
酵母在传代过程中生长性能、碳流向及抗氧化特性的研究
2022-02-15 05:47孙英淇谢东东韩知雨任顺成
河南工业大学学报(自然科学版) 2022年6期
孙英淇,谢东东,韩知雨,任顺成
河南工业大学 粮油食品学院,河南 郑州 450001
酵母可将葡萄糖转化为乙醇和二氧化碳,作为天然发酵剂,在发酵行业中具有重要的作用。酵母是酿酒行业必不可少的原料之一,不同种类的酵母可以产出不同的白酒(浓香型、酱香型、米香型和清香型)、啤酒、葡萄酒等。酵母也被应用在烘焙[1]、酵素生产[2]、生物产能[3]、农副产品增值[4]等方面。酵母在应用过程中伴随着老化的问题,需要不断地传代保藏,在传代保藏过程中会发生衰老,严重影响发酵产物的品质[5]。
酵母的衰老主要分为两种:(1)随着酵母不断传代、分裂次数的增多,活力逐渐降低,称为复制性衰老;(2)单个酵母产生有限的子细胞的时序性衰老[6-7]。引起酵母细胞衰老的原因主要有:(1)酵母细胞端粒理论[8-9],酵母细胞在每次分裂过程中细胞端粒缩短3~4 bp,直到细胞的平均端粒长度200 bp耗尽,酵母的DNA发生损伤造成衰老。(2)酵母内源性氧化胁迫[10],酵母在代谢过程中产生活性氧(ROS)并逐渐积累,造成酵母的氧化胁迫。(3)基因组的稳定性,基因组的不稳定影响酵母DNA损伤修复,导致酵母进入衰老的过程。(4)酵母外源胁迫,因为在传代的过程中生长环境不断变化,导致酵母受到高渗、乙醇以及氧化等胁迫产生的自由基逐渐积累,对酵母造成损伤,削减其活性[11]。同时,有研究表明酵母在传代过程中有机酸的代谢呈现先升高再降低,细胞中蔗糖酶、核酸酶、脂肪酶、α-淀粉酶、乳糖酶等一些与细胞代谢相关酶的活性不断下降[12],胞内自由基含量不断升高[13],抗氧化酶的活性不断提升[14]。酵母在传代过程中细胞壁中蛋白质的种类和含量也呈现出动态变化的趋势,使酵母形态发生变化,提高了酵母的絮凝力,生长速率和降糖能力下降[15]。
然而,针对传代过程中酵母碳流向的具体研究较少。研究表明多数的胁迫环境都会引起碳代谢的改变,如冷冻[16]、高糖[17]、金属胁迫[18]等。其中,甘油海藻糖的产量均在酵母受到胁迫时提升,以保护酵母细胞的活性[19-20]。有学者通过基因的增强和切割以放大代谢效果,使酵母产出更多的保护物质,从而更好地应对胁迫[21-22]。因此,研究传代酵母的碳代谢流向的变化规律,能够为酵母抗衰老的研究提供基础。同时,通过了解酵母在传代过程中生长和发酵代谢以及细胞内活性氧含量和抗氧化酶活性变化规律,探究酵母在传代过程中衰败的关键代数和衰败规律,可以更科学高效地利用酵母。
本研究以酿酒酵母为对象,测定酵母在第1、5、10、15、20代时的生长特性和细胞膜性能,检测酵母碳代谢产物乙醇、海藻糖、甘油等物质的产量,测定超氧化物歧化酶(SOD)、过氧化氢酶(CAT)的活性及活性氧(ROS)含量,研究酵母在传代过程中代谢、氧化损伤和抗氧化能力的变化,揭示酵母细胞衰老的机制,为工业发酵提供一定的理论支撑。
1 材料与方法
1.1 酵母与培养基
活性干酵母:安琪酵母股份有限公司。
YPD培养基原料:琼脂(粉)(天津市科密欧化学试剂有限公司),蛋白胨、酵母膏(北京奥博星生物技术有限责任公司)。
1.2 仪器与设备
SW-CJ-2 D双人单面净化工作台:苏州净化设备有限公司;HPC-300BSH-Ⅲ恒温恒湿箱、DHG-907电热恒温鼓风干燥箱:上海新苗医疗器械制造有限公司;TGL-18高速冷冻离心机:四川蜀科仪器有限公司;UV-1800BPC紫外可见分光光度计:上海美普达仪器有限公司;LDZX-75KBS立式高压蒸汽灭菌锅:上海申安医疗器械厂。
1.3 试验方法
1.3.1 酵母菌的培养
采用酿酒酵母(Saccharomycescerevisiae),培养基使用YPD培养基 (1%酵母膏、2%葡萄糖和2%蛋白胨)。
接种1%活性干酵母于YPD培养基的锥形瓶中,30 ℃、180 r/min培养24 h。取1 mL菌液稀释涂布,30 ℃培养24 h后筛选出单菌落。将取出的单菌落酵母在YPD液体培养基中培养24 h后划线纯化2次,取单菌落甘油保藏。
将甘油保藏的酵母以2%接种量接种至YPD培养基中,30 ℃、180 r/min培养24 h作为种子液。取2%的种子液接种到新的YPD液体培养基中,作为第1代,每24 h接种1次为1代[12],在双功能气浴恒温摇床中180 r/min、30 ℃条件下培养。在第1、5、10、15、20代生长24 h后取酵母样液进行后续检测。
1.3.2 不同代数酵母的生长曲线测定
采用雷雅男等[23]使用的方法测定生长曲线。
1.3.3 发酵液电导率和核酸含量的测定
利用电导仪测定电导率。采用许艳俊等[24]使用的方法,用紫外分光光度计在260 nm和280 nm处测定发酵液的吸光度,采用M.wave Professional 2.0计算核酸的含量。
1.3.4 还原糖含量的测定
用DNS法测定还原糖的含量[25]。
1.3.5 乙醇含量的测定
取适当稀释的样液1.00 mL于比色管中,加入2.00 mL重铬酸钾溶液,用蒸馏水定容至10 mL,沸水浴10 min后,立即用流水冷却,用分光光度计在波长600 nm处测定其吸光度,利用乙醇含量标准曲线计算乙醇含量[26]。
1.3.6 甘油含量的测定
甘油含量的测定采用张永生等[27]的方法。
1.3.7 海藻糖含量的测定
海藻糖含量的测定参考Dong等[20]使用的方法,并优化。取5 mL菌液在4 000 r/min条件下离心5 min,弃去上清液。加入5 mL 0.5 mol/L三氯乙酸,然后放入冰水浴30 min(每5 min摇晃1次),取出后在4 000 r/min条件下离心10 min,取1 mL上清液于比色管中,加入5 mL蒽酮-硫酸试剂,沸水浴5 min后流水冷却至室温,590 nm处测定吸光度。
1.3.8 酵母胞内蛋白含量的测定
酵母胞内蛋白含量的测定采用Bradford法。
1.3.9 酵母胞内ROS含量和SOD、CAT活性的测定
分别取不同代酵母的菌液100 mL,5 000g离心10 min收集菌体。将收集到的菌体置于研钵中,加入适量液氮研磨至糊状。之后使用生理盐水定容至10 mL,混匀后在4 ℃、9 000g离心10 min。使用南京建成试剂盒E004-1-1、A001-1-1、A007-1-1分别对酵母胞内ROS含量以及SOD、CAT的活性进行测定。
1.4 数据处理
试验数据以平均值±标准误差表示,采用 SPSS 16.0 中Duncan多重比较的检验方法和T检验法对数据进行分析,并采用 Microsoft Excel 2016作图。
2 结果与分析
2.1 不同代酵母的生长曲线
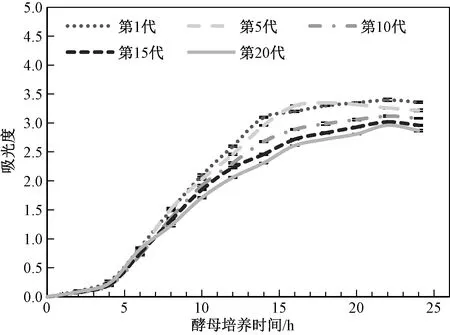
图1 不同代酵母生长曲线Fig.1 Growth curves of different generations of yeast
由图1可知,酵母的生长趋势随着代数的增加呈现出显著下降的趋势。在前4 h的迟滞期,不同代数酵母的生长没有显著的差异。在4~14 h酵母的对数生长期,不同代数酵母的生长开始出现差异,生长速度随着酵母传接代数的增加而减慢,第1代酵母在14 h时开始进入平稳期,而第5、10、15和20代酵母分别在14 h、16 h、20 h和22 h时进入平稳期,酵母进入平稳期,吸光度峰值随着酵母的代数的增加而降低。酵母在传代过程中的生长增殖能力逐渐下降。一方面可能是由于酵母的葡萄糖利用量减少,代谢下降所致;另一方面可能是由于随着传接代数的增加,酵母的自由基含量积累,细胞受到损伤,导致生长性能下降[13]。
2.2 不同代酵母发酵液中电导率、核酸和蛋白质含量的变化
由表1可知,酵母菌液的电导率随传代次数的增加呈现出显著升高的趋势,第20代的电导率显著高于其他处理组。说明酵母在传代过程中细胞膜的通透性可能受到影响。一方面可能是随着细胞老化,活性氧含量的增加,细胞膜受到损伤,导致老化的酵母细胞膜的功能衰弱,胞内物质外泄[28];另一方面可能是酵母在代谢减弱时,通过增加细胞膜的通透性,以吸取营养物质来抑制代谢活力的减弱[24]。
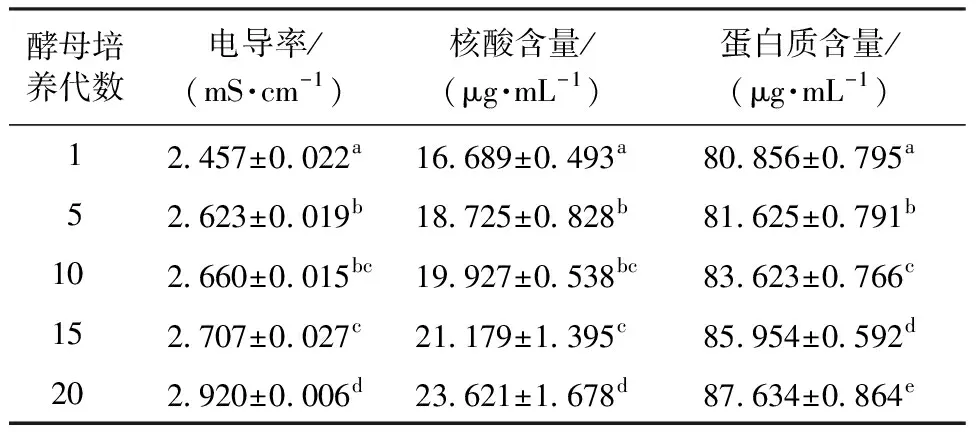
表1 不同传代次数酵母菌液的电导率、核酸和蛋白质含量Table 1 Conductivity, content of nucleic acid and protein of yeast medium with different times of passage
由表1可知,第20代酵母的胞外核酸含量最多且与15代之间的变化值最大,第5代显著大于第1代,第15代显著大于第5代,该结果同电导率结果相似。同时随着传代次数的增加发酵液中的蛋白质含量也呈显著增加的趋势。酵母在传代过程中,细胞内核酸和蛋白质等重要生命物质出现一定程度的外泄,并且在15—20代时最为严重。这可能是因为老化的酵母细胞膜功能减弱,酵母发生自溶,胞内的核酸及蛋白质外泄[29],或是因为在氧化胁迫条件下部分细胞损伤破裂。酵母培养基中蛋白质含量的增加也可能是因为酵母自身对老化的组织降解排出胞外的结果[30]。
2.3 不同代酵母培养基中葡萄糖、甘油、海藻糖、乙醇含量的变化
不同代酵母发酵液中剩余葡萄糖的含量如表2所示。第5代与第1代酵母发酵液中葡萄糖含量相比没有显著变化,第10代与第5代酵母发酵液中葡萄糖含量相比显著增多,第15代与第10代酵母发酵液中葡萄糖含量相比没有显著变化,而第20代发酵液中葡萄糖含量与第15代相比有显著增加并且差距最大。发酵液中残糖含量反映了酵母对培养基中葡萄糖的吸收利用能力,残糖含量越少说明酵母利用量越高。结果表明,酵母细胞传代次数的增加会影响酵母的碳利用率,细胞的衰老影响了酵母细胞的碳利用率,影响细胞的生长,该结果也同酵母的乙醇产量相一致。这可能是因为酵母在传代过程中自由基不断积累,导致高代数的酵母自身ROS含量较高,严重减弱了细胞膜物质转运的功能[13,30],削弱了细胞代谢的能力,导致酵母的碳代谢量降低。

表2 不同传代次数酵母碳代谢相关物质的含量Table 2 Content of substances related to carbon metabolism in yeast with different times of passage
由表2可知,第1代、第5代和第10代酵母随着传接代数的增加发酵液中甘油含量依次显著增加,第15代和第10代酵母相比没有显著差异,第20代与第15代相比显著增加,并且增加量是前3组组间差值的1.78~2.80倍,第20代的甘油含量是第1代的2.84倍。传代次数多的酵母细胞甘油产量增多,表明老化的细胞碳代谢更加偏向于甘油代谢,这可能与酵母受到高渗冷冻胁迫相似[20,31],在传代过程中受到外界胁迫导致酵母启动HOG应答机制,产出更多的甘油来保护自身细胞。
由表2可知,酵母胞内海藻糖的含量随着传接代数增加呈现显著升高的趋势。在第5—10代和第15—20代增幅较大,分别为35.673、64.015 μg/mL。传代次数多的酵母细胞中海藻糖含量的增多同甘油的结果相似,说明老化的酵母细胞的碳代谢倾向于甘油合成的同时也增加了海藻糖的合成强度,这可能是酵母对传代过程中受到不断积累的胁迫效应做出的反应应答的结果[16]。
由表2可知,第1代、第5代、第10代酵母发酵液中乙醇含量依次显著降低,第15代酵母发酵液中乙醇含量与第10代相比稍降低,第20代酵母发酵液中乙醇含量显著降低并且减少量是第1—15代每相邻实验组之间差值的2.36~6.80倍,第20代的乙醇含量也仅为第1代的30.27%。传代次数越多的酵母产出的乙醇越少说明酵母发酵力减弱[32]。所以,随着不断地传代衰老,酵母的碳代谢中糖酵解代谢能力下降,发酵力减弱,其中第20代最为严重。这可能与酵母利用的葡萄糖量相关[21],另外,根据表2中海藻糖和甘油含量的变化,推测酵母可能减少乙醇的代谢以产生更多的海藻糖和甘油,来抵御活性氧对细胞的损伤。
2.4 不同代酵母胞内ROS含量、胞内CAT、SOD的活性
如表3所示,523.6 nm处的荧光峰值体现酵母ROS荧光强度,酵母细胞内ROS含量随着酵母传代次数的增多而显著增加,其中第15、20代的酵母胞内ROS的荧光强度分别是第1代的2.29、2.81倍。酵母在受到胁迫时会产生氧化应激,导致线粒体中ROS含量增加,ROS的产生严重削减酵母的活力。ROS含量的增高进一步证明酵母在传代过程中所受到损伤胁迫的积累,同时也解释了酵母活性降低的原因,该结果同丁华建等[13]的研究结果相一致。
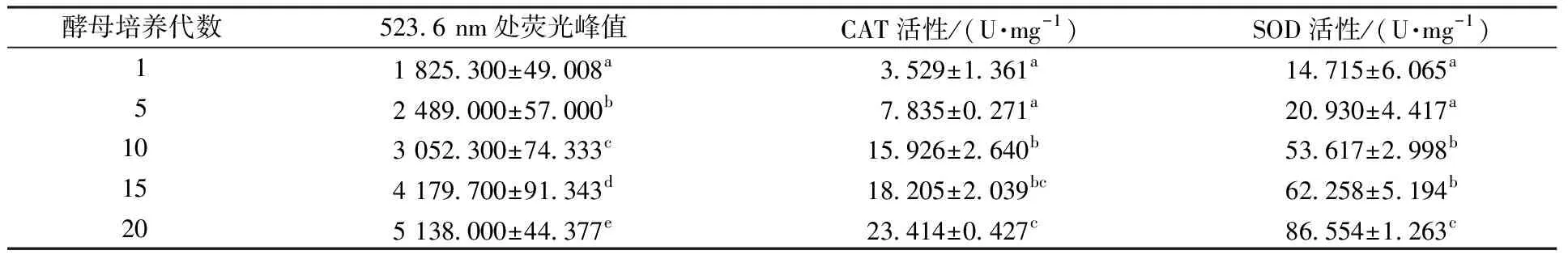
表3 不同代酵母的ROS含量及抗氧化酶活性Table 3 ROS content and antioxidant enzymes activity of different generations of yeast
由表3可知,酵母第1代和第5代、第10代和第15代之间胞内CAT和SOD的活性没有显著的差异,在第5代和第10代之间有显著的差异。第15代和第20代酵母胞内CAT活性没有显著差异,而SOD的活性显著增加。酵母细胞内的CAT、SOD都是抗氧化酶的一种,能够清除酵母细胞内的活性氧自由基,减少酵母的氧化损伤,这也是酵母抵抗衰老的表现[33]。酵母每间隔10代抗氧化酶有显著的增长,这同酵母胞内ROS含量的增加相一致,可能是酵母在氧化胁迫的情况下自身的应答,通过提高抗氧化酶的活性来降低氧化胁迫造成的伤害。而与此不同的是尹亚辉等[14]却发现酵母在传代过程中胞内抗氧化酶的活性先升高再降低,这可能是由于所选菌种和代数定义不同导致的酵母老化程度不同。抗氧化酶活性提升的同时,胞内的ROS含量依然在不断增加,这可能是酵母所受到的胁迫超过了自身的抵抗能力的范围。
3 结论
酵母在传代过程中不断老化,细胞膜的功能衰弱,对葡萄糖的利用率降低,发酵力减弱。在酵母的碳代谢量降低的前提下,细胞的代谢从糖酵解代谢转向海藻糖和甘油代谢。酵母胞内抗氧化酶活力的提升不能很好地抵抗酵母受到的氧化胁迫。酵母生长速率在第10代开始受到影响,并且在第5代、第10代和第20代时的发酵力分别显著下降。本研究初步探索了酵母在传代过程中的生长特性、糖代谢变化和抗氧化特性,下一步可结合酵母各基因的表达强度的进行更加深入的研究。
猜你喜欢
中国乳品工业(2022年4期)2022-05-19
生物化工(2021年6期)2022-01-27
中国兽医杂志(2019年10期)2019-05-20
能源(2017年7期)2018-01-19
中成药(2017年12期)2018-01-19
中学科技(2017年11期)2017-12-26
科学与财富(2016年32期)2017-03-04
中国民族医药杂志(2016年4期)2016-05-09
安徽医科大学学报(2015年9期)2015-12-16
中国兽药杂志(2014年5期)2014-05-29
