
普通小麦川麦60-簇毛麦 3V(3D)代换系的分子细胞学鉴定
2022-03-01 04:48郭元林邓光兵
麦类作物学报 2022年2期
张 洁,蒋 云,郭元林,王 颖,邓光兵,宣 朴,龙 海
(1.四川省农业科学院生物技术核技术研究所,四川成都 610061; 2.农业部西南地区小麦生物学与遗传育种重点实验室,四川成都 610066; 3.中国科学院成都生物研究所,四川成都 610041; 4.四川省农业科学院农产品加工研究所,四川成都 610066)
小麦条锈病是由小麦条锈菌(f.sp.)引起的一种常见的真菌性病害,具有变异程度高、流行程度广等特点,发生时可严重影响小麦品质和产量。因此,小麦条锈病是中国乃至全世界小麦生产上最具威胁的病害之一。培育抗病品种是防控小麦条锈病最经济、有效和对环境友好的策略。近年来,前人已从小麦及其近缘物种中鉴定到大量抗条锈病基因。但由于条锈生理小种与抗源的协同进化,许多过去大量使用的基因(如、、等)对目前流行的条锈生理小种已经失去了抗性。因此,亟需从小麦及其近缘物种中挖掘、筛选新的抗条锈基因,以满足小麦生产的需要。
普通小麦(L.,2=6=42,AABBDD)的近缘物种是小麦的重要基因源。将其与普通小麦进行杂交,可有效拓宽小麦的遗传背景,丰富小麦的遗传多样性。一年生二倍体簇毛麦(L.,2=2=14,VV)起源于地中海地区东北部,是小麦重要的三级基因源,携带有多种抗性基因,如锈病、眼斑病、白粉病、全蚀病以及耐盐胁迫等。其中,来自6VS染色体上的抗白粉病基因已被深入研究,并得到广泛利用。但对于簇毛麦条锈病抗性基因的挖掘和利用的研究尚未见报道。
本课题组前期通过杂交创制了一个对小麦条锈病具有优良抗性的普通小麦(中国春,CS)-簇毛麦3V(3D)二体代换系,命名为CD-3。通过对F代群体进行遗传分析,发现CD-3的条锈病抗性由一对隐性基因控制,且该基因位于簇毛麦3V染色体上,并将此基因命名为。川麦60(98-1231//贵农21/生核3295)分别于2011年和2012年完成国家和四川省审定,具有高产、高抗条锈病、综合性状较好等优点。但近年来,随着条锈菌生理小种和抗源的协同进化,川麦60的抗性已完全丧失。为进一步利用簇毛麦3V染色体上的条锈病抗性基因,创制培育出综合性状优良且可用于小麦遗传育种的抗性材料,本研究通过传统杂交手段,将3V染色体向川麦60背景中进行转移,在后代中筛选到一个高抗条锈病的株系,命名为1901-245,并从分子细胞学水平上对其进行精确鉴定和评价,以期为小麦条锈病抗性改良育种提供优良的资源材料。
1 材料与方法
1.1 试验材料
本研究使用的小麦栽培品种川麦60由四川省农业科学院作物研究所朱华忠研究员提供;CS-簇毛麦3V(3D)二体代换系(CD-3)由本实验室创制;CS-簇毛麦1V、2V、4V~7V附加系(#3)由电子科技大学杨足君教授提供;诱发材料Sy95-71、簇毛麦ZC6由本实验室保存;川麦60与CD-3回交自交后代1901-245(从以川麦60作回交父本的BCF代中筛选得到)由本实验室创制。抗条锈病鉴定所用的条锈病混合生理小种CYR32、CYR33和CYR34由甘肃省农业科学院植物保护研究所提供。
1.2 分子标记分析
取亲本CD-3和川麦60、回交自交后代1901-245以及对照材料中国春、簇毛麦ZC6、CS-簇毛麦1V、CS-族毛麦2V、CS-族毛麦4V~7V附加系的幼嫩叶片,使用CTAB法提取总基因组DNA。用Li等报道的簇毛麦3V染色体特异引物(表1)对供试材料进行检测,引物由上海生工生物工程公司合成。PCR反应体系为25 μL,包含Taq PCR mix预混液17.5 μL(生工生物工程公司,上海),基因组DNA (50~100 ng·μL) 1.8 μL,上下游引物(10 μmol·L)各 1.4 μL。配制好的PCR体系置于Biometra PCR仪(耶拿,德国)上进行扩增,PCR反应程序:94 ℃预变性5 min;94 ℃变性1 min,55 ℃退火1 min,72 ℃延伸2 min,共34个循环;最后72 ℃延伸10 min。扩增产物使用浓度为1%的琼脂糖进行凝胶电泳,EtBr染色后在BIO-RAD紫外凝胶成像系统 (加利佛尼亚,美国)下进行拍照。

表1 本研究所使用的3V特异引物Table 1 3V- specific primers used in the study
1.3 根尖细胞染色体FISH分析
待供试材料的根长至1~2 cm时剪取根尖,按照Zhang等的方法对根尖进行笑气(NO)预处理。中期染色体制片方法按照Kato等报道的方法进行。在寡聚核苷酸探针Oligo-pSc119.2和Oligo-pTa535的5′端分别连上荧光基团6-FAM和6-TAMRA,以识别小麦A、B、D组染色体身份;在簇毛麦特异探针Oligo-Hv62-1的5′端连上Cy5,以识别簇毛麦3V染色体,所有探针由上海捷运生物科技有限公司合成。FISH分析按照Fu等的报道进行。DAPI染色后使用莱卡DM2500荧光显微镜拍摄图片。每个材料调查3个中期分裂相以确定最终核型。
1.4 花粉母细胞减数分裂期染色体配对分析
待花粉母细胞处于减数分裂中期I时,收集1901-245的花药。用卡诺氏固定液无水乙醇∶氯仿∶冰醋酸(V∶V∶V=6∶3∶1)固定24 h后,使用95%的酒精冲洗花药,之后将花药转入70%的酒精中,-20 ℃保存。在载玻片上用45%的醋酸压片,随后在液氮中揭片。荧光原位杂交的探针选择、操作流程及图像获取同 1.3。每个材料观察30个花粉母细胞以分析染色体配对 情况。
1.5 条锈病抗性鉴定
将亲本CD-3、川麦60和10颗1901-245的种子种植于四川省农科院郫县试验基地。试验区块周围种上诱发行。连续两年(2019和2020年)对上述材料进行田间条锈病抗性鉴定。操作如下:在植株三叶期时,用牙签蘸取含条锈菌生理小种CYR32、CYR33和CYR34的混合菌种,并将其涂抹于供试材料和诱发材料的心叶上,随后适量喷浓度为0.01%的吐温20进行保湿。接种18~20 d后观察植株抗性反应。当诱发材料发病严重度达到80%以上时,对1901-245及其亲本CD-3和川麦60进行成株期抗性调查,记录其反应型。随后每隔10 d调查一次,一共调查3~4次。反应型(IT)按照Mcintosh等的报道分为0~4级,分别为0(免疫)、0;(近免疫)、1(高抗)、2(中抗)、3(中感)和4(高感)。
2 结果与分析
2.1 分子标记分析结果
用引物3V-7、3V-11、3V-14、3V-21和3V-24对供试材料CS、川麦60、簇毛麦ZC6、CD-3、1901-245以及CS-簇毛麦1V、2V、4V~7V附加系进行扩增。结果(图1)表明,这5对引物均能在ZC6、CD-3和1901-245中扩增出3V染色体的目标条带,而在CS、川麦60以及CS-簇毛麦1V、2V、4V~7V附加系中不能扩增出簇毛麦3V染色体的目标条带(图1)。另外,引物3V-21除了能扩增出3V染色体特异条带(图1d)外,还能在CS、川麦60以及CS-簇毛麦1V、2V、4V~7V附加系中扩增出小麦3D染色体的目标条带。
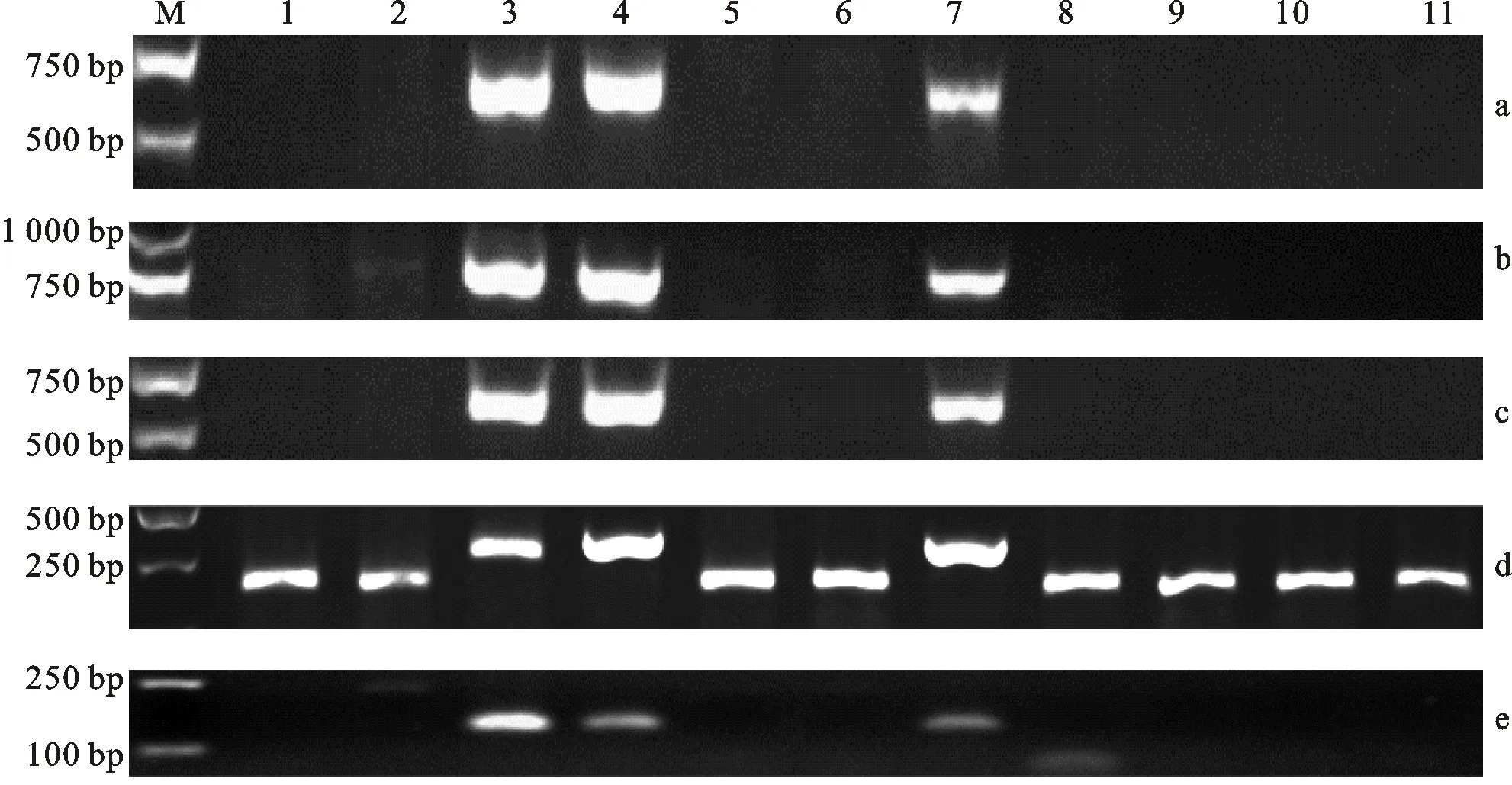
M:DNA marker Trans2K plus;1:中国春;2:川麦60;3:簇毛麦ZC6;4:CD-3;5:CS-簇毛麦1V附加系;6:CS-簇毛麦2V附加系;7:1901-245;8:CS-簇毛麦4V附加系;9:CS-簇毛麦5V附加系;10:CS-簇毛麦6V附加系;11:CS-簇毛麦7V附加系。a~e分别为用3V-7、3V-11、3V-14、3V-21和3V-24引物对供试材料的PCR扩增结果。M:DNA marker Trans2k plus; 1:Chinese Spring(CS); 2:Chuanmai 60; 3:D.villosum ZC6; 4:CD-3; 5:CS-D.villosum 1V additional line; 6:CS-D.villosum 2V additional line; 7:1901-245; 8:CS-D.villosum 4V additional line; 9:CS-D.villosum 5V additional line; 10:CS-D.villosum 6V additional line; 11:CS-D.villosum 7V additional line.a-e indicate PCR amplicons of the wheat materials amplified by specific primers 3V-7,3V-11,3V-14,3V-21 and 3V-24,respectively.图1 3V染色体特异引物对供试材料的PCR扩增结果Fig.1 PCR amplicons of wheat materials by 3V-specific primers
2.2 亲本染色体核型构建及1901-245的染色体鉴定结果
用寡聚核苷酸探针Oligo-pSc119.2、Oligo-pTa535和簇毛麦特异探针Oligo-Hv62-1构建CD-3(CS背景)、川麦60以及1901-245的标准核型,同时对1901-245的染色体构成进行鉴定。FISH核型分析显示,Oligo-pSc119.2和Oligo-pTa535可有效区分21对小麦染色体(图2a、图2c、图2d),Oligo-Hv62可在簇毛麦3V染色体长臂、短臂端部以及着丝粒区杂交出明显的簇毛麦特异信号(图2b、图2e)。由FISH鉴定结果可知,1901-245携带一对簇毛麦3V染色体,而丢失了一对小麦3D染色体(图2d~图2f)。另外,由图2f可知,CD-3中的小麦染色体和川麦60部分染色体之间存在FISH条带多态性,如在2A染色体上, CD-3的长臂端有较弱的Oligo-pSc119.2信号,而川麦60的长臂端则不具有Oligo-pSc119.2信号;在4A染色体上,CD-3的长臂端有强烈的Oligo-pSc119.2信号,而川麦60长臂端仅有大范围的Oligo-pTa535信号;在5A染色体上,CD-3的长臂中部有强烈的Oligo-pTa535信号,而川麦60的长臂中部Oligo-pTa535信号却很弱,同时还具有Oligo-pSc119.2信号;在3B染色体上,CD-3的短臂端Oligo-pSc119.2有强烈的信号,而川麦60的短臂端Oligo-pSc119.2信号却较弱(图2F)。根据这些多态性,可初步判定其后代1901-245,2A和3B染色体来源于川麦60,而4A和5A染色体来源于CD-3。另外,核型比较还发现,1901-245的2D染色体长臂与亲本CD-3和川麦60都存在差异。
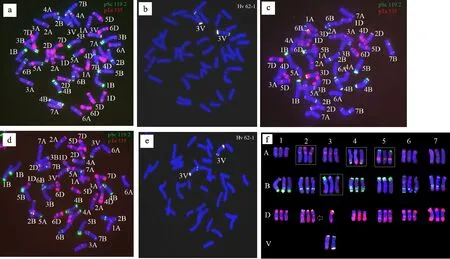
绿色、红色和白色分别为探针Oligo-pSc119.2、Oligo-pTa535和簇毛麦特异探针Oligo-Hv62-1信号。a~b:CD-3的 FISH鉴定图;c:川麦60的FISH鉴定图;d~e:1901-245的FISH鉴定图;f:CD-3、川麦60和1901-245的染色体FISH核型图。 图f中每一组三条染色体从左到右依次为CD-3、川麦60和1901-245,其中3V染色体为1901-245中的3V染色体,左为以Oligo-pSc119.2和Oligo-pTa535为探针的核型,右为簇毛麦探针Oligo-Hv62-1的核型。方框所示为亲本间具有多态性的染色体,箭头所示为1901-245 2D染色体与亲本2D染色体的多态性。Green signals,red signals and white signals were probes Oligo-pSc119.2,Oligo-pTa535 and Oligo-Hv62,respectively.a-b:Image of chromosomes in CD-3 analyzed by FISH; c:Image of chromosomes in Chuanmai 60 analyzed by FISH; d-e:Image of chromosomes in 1901-245 analyzed by FISH; f:FISH karytypes of CD-3,Chuanmai 60 and 1901-245,and in every group,chromosomes from CD-3,Chuanmai 60 and 1901-245 were shown from left to right,respectively.Additionally,3V chromosome is originated from 1901-245 (The left is the chromosomal karyotype with probes Oligo-pSc119.2 and Oligo-pTa535,and the right is the chromosomal karyotype with V genome-specific probe of Oligo-Hv62-1).Rectangles indicate the obvious different karyotypes of the chromosomes,originated from its parents CD-3 and Chuanmai 60,respectively.Arrow indicates the different karyotype of 2D chromosome originated from 1901-245,compared with its parents CD-3 and Chuanmai 60.图2 CD-3、川麦60和1901-245的FISH 分析图及标准核型图Fig.2 FISH analysis and karyotype of CD-3,Chuanmai 60 and 1901-245
2.3 花粉母细胞染色体减数分裂期的行为
用寡聚核苷酸探针Oligo-pSc119.2、Oligo-pTa535和簇毛麦特异探针Oligo-Hv62-1对1901-245花药的37个花粉母细胞进行染色体行为统计分析,发现1901-245花粉母细胞的染色体总数均为42,平均含有单价体0.11个,环状二价体19.73个,棒状二价体1.22个,未发现多价体。因此,1901-245花粉母细胞染色体平均构型为:2=42=0.11I+(19.73环+1.22棒)Ⅱ。出现单价体的染色体分别为3B、4B和5A,未发现簇毛麦3V染色体出现单价体的情况(图3a~3f)。另外,在减数分裂后期I也未发现落后染色体或染色体桥的出现(图3g、3h)。
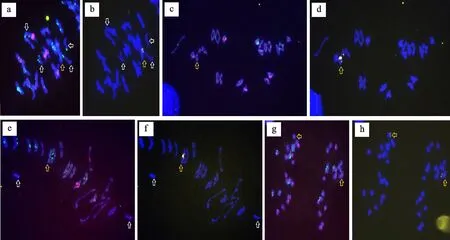
a~f:减数分裂中期I;g-h:减数分裂后期I。白色箭头所示为单价体,黄色箭头所示为簇毛麦3V染色体。a-f:Chromosome behaviors at meiotic metaphase I; g-h:Chromosome behaviors at meiotic telophase I.White arrows indicate the univalent,and yellow arrows indicate the 3V chromosomes of D.villosum.图3 1901-245的花粉母细胞减数分裂期染色体行为Fig.3 Chromosome behaviors in meiosis of 1901-245
2.4 田间条锈病抗性鉴定及植株表型观察结果
用条锈病混合生理小种(含CYR32、CYR33 和CYR34)对诱发材料Sy95-71、簇毛麦ZC6、CS、亲本川麦60和CD-3以及1901-245进行人工接种,并连续两年对其进行田间条锈病抗性鉴定,结果(图4c)表明,诱发材料Sy95-71和亲本川麦60对条锈病混合生理小种表现为高感(IT=4);CS表现为中感(IT=3);CD-3和1901-245表现为高抗(IT=1);簇毛麦ZC6表现为近免疫(IT=0;)。对植株形态进行观察发现,1901-245的株高与川麦60相似,但其植株形态和穗型更接近于CD-3(图4a、图4b)。对CD-3、1901-245和川麦60的籽粒大小进行调查发现,1901-245的粒宽和粒长都与川麦60更相近(图4d、图4e)。

CM60:川麦60。a:植株形态图;b:穗型图;c:叶片抗性反应图;d和e:籽粒形状。CM60:Chuanmai 60.a:Plant morphology; b:Spike shape; c:Leaf response to stripe rust; d and e:Seed shape.图4 1901-245及其亲本CD-3和川麦60的表型Fig.4 Phenotype of 1901-245 and its parents CD-3 and Chuanmai 60
3 讨 论
3.1 一年生二倍体簇毛麦中抗病基因的研究 应用
一年生二倍体簇毛麦因具有抗白粉病、锈病、眼斑病、全蚀病等多种小麦病害抗性,而被广泛应用于小麦抗性改良中。近年来有少量关于簇毛麦携带条锈抗性的报道,但对于簇毛麦3V染色体上条锈病抗性基因的挖掘与利用的研究尚未见有报道。本课题组前期研究发现,CD-3的条锈病抗性由一对隐性基因控制,且该基因位于簇毛麦3V染色体上,为进一步利用簇毛麦的条锈病抗性基因奠定基础。
3.2 1901-245及其亲本CD-3和川麦60染色体FISH核型的多态性
重复序列在染色体上的分布在一定程度上可以反映植物染色体组的结构以及基因组进化过程。目前,前人已基于重复序列开发出大量寡聚核苷酸探针。这些探针可用于构建小麦近缘物种的染色体核型,以识别小麦背景下的外源染色体(片段)。为准确鉴定1901-245的染色体身份,我们构建了基于寡聚核苷酸探针Oligo-pSc119.2和Oligo-pTa535的普通小麦川麦60以及CD-3(CS背景)的FISH核型,并使用簇毛麦特异探针Oligo-Hv62-1来特异追踪来自簇毛麦的3V染色体。由FISH核型鉴定结果可知,寡聚核苷酸探针Oligo-pSc119.2和Oligo-pTa535联合使用,可有效区分六倍体小麦A、B、D组的42条染色体,与Tang等报道的结果一致。Oligo-Hv62-1可在3V染色体臂端杂交出明显的簇毛麦特异信号,与Li 等报道的结果一致。通过比较CD-3和川麦60的FISH核型发现,两者在2A、4A、5A和3B染色体上存在FISH核型多态性。前人对小麦染色体FISH核型存在多态性已有较多报道。在本研究中,CD-3与川麦60的FISH核型多态性应该与栽培种培育过程中的人工选择有关。通过比较1901-245与两个亲本CD-3和川麦60的FISH核型,可初步判定1901-245的2A和3B染色体来源于川麦60,而4A和5A染色体来源于CD-3。这表明1901-245的创制过程中,川麦60的染色体并未完全替代CD-3的染色体,这提示我们应将1901-245继续与川麦60进行回交。另外,1901-245的2D染色体与亲本CD-3和川麦60的2D染色体长臂端均存在显著差异,这表明在杂交过程中2D染色体可能进行了重排。在后期研究中,我们将持续关注2D染色体长臂的变异,并进一步利用分子标记和基因组荧光原位杂交技术判定2D染色体长臂结构的变异情况。
3.3 1901-245的田间条锈病抗性及其应用
本研究分子细胞学鉴定结果表明,1901-245材料携带一对簇毛麦3V染色体,但丢失了一对小麦3D染色体,说明1901-245为普通小麦川麦60-簇毛麦3V(3D)代换系。对其花粉母细胞染色体行为进行分析发现,该材料染色体配对正常,来自簇毛麦的3V染色体未出现单价体,表明1901-245遗传较为稳定。另外,对其农艺性状和条锈病抗性调查结果显示,1901-245籽粒较大,株高较矮,与川麦60相似;条锈病抗性表现为高抗,与CD-3一致,表明1901-245已具有部分川麦60的优良性状,且3V染色体上的条锈病抗性基因在1901-245的遗传背景下也能正常表达。这一点说明,来自于簇毛麦3V染色体上的条锈病抗性基因可向其他非CS背景的小麦中进行转移且保持其原有抗性,应进一步得到应用。
CS作为地方小麦品种,具有较多的不良性状,比如植株高、易倒伏、株型差、千粒重低等,因此,不易对其进行遗传改良。本研究中利用的CD-3是以CS为背景创制的高抗小麦条锈病的CS-簇毛麦3V(3D)代换系,但株型、株高、穗型、千粒重等主要农艺性状均与中国春一致。因此,在对CD-3的条锈病抗性利用上具有一定难度。本研究经过将CD-3与四川推广小麦品种川麦60进行杂交并回交,经过两代自交后得到了一个抗性稳定的高抗小麦条锈病株系1901-245。该材料在株高、粒长和粒宽上相较于CS也有了较大改善。因此,1901-245是宝贵的高抗小麦条锈病的资源材料,可在小麦抗性改良中进行深入研究与利用。鉴于其仍表现出部分CS性状(如穗型、株型),后期我们将继续用1901-245与川麦60进行回交,并加强分子细胞学的追踪,以确保簇毛麦3V染色体稳定遗传。
猜你喜欢
农民致富之友(2020年22期)2020-09-02
三农资讯半月报(2020年13期)2020-07-31
农民致富之友(2020年12期)2020-05-11
当代化工(2020年2期)2020-03-18
农民致富之友(2019年20期)2019-07-27
农民致富之友(2019年16期)2019-07-01
农民致富之友(2017年10期)2017-06-15
江苏农业科学(2016年4期)2016-06-14
卷宗(2014年7期)2014-08-27
卷宗(2014年1期)2014-03-20
