
麻疯树中一种II型核糖体失活蛋白基因的克隆和表达调控分析
2021-09-18 10:07李笑王菲彭婕陈放徐莺
农村实用技术 2021年8期
李 笑 王 菲 彭 婕 陈 放 徐 莺
(四川大学生命科学学院 生物资源与生态环境教育部重点实验室,四川 成都 610065)
1 引言
麻疯树是一种多用途能源植物,具有适应性强且种子含油量高的特点,同时含有多种活性成分,如佛波酯、麻疯树毒蛋白、凝集素等[1]。麻疯树毒蛋白及一些凝集素都属于核糖体失活蛋白(ribosome-inactivating protein, RIP)。
RIP是一类能破坏真核细胞核糖体结构的毒蛋白,在植物应对生物和非生物胁迫时承担调节作用[2]。根据基因的序列结构和性质,RIP家族可分为3类[3]:I型RIP数量最多,含有单个RIP结构域(PF00161);II型RIP毒性较强,除RIP结构域外,其C端还含有凝集素结构域(PF00652),分别称为A链和B链,由二硫键连接;III型RIP结构与I型相比具有一个额外的20kDa的C端区域,前体多肽需经翻译后加工步骤去除N端、C端的短片段及中间连接序列后才具有活性,目前仅发现来自大麦的JIP60和来自玉米的b-32。
大戟科的沙匣树、商陆科美洲商陆衰老和受胁迫的叶片中都提取出了核糖体失活蛋白,表明这类蛋白可能在胁迫响应中发挥功能[4]。若能从麻疯树中分离出高毒性的凝集素,将进一步提高麻疯树的应用价值。因此,本研究对麻疯树中II型RIP基因编码的序列同源性、重要氨基酸位点进行分析,并对其时空表达模式进行探究,以期为高抗性、高产量麻疯树的育种提供理论基础。
2 材料与方法
2.1 植物材料及处理
麻疯树种子购买自四川攀枝花,在种子萌发后两周左右,待第一片真叶长约0.5~1cm,进行组织取材,包括麻疯树的胚乳、子叶、第一片真叶、叶柄、茎、侧根、主根等组织。在对麻疯树进行干旱处理,分别在处理的第1、2、4和7天四个时间阶段收集根和叶组织,同时将未处理的组织作为对照组,迅速放入液氮中,置于-80℃冰箱中保存。
2.2 生物信息学分析
从NCBI(https://www.ncbi.nlm.nih.gov/)上下载麻疯树的基因组数据,从Pfam数据库(http://pfam.xfam.org/)中下载RIP结构域模型(PF00161)和Ricin_B_lectin结构域模型(PF00652),利用本地HMMER程序对麻疯树的基因组进行检索,将 E值小于1e-5的作为潜在的候选RIP序列。用ExPASy(htp://au.expasy.org)的 ComputepI /Mw计算蛋白质的等电点、分子量。用BLAST进行同源性对比,用MEGAX构建系统进化树。氨基酸序列比对在Clustal Omega中完成。蛋白三维结构的预测在SWISS-MODEL(https://swissmodel.expasy.org/)完成。
2.3 引物设计
利用NCBI中的primer-blast工具,根据序列设计扩增产物和定量引物(表1)。
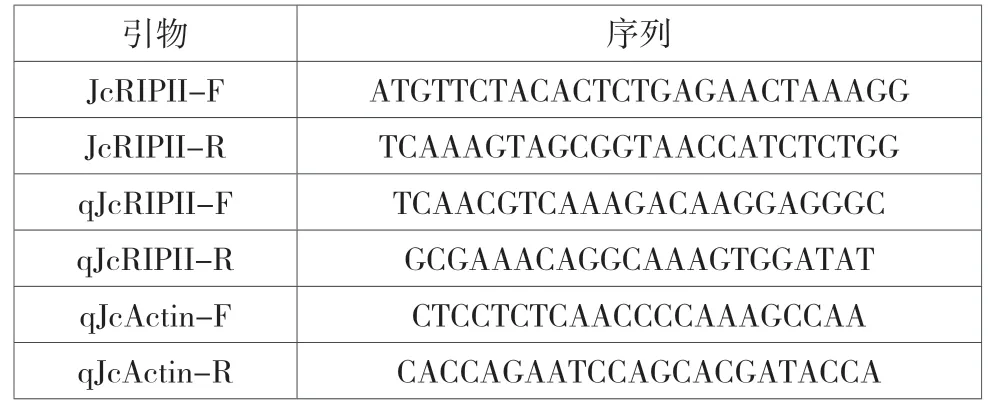
表1 所用引物序列
2.4 RNA提取及cDNA合成
根据福际生物生产的植物基因组DNA提取试剂盒和植物总RNA提取试剂盒说明书的要求分别提取DNA和RNA,随后立即用Takara生产的反转录试剂盒根据说明书对总RNA进行反转录,合成的cDNA保存于-20℃冰箱。
2.5 PCR扩增与表达分析
分别以DNA和cDNA为模板,利用JcRIPII-F和JcRIPII-R扩增基因序列。将与预期大小一致的扩增产物送至生工生物公司测序。
为保证荧光定量引物的特异性,以DNA为模板进行PCR扩增后验证。以麻疯树Actin基因作为内参基因,以保存的不同组织及温度处理后的cDNA为模板,在荧光定量PCR仪进行实时荧光分析。按照AceQ qPCR SYBR Green Master Mix(Vazyme)说明书进行RT-qPCR,相对表达量利用2-ΔΔCT方法进行分析。对于每种处理均使用三个技术重复和至少三个独立的生物学重复。
3 结果与分析
3.1 JcRIPII基因克隆及氨基酸序列分析
在麻疯树基因组中找到一个可能编码的II型RIP蛋白的基因LOC105637075,经可变剪切有两种可能的蛋白编码产物,分别为NP_001295735.1和XP_020536135.1(图1A),后者比前者多出了10个氨基酸,可能是因为前者的第1位和第11位都为甲硫氨酸,是可能的起码密码子,因此拼接时无法判断哪个是真正的翻译起始位置,于是将较长的蛋白做为主要研究对象,将较短的作为其亚型。
以麻疯树DNA和cDNA为模板,均能扩增出长度为1800bp左右的片段,并且测序结果片段的长度和序列一致,表明该基因的CDS区不具有内含子。根据这一基因所在物种、编码蛋白的名称和类型将其命名为JcRIPII(图1B),其对应的理论蛋白含561个氨基酸,分子量为6.21kDa,等电点为6.18,为弱酸性蛋白。
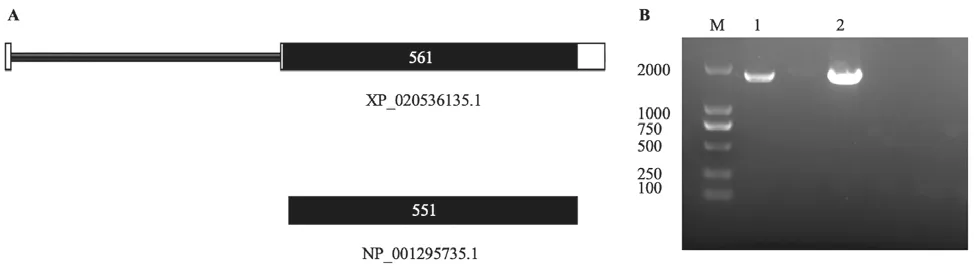
图1 麻疯树JcRIPII基因结构及扩增产物
3.2 JcRIPII基因编码产物结构域分析
结构域预测结果显示,JcRIPII基因编码蛋白序列的N端54~247位氨基酸为A链RIP结构域,C端有两个Ricin_B_lectin结构域(以下简称RBL结构域),分别位于309~429位和441~557位。利用SWISS-MODEL构建蛋白结构模型时比对到一致度和覆盖长度最完整的模板是樟科的cinnamomin III,故将其作为模板预测JcRIPII的结构(图2A)。N端未预测出结构的区域前43个氨基酸可能是信号肽区域。Cinnamomin III的A、B两条链由Cys250-Cys289这两个半胱氨酸形成的二硫键连接[5],序列比对显示在JcRIPII的相似位置处存在Cys288和Cys300,可能是它的A、B链间二硫键相连的位置,因此连接肽的位置应位于这两个半胱氨酸之间。
JcRIPIIA链的6个β-折叠和2个α-螺旋的位置与cinnamomin III相似。RIP结构域中通常含有5个对于N-糖苷酶活性发挥至关重要的残基位点,在JcRIPII中对应于ricin极性亲水的氨基酸Tyr80的Tyr117被替换成非极性氨基酸Gly117(图2B),其他活性位点均保守。在ricin中Tyr80的羟基被认为可与Gly121的羰基氧原子形成氢键,从而旋转形成容纳腺嘌呤的空间,因此JcRIPII这一位点的改变可能会降低JcRIPII的N-糖苷酶活性[6,7]。
JcRIPII的B链中由两个同源的球状结构域组成,每个结构域包括α、β和γ三个亚基,与ricin B链的结构非常相似。首先,在JcRIPII的B链含有9个半胱氨酸,除Cys491外的8个半胱氨酸与ricin的分布十分相似。在ricin中,这些半胱氨酸被认为形成4个二硫键,起着稳定结构的作用[8]。同ricin一样,JcRIPII B链的疏水核心的氨基酸高度保守[9](图2C),每个亚基中都含有QxW重复基序,只在JcRIPII中1γ亚基中表现为QxF[10]。不过个别氨基酸的变异值得注意,如与ricin相比1α中的Ile371被Leu354替代,1γ的Trp407被Phe429替代,2γ的Leu547被Met532替代,其中后两处是不同性质氨基酸的替代,这些变化可能对凝集素的活性造成影响。Ricin和cinnamomin属于半乳糖型凝集素,这些II型RIP B链的六个亚基中仅有两个亚基(1α和2γ)显示出凝集素活性,都是依靠五个氨基酸发挥半乳糖结合作用[11]。在JcRIPII的B链1α和2γ的半乳糖结合位点与ricin相比存在两处氨基酸的替换,其中2γ中Trp547取代了Tyr562,1α中His332取代了Gln349。在前一位置两者都是芳香族氨基酸,变可能不会造成结合活性方面根本性的改变。但是,后一位置的变化引入了正电荷,其对凝集素活性的可能影响值得关注。
3.3 JcRIPII基因编码产物同源性分析
利用BLAST在线工具查找与JcRIPII同源性较高的蛋白序列,结果显示它与棕榈科植物油棕、椰子以及山茶科的茶中的同源物具有50%以上的一致性(51.05%~54.8%)。其次是来自樟科的香樟和沉水樟的多条RIPs序列,其中有代表性的是已获得晶体结构的cinnamomin III。与同为大戟科的蓖麻中的II型RIP一致度最高为47.19%。选取与麻疯树一致度最高的序列构建NJ树(图3A),系统发育树的结果与序列一致度的结果相似,未表现出明显的单双子叶或同一科聚类偏好的现象。大戟科植物油桐、橡胶树、蓖麻与禾本科植物疣粒稻中的II型RIP形成一个进化树分枝,而与JcRIPII关系较远。
为探究RIP中典型RIP结构域的进化历程,我们利用JcRIPII和II型RIP及两条来自麻疯树I型RIP的RIP结构域部分建构了系统发育树(图3B)。结果显示,仅从RIP结构域序列来看,JcRIPII与同科植物II型RIP的进化关系比与同种植物I型RIP的进化关系更近。具体来说,麻疯树的JcRIPII和同科植物油桐及橡胶树中的II型RIP的RIP结构域序列聚类于相邻的进化分枝上,但是麻疯树中的I型和II型RIP含有的RIP结构域序列在进化树上距离相对较远。值得注意的是,JcRIPII并不是与所有同科的II型RIP都紧密相邻,它与来自蓖麻的II型RIP在进化树上距离就较远。同时,RIP结构域的NJ树中依然体现了JcRIPII与来自油棕、椰子和茶的RIP结构域较近的进化关系,它们与JcRIPII的距离仅次于两条同科植物的II型RIP。
同时为研究II型B链之间的进化关系,利用B链中保守的RBL结构域区序列构建了NJ树(图3C)。此时来自麻疯树的JcRIPII和同科植物蓖麻的II型RIP表现出较近的进化关系,与油桐和橡胶树II型RIP形成的分枝关系较远。
3.4 JcRIPII上游调控元件预测分析
顺式作用元件预测结果显示JcRIPII起始密码子上游的启动子区中含有多种胁迫响应和光调节有关的元件(表2)。胁迫响应元件包括在植物创伤诱导元件,此外还含有干旱、厌氧、创伤诱导元件,表明JcRIPII可能在逆境调控中发挥作用。但是没有发现与温度胁迫相关的响应元件,如HSE和LTRE等。

表2 JcRIPII上游顺式作用元件预测分析
3.5 JcRIPII的表达模式初步研究
利用实时荧光定量PCR技术对JcRIPII在麻疯树各组织中的表达情况进行分析(图4A)。发现除幼嫩的第一片真叶外,其他组织器官中JcRIPII基因均有表达,其中在成熟种子胚乳中表达量最高,然后依次是侧根、子叶、主根、叶柄、茎,在幼嫩的真叶中未检测到其转录产物。

图4 JcRIPII基因表达
在干旱胁迫下,麻疯树叶和根中JcRIPII基因表达趋势总体相似,即随着胁迫的进行JcRIPII基因表达量逐渐下降,在第2天下降到最低,随后逐渐上调。但是JcRIPII基因在两个组织的表达也存在一定的差异。首先,在前2天JcRIPII基因表达量下调更显著。同时,叶中的JcRIPII基因在第7天时表达量相对于第4天上调,恢复到处理前的水平,但是根在第7天时表达量相对于第4天下调,仅为对照组中的0.4倍。
4 结语
研究表明毒麻疯树中存在凝集素[12]。同源比对显示,JcRIPII的B链为RBL结构域,也属于凝集素家族,但是关于麻疯树中凝集素蛋白的核酸及序列信息尚未见报道。在本研究中,我们首次对麻疯树基因组中鉴定到的一个II型核糖体失活蛋白基因JcRIPII,基于现有的生物信息学工具对其可能编码的蛋白序列和结构进行了分析,并对其表达模式进行了研究。
JcRIPII蛋白与ricin等II型RIP在三维结构结构和N-糖苷酶活性位点存在相似性,差异主要体现在数个重要氨基酸位点的改变。如A链中对于N-糖苷酶活性至关重要的Tyr117被非极性氨基酸Gly117替换,这一替换对蛋白活性的影响是不确定的。例如将SLT-IA中的Tyr77突变为Ser后活性降低1000倍,突变为Phe后活性降低15倍,但是ricin中Tyr80突变为Phe其活性不发生改变[13,14]。B链中1α和2γ这两个发挥凝集素活性的亚基中也有氨基酸的替换,在cinnamomin的2γ亚基中也存在一个相同的变化(His575替代了ricin中的Gln570),并且相关研究表明这是其活性较低ricin低的重要原因[15,16],因此这一变化可能也会降低JcRIPII的毒性。
为探究JcRIPII基因的进化方式,利用完整的II型RIP蛋白序列构建了系统发育树,其中麻疯树JcRIPII并没有与其他大戟科中的序列表现出较近的亲缘关系,表明JcRIPII可能存在不同于大戟科其它II型RIP如ricin的进化起源。为探究这一现象是否可能是由于RIP和凝集素结构域进化之间的明显差异所致,所以我们分别对这两个结构域区的序列进行了系统发育分析。已有研究表明,大戟科中的I型RIP由原始的I型RIP进化而来,与RBL结构域融合形成II型RIP,部分II型RIP凝集素结构域的缺失重新变成I型RIP[17]。在RBL结构域构建的进化树,蓖麻中的II型RIP的B链序列位于麻疯树和茶等多种植物构成的分枝外侧,与JcRIPII距离较近,表明这一分枝上的II型RIP可能是源于自身I型RIP和蓖麻中凝集素基因的融合。在RIP结构域构建的进化树中,JcRIPII与海桐、橡胶树II型RIP构成的分枝和麻疯树中的I型RIP相近而不相邻,与蓖麻中II型RIP在进化树上关系更远,中间甚至混合了来自单子叶植物的RIP。这种镶嵌分布反映了RIP结构域复杂的进化方式,RIP的起源应该较早,至少早于单双子叶植物分化以前。综上,JcRIPII与樟科、山茶科、棕榈科较近的亲缘关系是因为它们B链中的凝集素结构域可能都是源于蓖麻,A链中起源较早的RIP在进化的过程中经历了多种进化方式,造成了同科、同种植物中RIP结构域的差异。
组织表达的结果显示JcRIPII基因在胚乳中具有相对较高的表达量,与ricin等在种子高表达特点具有统一性[18]。同时JcRIPII基因在侧根中具有比主根更高的表达,增加侧根是植物抵御干旱的生理机制的主要器官,结合JcRIPII基因上游启动子区中存在的干旱胁迫相关顺式调控元件,表明JcRIPII基因可能也在干旱胁迫中发挥作用[19]。I型RIP基因的表达在干旱或聚乙二醇处理下往往上调,许多Ricin_B蛋白在逆境胁迫下也是诱导表达,而JcRIPII基因随干旱时间的延长先下调再上调,表现出一定的特异性,后续还需要进行更多的研究来阐明JcRIPII的作用方式[20,21]。
猜你喜欢
江汉大学学报(自然科学版)(2022年4期)2022-09-02
湖北农业科学(2022年11期)2022-07-18
老友(2020年11期)2020-11-28
中国诗歌(2018年3期)2018-11-14
食品界(2017年7期)2017-08-24
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
医学美学美容·中旬刊(2015年1期)2015-10-21
创新作文(3-4年级)(2015年8期)2015-09-15
