD作用产生的过量过氧化氢。王建华等[36]研究表明,外源BR通过提高SOD活性来抑制膜脂质过氧化。前人研究发现,在逆境条件下,喷施BR后植物叶片中的SOD、POD和CAT活性升高[37-38]。本试验结果表明,盐碱胁迫下喷施BR可以明显提高甜菜生长中后期叶片SOD活性,与上述结果一致。王秀峰等[39]研究表明,盐碱胁迫下,喷施BR使辣椒叶片SOD、POD、CAT活性均呈先上升后降的趋势。本试验中,盐碱胁迫下,不同时期喷施BR后甜菜叶片的POD和CAT活性整体呈先上升后下降的变化趋势,但甜菜整个生长期的POD活性均受盐碱胁迫抑制,这可能是因为POD更多地参与了甜菜的其他生理调控过程。
BR能提高作物中磷酸蔗糖合成酶的活性,增加蔗糖积累量,进而提高作物产量[40]。谢云灿等[41]发现,高温胁迫下,外源BR提高了大豆籽粒可溶性总糖含量,增加了大豆产量。唐鑫华等[42]研究显示,喷施BR增加了马铃薯块茎的淀粉含量,提高了单株产量。此外,BR同样能提高水稻[43]、玉米[44]、小麦[45]等作物的产量及可溶性糖、蔗糖和淀粉含量。本研究发现,盐碱胁迫下,喷施外源BR提高了甜菜的块根产量、含糖率和产糖量。
4 结 论
外源BR可明显提高甜菜叶片的保护酶活性,增加氮素积累量,促进内源激素生成,调节内源激素的比例,提升甜菜耐盐碱和抗氧化的能力,提高块根产量、含糖率和产糖量。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年9期)2022-07-18
作物学报(2022年8期)2022-05-29
中国糖料(2022年2期)2022-04-06
金桥(2021年11期)2021-11-20
小哥白尼(趣味科学)(2021年12期)2021-03-16
悦游 Condé Nast Traveler(2021年4期)2021-01-13
东坡赤壁诗词(2018年2期)2018-05-10
环球时报(2017-09-19)2017-09-19
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
