
晚中新世C4植被扩张与大气二氧化碳分压的关系
2021-11-05 02:18汪镇田军
海洋地质与第四纪地质 2021年5期
汪镇,田军
同济大学海洋地质国家重点实验室,上海 200092
晚中新世(约11~5.3 Ma)是新生代变冷以来值得关注的一个时期,地球气候系统发生了一系列重大的变化,包括晚中新世大洋碳位移(Late Miocene Ocean Carbon Shift, LMOCS)事件[1-9]、C4植被扩张事件[10-12]等。在海洋系统,全球底层海水溶解无机碳的碳同位素在7.7~6.6 Ma发生了振幅约1‰的显著负偏,即LMOCS事件[1,3]。由于表层海水有着相似的负偏记录[9,13],涉及到整个海洋碳库的变化,因此,LMOCS事件与陆地输入有紧密的联系,表现为陆地有机碳库对海洋无机碳库碳同位素的影响,而导致这种现象的原因可能是陆地C4植被的大规模扩张[2,14]。C4植被的大规模扩张是晚中新世重要的气候事件,土壤碳酸盐、叶蜡正构烷烃等的δ13C值[15-24]变化证明,8~6 Ma陆地生态系统中碳同位素偏重的C4植物占据主导地位,由此也引发了一系列的气候效应[25-26]。基于C4植物生长的适宜环境条件[27]进行考虑,扩张的主要原因通常有两种:一是气候变化[28]包括季节性气候增强以及引发的火灾,二是大气CO2浓度的下降[12]。但早期的大气pCO2重建结果[29]和相关水文变化[30]的研究却直接否认了大气CO2的影响,而强调季节性气候在C4植被扩张中的重要作用。但是最近的大气pCO2重建记录[31-32]却显示,在晚中新世特别是LMOCS时期,大气CO2浓度处于下降的趋势,同时依据C4植被在晚中新世的地理扩张范围而得到的数值模拟实验结果也显示,满足C4植被在晚中新世扩张所需的大气CO2浓度与地质记录重建的大气CO2浓度在数值上[33]相吻合,一定程度上为大气CO2浓度变化引起C4植被扩张的假说提供了证据支持(如图1所示)。

图 1 实验模拟与地质记录揭示的晚中新世C4植被扩张范围中间地理图为实验模拟[33]的400×10−6大气pCO2条件下的C3/C4植被全球分布;上、下图,地质记录[15-24](土壤碳酸盐、叶蜡正构烷烃等的δ13C值)显示的晚中新世C4植被扩张范围,主要包括:非洲东部和西北部[18,22](赤道东大西洋659、959站,赤道西印度洋241、235站)、非洲南部和西南部[20-21](南大西洋1081、1085站)、印度西南部[24](阿拉伯海U1457站)、尼泊尔[23]、巴基斯坦[16]、北美大平原[19]、阿根廷西北部[15]、中国西北部[17]。Fig.1 The reconstructed Late Miocene C4vegetation expansion areas based on numerical simulations and geological records Middle, the geographical map shows the global distribution of C3/C4vegetation under 400×10-6atmospheric pCO2in the experimental simulation. Upper and lower, the geological records[15-24](δ13C values of soil carbonate, leaf paraffin, etc.)indicate that the C4vegetation expansion areas in the Late Miocene mainly include the east and northwest of Africa[18,22](eastern equatorial Atlantic site 659, 959, the equatorial western Indian Ocean site 241, 235), the south and uthwest of Africa[20-21](South Atlantic site 1081, 1085), the southwest of India[24](Arabian sea site U1457), Nepal[23], Pakistan[16], The Great Plains[19], the northwest of Argentina[15], and the northwest of China[17].
本文将首先介绍大气pCO2的重建方法以及在晚中新世时期的研究现状,然后具体阐述晚中新世C4植被扩张事件及其影响,最后探讨C4植被扩张与大气pCO2的关系及相关争议。
1 晚中新世的大气pCO2记录
1.1 重建大气pCO2的方法
目前重建大气pCO2的方法有很多,冰芯气泡中可以直接捕获最近几十万年的大气CO2记录,而在更长的时间尺度上,通常利用古植物叶片的气孔指数[34-36]、古土壤碳酸盐的碳同位素[37-39]、浮游藻类生物标志物的碳同位素[29,40-42]以及浮游有孔虫壳体的硼同位素[32,43-45]等方法来实现地质历史时期大气pCO2的重建(表1)。
植物主要通过叶片气孔与大气进行CO2、水蒸气等成分的交换。防止水分流失以及最大限度吸收CO2是植物生长的有利条件,高大气pCO2下会使植物减少气孔打开来保持水分,而更低的CO2浓度则允许气孔增加以维持碳的同化速率,因此气孔的形成一定程度上受到CO2浓度的控制[35]。气孔密度(SD)是单位面积叶片上气孔的数量,能够反映叶片气孔形成时的大气pCO2情况,由于SD不仅与气孔数量有关,与表皮细胞的生长面积也有联系,受到温度、湿度、光照等外部条件的控制,因此引入气孔指数(SI)对表皮细胞面积规范化处理来消除其他因素的影响[35],突出CO2的作用。根据现有的植物属种(如银杏)归纳出SI与CO2浓度的经验关系[36],将此关系推广到亲缘的古植物属种以重建古大气pCO2(表1)。
土壤碳酸盐是在土壤风化成土过程中形成的次生碳酸盐,多形成于年降雨量较低的干旱到半干旱地区。当处于降水增多的土壤生物生长时期,原生碳酸盐受到土壤生物呼吸释放的CO2影响而发生溶解,释放的钙离子随着水分不断向下渗透,当蒸发作用大于降水作用时发生碳酸钙的再沉淀,形成次生碳酸盐[39]。土壤碳酸盐的碳同位素组成直接取决于土壤中的CO2,而土壤中的CO2主要是大气CO2与土壤生物呼吸产生CO2的混合物,因此利用Cerling[37]提出的扩散反应模型可以推导出大气pCO2的计算公式(表1)。
浮游藻类的生物标志物主要指长链烯酮,烯酮碳同位素重建古大气pCO2的方法依赖于光合作用的碳同位素分馏与被动扩散模型[40]。大气CO2溶解于海水并被藻类生物利用进行光合作用,发生碳同位素的分馏,分馏值(εp)可以通过溶解CO2和生成有机物的碳同位素直接计算,而影响εp的控制因素有溶解CO2浓度、细胞生长速率、细胞体积与表面积比值等[46]。被动扩散模型可以用来解释εp与影响因素之间的关系,CO2由于细胞内外浓度的梯度差进入或流出细胞,到达细胞内的叶绿体进行光合作用形成有机碳,这两个过程分别称为碳的运输和碳的固定,利用过程中的分馏关系以及藻类的相关生长参数,可以总结出εp与溶解CO2浓度间的计算公式[40],根据亨利定律[43]进一步重建大气pCO2(表1)。
浮游有孔虫壳体的硼同位素重建古大气pCO2是基于海水中硼酸根(B(OH)4-)与硼酸(B(OH)3)转化过程中的硼同位素分馏。B(OH)4-与B(OH)3的平衡转化存在明显的同位素分馏现象,B(OH)4-富集相对轻的10B,而B(OH)3以重的11B为主。通过硼酸体系的反应方程与分馏系数、海水总硼的硼同位素组成,可以推算出海水pH值与δ11B(OH)4-值的计算关系[43]。浮游有孔虫壳体δ11BCaCO3值通常与海水δ11B(OH)4-值密切相关,主要受到生命效应、分析仪器的影响[44],存在一定的经验关系。因此利用浮游有孔虫壳体的硼同位素能有效反演海水pH值,并结合碳酸盐系统的第二个参数[43]以达到重建大气pCO2的目的(表1)。

表 1 古大气pCO2的重建方法Table 1 Methods for reconstruction of ancient atmospheric pCO2
重建大气pCO2的每种方法都存在一定的局限性。由于化石植物的表皮细胞壁模糊不清,古植物叶片方法在统计气孔指数时存在困难,另外树叶的阴叶和阳叶又具有不同的气孔指数[47],同时气孔指数对高浓度大气CO2的响应极其不敏感[48],因此化石保存程度、阴阳叶混杂、高浓度大气CO2都会使重建结果造成偏差。古土壤碳酸盐容易受到成岩作用的改造,在样品选取时需要注意识别并避免成岩作用较强的次生碳酸盐。另外对于参数S(z)尚未有可靠的测量或计算方法,这种不确定性对结果会产生很大的差异[39]。浮游藻类的烯酮方法依赖于被动扩散模型[49],但是并非所有产烯酮藻类都符合,同时参数b值[50]即总碳同位素分馏的生理作用值具有不确定性,这也一直是该方法需要攻克的重要难题。有孔虫硼同位素方法在计算海水pH值时有化学原理与公式的支撑[51],具有可靠性,但是进一步求解大气pCO2还要结合碳酸盐体系的第二个参数[43],需要进行必要的假设与推算,因此会使重建结果产生误差。由于大气pCO2的重建方法仍处于探索性阶段,并且CO2在地质记录上存在大量的空白,因此无论哪种方法取得的pCO2结果都具有建设性意义。
1.2 晚中新世大气pCO2的研究现状
前人对古大气pCO2的研究主要聚焦于地质历史时期大气CO2的长期变化趋势上。Ekart等[38]汇编了已有的古土壤碳酸盐的碳同位素数据集,估计出400 Ma以来的大气pCO2演化历史;Retallack[36]则依据现代银杏亲缘属种的叶片化石,提供了300 Ma以来的CO2重建记录,这些都为地球气候系统长期变化的研究提供了数据基础。在更短的时间跨度上,Beerling和Royer[52]汇总了新生代65 Ma以来不同方法重建的大气CO2记录,发现了大气CO2与底层海水温度基本保持一致的变化趋势。更高分辨率的大气CO2记录主要集中于中中新世[44,53]和上新世以来的时期[54-55],尚未有学者针对晚中新世时期系统性建立高精度的大气pCO2记录,尽管已有的记录均为一些零星或低分辨率的数据,但是也具有一定的参考价值。
图2总结了晚中新世时期的大气pCO2数据,主要为古植物叶片、古土壤碳酸盐、浮游藻类以及浮游有孔虫方法的重建结果[29,31,32,52,56-57]。浮游藻类方法的重建数据相对较多,最早Pagani[29]的烯酮记录显示,晚中新世大气CO2浓度维持在(250~320)×10−6,变化波动较小,且早在渐新世末,大气CO2浓度就已经下降到工业革命前的低水平,进入中新世后一直处于上升的状态,不支持大气CO2浓度下降导致C4植被扩张的观点。但是最近Mejia[31]的烯酮记录却表明,大气CO2浓度从11 Ma的约454×10−6下降至6 Ma的约250×10−6,其他重建结果例如Sosdian[32]的浮游有孔虫方法,也支持在晚中新世尤其是LMOCS时期,大气CO2浓度存在下降的趋势。另外,Holbourn[13]利用浮游有孔虫与底栖有孔虫的碳同位素差异来反映大气CO2的变化,也体现出7.5~7 Ma的降低趋势。综合所有重建方法的结果,晚中新世时期的大气CO2浓度为(200~400)×10−6,并且整体呈现出缓慢的下降趋势,而在进入上新世之前有所上升。从长时间尺度的碳循环来看,岩浆的脱气作用与大陆硅酸盐化学风化作用整体上调控着大气CO2的浓度。Raymo 和 Ruddiman[58]提出著名的“构造抬升−风化”假说,认为青藏高原的隆升使得更多的岩石暴露到地表并发生碎裂,增加与大气的接触面积,促使大陆硅酸盐风化加强,消耗大气CO2。在晚中新世时期,相关的地质记录表明喜马拉雅和青藏高原地区发生了一次大规模隆起[59],因此该时期大气CO2浓度的下降可能与这次构造运动有一定的联系,风化作用增强,更多的大气CO2进入到陆地碳库。同时,该时期的海洋生物勃发事件[3]也促使了大气pCO2的降低,通过增强的生物泵作用将CO2封存至深海碳库,在晚中新世全球变冷的背景条件下,海洋加强的溶解泵作用也对大气CO2浓度的降低起到了重要的作用。由于该时期大气pCO2重建记录的分辨率较低,不同方法获得的结果也有一定的差异,因此不排除大气pCO2存在上升的可能,需要后续的相关研究进行补充,从而更精确地识别大气pCO2的变化。
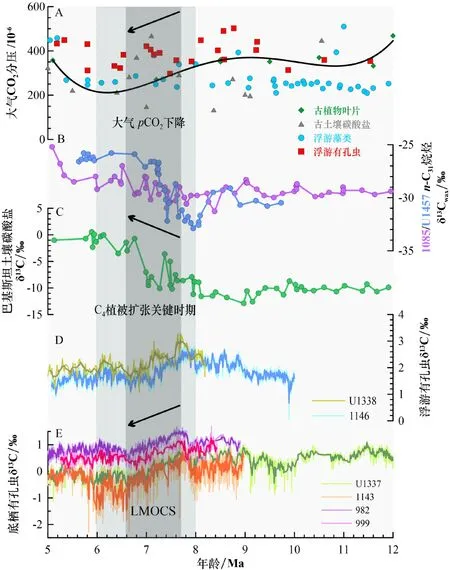
图 2 大气pCO2、C4植被扩张和晚中新世大洋碳位移事件A.不同指标重建的大气pCO2[29,31,32,52,56-57],包括古植物叶片[52,56,57]、古土壤碳酸盐[52,56-57]、浮游藻类[29,31,52,56-57]、浮游有孔虫[31-32,56-57];B, C.不同指标指示的C4植被扩张事件[16,20,24]:南大西洋1085站、阿拉伯海U1457站叶蜡正构烷烃[20,24]、巴基斯坦土壤碳酸盐[16]的δ13C值;D, E.浮游[9,13]与底栖有孔虫[2,4,7-8]δ13C值记录的LMOCS事件:赤道东太平洋U1338站[9]、南海1146站[13]、赤道东太平洋U1337站[8]、南海1143站[7]、北大西洋982站[4]、加勒比海999站[2]。图中浅色柱为C4植被扩张关键时期(8~6 Ma),深色柱为LMOCS事件(7.7~6.6 Ma),A中曲线为四次多项式拟合趋势,D,E中浅色、深色曲线分别为原始数据、5点滑动平均后数据。Fig.2 Atmospheric pCO2and C4vegetation expansion and the LMOCS(Late Miocene Ocean Carbon Shift)A. Atmospheric pCO2reconstructed by different proxies[29,31-32,52,56-57], including leaves of the ancient plants[52,56-57], paleosoil carbonates[52,56-57], planktonic algae[29,31,52,56-57], and planktonic foraminifera [31-32,56-57]; B, C. C4vegetation expansion indicated by different indices[16,20,24]: δ13C values of leaf paraffins[20,24]from South Atlantic site 1085, Arabian Sea site U1457, and soil carbonates[16]in Pakistan. D, E. LMOCS recorded by planktonic[9,13]and benthic foraminifera[2,4,7-8]δ13C values: eastern equatorial Pacific site U1338[9], South China Sea site 1146[13], eastern equatorial Pacific site U1337[8], South China Sea site 1143[7], North Atlantic site 982[4], and Caribbean Sea site 999[2]. In the figure, the light colored column represents the critical period of the C4vegetation expansion(8~6 Ma),and the dark colored column represents the LMOCS(7.7~6.6 Ma). The curve in A represents the Quaternary polynomial fitting trend, and the light and dark curves in D and E are the original data and the 5-point moving average data respectively.
2 晚中新世C4植被扩张的生态系统格局
如图3所示,植物在全球碳循环系统中扮演着重要的角色,通过光合作用、呼吸作用与大气发生直接的碳交换,然后利用海气作用包括溶解泵、生物泵和碳酸盐泵来间接影响海水的碳同位素组成,另外还可以经过土壤实现陆地碳库对海洋碳库的碳输入。追踪地质历史时期陆地植物群落的组成,即C3/C4植被的演变,除了直接鉴别植物化石种属,主要还是依据C3与C4植物的碳稳定同位素(δ13C)差异。C3植物的碳同位素值范围很广,从-35‰到-20‰,平均约-27‰;C4植物有着更窄的碳同位素范围,从-14‰到-10‰,平均约-12‰,明显大于C3植物[61]。地质记录中通常利用草食性哺乳动物牙化石[12]、土壤碳酸盐[10]、植物叶蜡[62]等的δ13C值变化,间接分辨出陆地C3与C4植物的相对比例。

图 3 C3/C4植被参与的全球碳循环过程[60]Gt为十亿吨。Fig.3 Global carbon cycle processes involving C3/C4vegetation [60]Gt is a billion tons.
光合作用的原始途径,即C3光合作用早在2800 Ma前的高大气CO2浓度下就已形成[63],C4光合作用则是为了适应地质历史时期气候环境的演变,而从C3光合作用中进化而来。在32 Ma左右发现隶属于C4植物谱系的虎尾草亚科记录[64],证实了渐新世时期C4植物已经开始出现。大多学者都认为这种C4光合作用的进化是对大气CO2浓度下降的响应[65]。大气pCO2重建记录显示[41],始新世时期的大气CO2浓度为(1000~1500)×10−6,而进入渐新世,大气CO2浓度在32~25 Ma期间突然下降至500×10−6以下,刚好与C4植物出现的时间相吻合,同时相关的模拟实验结果也证实,该时期大气CO2浓度下降对C4光合作用的进化产生了重要影响[64]。但直到晚中新世时期,C4植被才开始出现第一次大规模的扩张。
C4植被在晚中新世时期的扩张范围主要覆盖非洲、南亚以及北美地区。Cerling[12]收集了亚洲、非洲、北美洲和南美洲等区域哺乳动物牙化石的碳稳定同位素记录,发现8~6 Ma期间出现明显的偏重现象,意味着晚中新世时期的陆地植物有着更高的δ13C值,预示C4植被发生大规模扩张。C4植被的扩张并不是全球同步开始的,最早发生于非洲的低纬度地区。Uno[18]对东非地区晚中新世至上新世牙化石的碳同位素数据进行统计分析,发现大部分草食动物的饮食习性在9.9 Ma时已经偏向于C4植物;Polissar[22]发现非洲东部与西北部的深海沉积物中,植物叶蜡正构烷烃的δ13C在10 Ma左右出现显著的正偏,表明C4植被的覆盖率在非洲地区开始增加。而在非洲的南部与西南部,C4植被则在8 Ma左右相继增加[20-21]。随后C4植被在中纬度的南亚地区出现大规模扩张,使之也成为晚中新世C4植被扩张的主要区域之一。Freeman[66]提取了尼泊尔和巴基斯坦晚中新世时期古土壤中的植物叶蜡,发现其正构烷烃的δ13C在8~6 Ma出现显著正偏;Behrensmeyer[16]发现巴基斯坦中北部土壤碳酸盐的δ13C在8~5 Ma明显增高;Neupane[23]对尼泊尔地区河流沉积进行研究,发现其中植物叶蜡正构烷烃的δ13C在8.5 Ma左右也开始逐渐变化;Feakins[24]在印度西部的深海沉积物中,发现植物叶蜡正构烷烃的δ13C在7.4~7.2 Ma大幅度变化。这些记录都指示南亚地区的C4植被可能在8~6 Ma开始大规模扩张。在同一纬度上北美洲的大平原区域,古土壤的δ13C记录显示,C4植被从晚中新世至早更新世呈现阶段性扩张[19]。而在东亚、南美洲、澳大利亚等地区,C4植被的扩张相对滞后[15,67-68],发生于早上新世时期,对应C4植被第二次大规模扩张的时间,基本形成现今的C3/C4植被分布的生态系统格局。
晚中新世C4植被的扩张对全球气候产生了重要的影响。开阔的草原取代茂密的森林,最直接的影响便是地面反照率的增强与蒸腾作用的减弱,促使气候变得干冷[25],可能与该时期的全球气候变冷[26]有关。另外,植被类型的演变使得整个生物圈的群落结构发生重大的变化,马科动物的分异度在晚中新世达到鼎盛,发生大规模灭绝,马的牙齿由低冠齿演化为高冠齿,体型也由小变大、由低变高[69];由于草类特有的植物硅酸体的硅更易溶于水,C4植被的繁盛促进了大陆分化向海洋输送硅的能力,使海洋硅藻得以迅速发展,成为现在常见的浮游植物之一,这些都体现了C4植被的扩张所引起的生物圈内部的协同进化[25]。更重要的是C4植被扩张对全球碳循环过程的深远影响,草原土壤的有机质含量相对于森林更高,并且更加容易侵蚀,晚中新世陆地向海洋和河流输入的有机碳增加[70],同时C4植物的δ13C值明显大于C3植物,陆地扣留了更重的碳同位素组分,轻组分输送到海洋碳库,因此,C4植被扩张可能是触发LMOCS事件的原因之一。
3 晚中新世C4植被扩张与大气pCO2的关系
3.1 CO2与C4植物生长
陆地生态环境中的光合作用主要有三类固碳方式:C3途径(Calvin循环)、C4途径(Hatch-Slack循环)和CAM途径(固碳方式与C4途径相似,但时间与空间不同)。C3植物在光合作用中仅以Calvin循环的方式固碳,CO2进入叶肉细胞,与叶绿体中的受体1, 5-二磷酸核酮糖(RuBP)结合,在羧化酶的催化下形成最初的产物三碳化合物3-磷酸甘油酸(PGA),再经过一系列还原反应最终生成有机物和新的RuBP。而C4植物除了Calvin循环外还有其他固碳途径,通过碳浓缩机制实现对大气CO2的充分利用,如图4所示。CO2进入叶肉细胞首先与磷酸烯醇式丙酮酸(PEP)受体结合,羧化酶催化下固定为四碳化合物草酰乙酸(OAA),进一步还原为苹果酸(MAL),然后转移到维管束鞘细胞中通过脱羧酶作用释放CO2和产生丙酮酸(PVA),PVA返回叶肉细胞中循环利用,而释放的CO2在鞘细胞的叶绿体中进行Calvin循环生成有机物[27],相对于C3植物叶肉细胞中低浓度的CO2,C4植物维管束鞘细胞中CO2浓度始终维持在一个较高的水平。
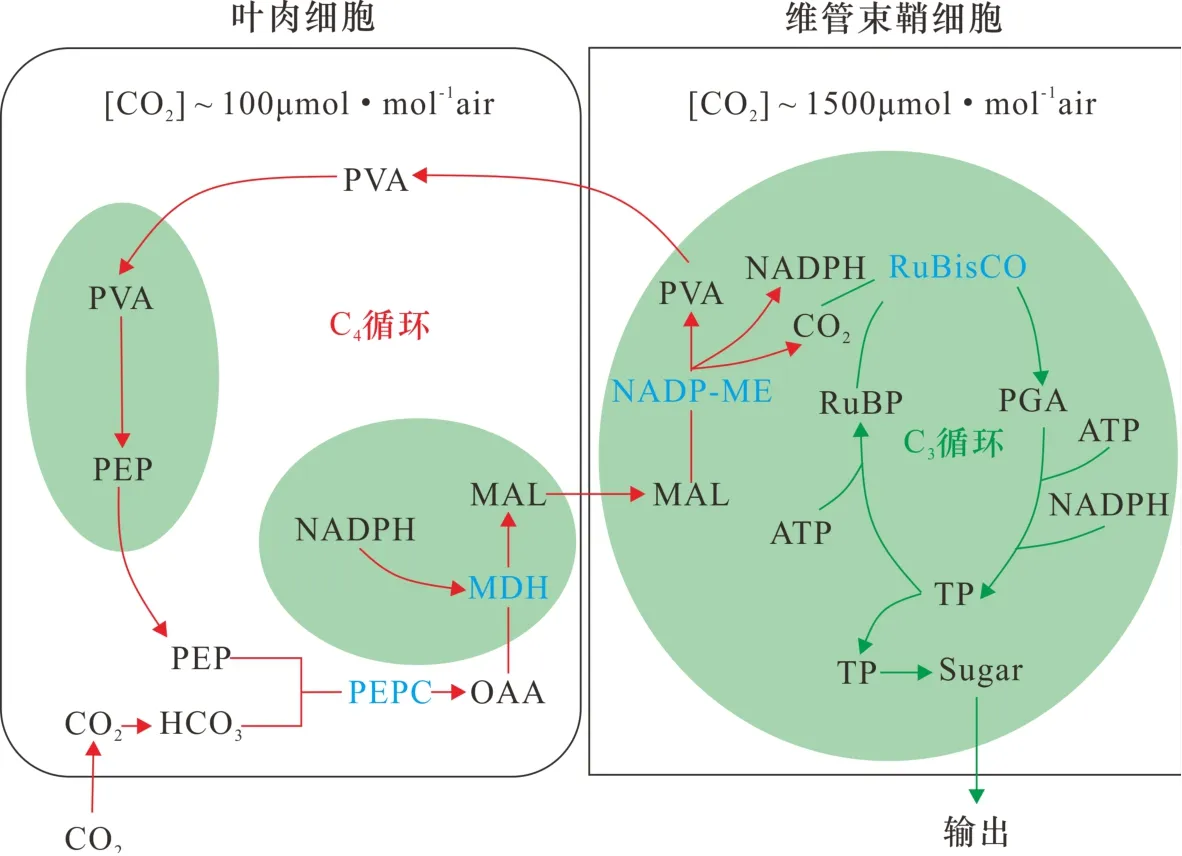
图 4 C4植物的CO2浓缩机制[27]C4植物在叶肉细胞中进行C4循环,富集和输送CO2至维管束鞘细胞中进行C3循环(Calvin循环),而C3植物仅在叶肉细胞中进行C3循环。其中,PEP是磷酸烯醇式丙酮酸受体,PEPC是PEP羧化酶,OAA是四碳化合物草酰乙酸,MDH是苹果酸脱氢酶,MAL是苹果酸,NADP-ME是烟酰胺腺嘌呤二核苷酸磷酸-苹果酸酶,PVA是丙酮酸,RuBP是1, 5-二磷酸核酮糖受体,RuBisCO是RuBP羧化酶,PGA是三碳化合物3-磷酸甘油酸,TP是磷酸丙糖。Fig.4 CO2concentration mechanism of C4vegetation [27]C4vegetation carries out C4cycle in mesophyll cells, enriching and transporting CO2to vascular bundle sheath cells for C3cycle(Calvin cycle), while C3vegetation only carries out C3cycle in mesophyll cells. PEP: phosphoenolpyruvate; PEPC: PEP carboxylase; OAA: oxaloacetic acid; MDH: malate dehydrogenase; MAL: malate; NADP-ME: NADP-malic enzyme; PVA: pyruvate; RuBP: ribulose-1, 5-bisphosphate; RuBisCO: RuBP carboxylase; PGA: 3-phosphoglyceric acid; TP: triose phosphate.
总之,C4植物并不是直接将CO2转化为有机物,而是通过改善CO2的供应方式来辅助固碳和还原,即在Calvin循环之前先利用叶肉细胞来富集和储存CO2并输送至维管束鞘细胞[71],同时PEP羧化酶(PEPC)对CO2的活性远远强于RuBP羧化酶(RuBisCO)[72],因此相较于C3植物,C4植物对CO2的亲和力更大,能够在更低浓度的CO2环境下生存。此外,由于这种碳浓缩机制会使得维管束鞘细胞中的CO2浓度长期处于较高的水平[27],高CO2/O2比率减少RuBisCO引起的副反应即氧化反应,抑制了光呼吸作用的能量和水分消耗[73],有利于光合产物的积累,同时降低了C4植物对外界CO2吸收的依赖,从而允许叶片减少气孔的打开,避免了水分的丢失,更能适应干旱的环境。这种光合作用上存在的差异导致两者的生活环境有所不同[74],C3植物通常适宜生活在温度较低和气候潮湿的环境,例如温带草原和高海拔地区,主要为乔木、灌木植物;而C4植物在高温、干旱、强光和低CO2浓度等环境下占据优势,生活在热带、亚热带草原和半干旱地区,主要是草本植物。
温度、水等环境要素在植物生长过程中固然扮演着重要的角色,而CO2作为光合作用的必要条件,在调控C3/C4植被的竞争关系时起到了关键性作用,影响到植被群落的全球分布。因此基于C3/C4植被生长的光合作用原理,晚中新世的C4植被扩张事件可能与大气CO2存在着紧密的联系。
3.2 晚中新世C4植被扩张假说及争议
晚中新世C4植被扩张的原因一直以来都饱受争议,主流的观点也是建立在适宜C4植物生长环境条件的基础上,包括气候变化与大气CO2浓度变化。
3.2.1 气候变化假说
与气候变化相关的假说主要考虑季节性气候对C4植物生长的影响[28]。原理上,季节性降雨增强有利于C4植物生长,季节性干旱增强有可能消除C3植物[28],因而营造出有利于C4植被竞争的环境优势;除了季节性气候带来的干旱与降雨的直接影响外,火灾效应也对C4植被扩张起到重要的作用[75],季节性气候增强会使生物量在雨季得到快速增加,为旱季提供更多的燃料从而引起火灾频发[76],因此会消除森林,阻止C3植被中木本植物的生长,让适应火灾环境的草原[77]占据更多的生态空间。同时模型和现代观测都表明,火灾与季节性降雨对维持亚热带草原目前的全球分布至关重要[78]。
如图5所示,Quade[10]发现巴基斯坦土壤碳酸盐的δ13C在7.4~7 Ma出现显著正偏,反映陆地C4植被的扩张,与此同时δ18O也发生明显的变化并且稍微领先于δ13C的变化,而通常土壤碳酸盐的δ18O受控因素较多,包括温度、降雨同位素、季节性降雨等,在没有进一步证据的情况下,只解释为当地某种气候变化引起了碳循环的变化。而依据该地区现今季风性气候较强而有利于C4植物生长,并结合晚中新世北印度洋硅藻组合的季风重建指标[10],推测可能是亚洲季风的加强促使了C4植被扩张。

图 5 降雨量、季风强度、火灾强度、C3/C4植被比例的替代性指标A, B. 亚丁湾231站植物叶蜡n-C30烷酸的δD与δ13C值[30,79],C.巴基斯坦土壤碳酸盐δ18O值[80],D.巴基斯坦古土壤多环烷烃浓度/n-C31烷烃浓度[76],E.巴基斯坦土壤碳酸盐δ13C值[80]。A, B中阴影部分解释了非洲东北部地区季节性干旱引起C4植被扩张,C, D, E揭示了亚洲季风、火灾活动与C4植被扩张的关系。Fig.5 Proxies of rainfall, monsoon intensity, fire intensity, and C3/C4vegetation ratioA, B. δD and δ13C of n-C30alkanoic acid from plant leaf wax in the Gulf of Aden site 231[30,79]; C. δ18O of soil carbonate in Pakistan[80]; D. Polycyclic alkane concentration/ n-C31alkane concentration in paleosoil of Pakistan[76]; E. δ13C values of soil carbonate in Pakistan[80]. In A and B, the shadowed part explains the C4vegetation expansion caused by seasonal drought in Northeast Africa. C, D and E highlight the relationship among the Asian monsoon, the fire activity andthe C4vegetation expansion.
同样在非洲东北部区域,Feakins[30]对植物叶蜡δD进行了分析,发现在11~9.7 Ma存在40‰的正偏,植物叶蜡δD主要反映的是降水δD的变化,而降水δD在热带地区不受温度效应的控制[81],仅与降雨量有关,意味着非洲东北部区域在该时期可能处于全面干旱的状态,来自印度洋水汽的输入减弱[30],而植物叶蜡δ13C显示的C4植被扩张时间为11~8.7 Ma[79],因此当地的水文循环的变化可能驱动了碳循环的变化。
除了干旱与降雨,季节性气候引起的火灾[76]也是影响C4植被扩张的重要因素。Karp[76]在巴基斯坦地区的相关研究发现,土壤碳酸盐δ18O变化表明10 Ma左右亚洲季风开始增强,而多环芳烃(PAH)的重建记录显示火灾在10 Ma之后特别是8~6 Ma频繁发生,同时伴随着松柏科植物燃料的输入减少,可能意味着季风的增强促进了火灾的形成,火灾的持续加强伴随着植物群落组成的改变,导致C3/C4植被的演化更替,最终从8 Ma开始,C3林地被更能适应火灾的C4草原所替代。Feakins[24]在印度河扇东部重建的木炭记录也显示了8~6.5 Ma草源木炭的增加,这些都证实了火-草的反馈机制[77]使得C4植被进一步大规模扩张。在非洲西南部地区,Hoetzel[21]也发现了8~3 Ma的C4植被扩张时期,烧焦粒子的比例在7.1~5.8 Ma有明显的增加,在5.8 Ma后木炭含量的降低与荒漠、半荒漠植被花粉比例的增加,意味着出现干旱期而减少了燃料供应,同样强调了火灾在C4植被扩张过程中的重要作用。
但是,季节性气候增强以及引起的火灾都是区域性的气候现象,而C4植被扩张是全球事件[12],尽管存在异步性,部分地区仍没有发现水文循环与碳循环的一致变化或者相关的火灾记录,因此气候变化驱动晚中新世C4植被扩张的观点受到质疑。Polissar[22]在非洲东部与西北部地区发现,降雨量的替代性指标植物叶蜡正构烷烃的δD在C4植被扩张期间基本没有发生变化,且风尘通量记录显示大陆干旱化加剧发生于C4植被快速扩张之后。类似的,Feakins[24]在印度河扇东部的研究也没有发现植物叶蜡正构烷烃δD的明显偏移。然而,目前重建的晚中新世植物叶蜡正构烷烃δD的记录[22,24,30]分辨率都较低,未来的工作可以通过提高分辨率来探索晚中新世水文气候变化。同时,由于降水同位素探测水文气候变化的敏感性因气候机制和地区而异,不同区域水文气
候替代性指标指示的气候变化可能不同,例如降水量、水汽来源等[81],需要联合使用其他指标来测试与检验。另外,火灾与C4植被扩张的因果关系也受限于火灾记录的分辨率[21,76]以及年龄框架的精准性[82],需要进一步的实验与研究以确认两者的先后顺序。
3.2.2 大气CO2浓度变化假说
该假说强调大气CO2浓度降低会突出C4植物的竞争优势,有利于C4植被扩张。C4植物的碳浓缩机制会使细胞内维持较高浓度的CO2,抑制了光呼吸作用的消耗[73],同时减少对外界CO2浓度的依赖,在低大气CO2浓度下能正常且高效地进行光合作用,而C3植物受限于外界CO2浓度,光合作用的效率会明显降低,光呼吸作用加强,因此缺少足够的有机物供给而死亡,逐渐被C4植物取代,早在渐新世初期,重建记录[41]与实验模拟结果[64]就已经证实大气CO2浓度的下降促使了C4植物的进化。
考虑到C4植被扩张的全球性,及与全球碳收支平衡的紧密关联[11],大气pCO2在C4植被扩张过程中的作用受到越来越多的重视。Cerling[11-12]发现,C4植被是在全球范围内扩张的,认为可能与大气CO2浓度的下降有关,并依据随大气CO2浓度和温度变化的C3/C4植被量子产率模型[83],用不同纬度的温度下CO2阈值的差异来解释C4植被在全球扩张的异步性。Polissar[22]的研究表明,C4植被的扩张与全球表层海水温度的降低在时间上有很好的对应关系,而全球平均海温的持续下降、高纬度地区的降温作用增强,与大气CO2浓度的下降在高纬度海区的反馈一致,因此也强调了大气CO2浓度下降与C4植被扩张的紧密联系。同时,C4植物生长的土壤相较于C3植物更厚,能储存更多的CO2,这种正反馈作用会降低大气CO2浓度而加快C4植被的扩张进程[82]。
气候模拟实验[33]也为这一观点提供支持。Zhou[33]利用中中新世条件下的全耦合气候模型,模拟了各种气候驱动因素,包括CO2、温度、水和光照在全球范围内对C3/C4植被竞争优势的控制。实验结果表明,当大气pCO2处于600×10−6时,C4植被在全球几乎都不占据优势,主要是水限制了C4植被的分布;而当pCO2处于400×10−6时,非洲东北部和亚洲中南部的生态系统出现明显的C4植被主导,与记录中晚中新世C4植被扩张的区域较为一致;当pCO2最终降低至270×10−6时,C4植被在北美洲、澳大利亚地区出现大面积覆盖,超过记录中晚中新世的覆盖范围,近似于上新世的扩张范围[84]。最新的大气pCO2重建记录[31-32]显示,晚中新世时期特别是8~6 Ma期间,大气pCO2处于下降的趋势,非常适宜C4植被的生长与扩张,并且重建数值在(200~400)×10−6的水平,与数值模拟实验中晚中新世C4植被扩张区域的大气CO2浓度[33]一致,因此大气CO2浓度无疑是驱动晚中新世C3/C4植被更替的关键气候因素。
对于大气CO2浓度下降引起C4植被扩张的观点,目前最不明确的就是尚未有准确可靠的高分辨率大气pCO2重建记录。早期的重建记录就不支持这一看法,比如Pagani[29]的大气pCO2重建结果显示,早在渐新世末,大气CO2浓度就已经下降到工业革命前的低水平,与C4植被扩张的时间相差15 Ma左右,不支持CO2驱动C4植被扩张。最新的重建结果[31-32]尽管在趋势和数值上都支持这一观点,但是分辨率相对较低,并且如前文所述,不同方法在重建大气pCO2时都存在差异与局限,因此有待于后续研究补充更加单一可靠的高分辨率记录。
4 结论与展望
陆地与海洋的地质记录显示晚中新世C4植被扩张的主要区域包括非洲、南亚、北美地区,扩张的关键时间为8~6 Ma,由此也引发了一系列的气候效应,例如晚中新世变冷事件、生物圈的协同进化、LMOCS事件等,体现出C4植被扩张对全球气候系统的影响。而晚中新世C4植被扩张的原因至今仍存在疑惑,主要在气候变化与大气CO2浓度变化中产生争论,又或许全球性的大气CO2浓度下降以及区域性的气候变化都起到了重要的作用,受限于重建指标的分辨率与可靠性,这一问题尚无明确的定论。
在最新大气pCO2重建记录与气候模拟实验的支持下,CO2的作用开始被重视起来。大气pCO2重建记录呈现晚中新世大气CO2浓度处于下降的趋势,有利于C4植被的生长与扩张;并且考虑到晚中新世C4植被的扩张区域,地质记录重建的大气CO2浓度与数值模拟实验中形成C4植被扩张所需的大气CO2浓度在数值上相吻合。由于不同方法在重建大气pCO2时都存在一定的差异与局限,因此为了进一步探究晚中新世C4植被扩张与大气pCO2的关系,该时期使用单一方法重建的更高分辨率大气pCO2记录是必不可少的参考依据,有待后续研究补充。
猜你喜欢
科学技术创新(2022年30期)2022-10-21
河北地质(2022年2期)2022-08-22
学校教育研究(2021年24期)2021-03-28
大众科学(2020年8期)2020-11-02
科学大众(2020年17期)2020-10-27
求知导刊(2017年14期)2017-07-15
分析化学(2017年5期)2017-06-21
科技资讯(2017年13期)2017-06-21
同位素(2014年3期)2014-06-13
同位素(2014年2期)2014-04-16
