
小麦Ta-miR154的特征及渗透胁迫响应
2022-03-01 04:45王苗苗袁晓波王晓腾李永春
麦类作物学报 2022年2期
王苗苗,袁晓波,李 静,王晓腾,张 莉,李永春
(河南农业大学农学院/省部共建小麦玉米作物学国家重点实验室,河南郑州 450046)
干旱、高盐等渗透胁迫是限制小麦高产稳产的主要逆境因子,探讨小麦渗透胁迫响应的生理和分子机制对于开展小麦抗逆分子育种具有重要意义。近年来大量研究表明,microRNAs(miRNAs)作为一类非编码小RNA(长度在21~24 nt之间),参与植物生长发育调控、逆境胁迫响应等多个生理调节过程,有关miRNA参与的逆境胁迫响应调控机制已成为植物抗逆分子机理研究的重要领域,特别是miRNA作为抗逆分子育种的重要靶标也日益受到重视。对模式植物拟南芥的研究发现,miR169参与干旱响应的分子调节过程,过表达转基因材料的耐盐能力显著提高。说明miRNA在植物的非生物胁迫应答中具有重要的调控作用。对其他作物研究发现,miR172和miR396与马铃薯的脯氨酸合成调控及干旱胁迫响应密切相关,推测这两个miRNA可能参与了脯氨酸合成过程中关键基因的表达调控;过表达转基因水稻的耐盐能力降低,推测该miRNA参与了气孔发育及盐胁迫响应的调控过程;玉米miR169也与干旱高盐胁迫调节密切相关。在小麦中,Shi等发现,miR1119参与了抗旱调控过程;Bai等发现,miR408对于耐盐生理调节至关重要。本课题组前期研究发现,小麦在种子脱水过程中迅速上调表达;在此基础上,本研究进一步分析了的序列特征及其对干旱和高盐胁迫的响应模式,以期为探讨的功能及其在小麦育种中的应用提供参考。
1 材料与方法
1.1 试验材料与设计
试验所用小麦(L.)品种包括中国春和矮抗58,种植于河南农业大学原阳试验基地。种子形成过程中,剥取授粉后第5、10、15、20、25和30 d的籽粒,用于分析小麦的特征;取三叶期的根和叶片,开花期的节间、节、穗下节、旗叶,以及成熟期的籽粒,用于组织表达特性分析。胁迫处理过程如下:首先,选取饱满一致的小麦种子用NaClO(10%)表面消毒;然后,在人工气候室[温度为24 ℃(光照)和18 ℃(黑暗),每天光照16 h,光照强度240 mmol·m·s]水培至两叶一心期时进行胁迫处理。模拟干旱胁迫采用PEG6000(20%)处理,模拟盐胁迫采用NaCl溶液(150 mmol·L)处理,分别剪取胁迫处理0、0.5、1、2、6、12、24、48 h的根和叶,用于分析胁迫下的表达量。上述试验均设3个生物学重复,所有组织用液氮速冻置于-80 ℃保存、备用。
1.2 RNA和DNA的提取
籽粒总RNA采用Trizol Plant试剂盒(全式金,北京)提取,其余组织总RNA采用RNAisoPlus试剂盒(Tarkara,日本);RNA反转录采用TransScript miRNA First-strand cDNA Synthesis Supermix试剂盒(全式金,北京),合成的cDNA于-80 ℃保存,备用;基因组DNA采用CTAB法提取。
1.3 基因片段的克隆及序列分析
用小麦基因组数据库Ensembl Plants(http://plants.ensembl.org/Triticum_aestivum/Info/Index)对小麦miR154序列(5′-GGCGAGGGACAUACACUGUACA-3′)进行比对分析,确定基因位点。候选基因序列下载后利用SnapGene软件进行3个部分同源基因序列比对分析,用RNA Folding Form(http://unafold.rna.albany.edu/?q=mfold/rna-folding-form)对RNA的发卡结构进行分析,用Primer 5.0软件设计引物。克隆的特异上游引物为YP2093(5′-ATTGTGAACTCCAAGTGGCATG-3′),下游引物为YP2095(5′-GGTTGTGTTGCTGTGCTCGATC-3′),用高保真酶LA Taq以基因组DNA为模板进行PCR扩增,PCR扩增体系为20 μL,包括10×LA PCR Buffer Ⅱ 2.0 μL,dNTP mix solution 1.8 μL,TaKaRa LA Taq Enzyme 0.2 μL,Template 1.0 μL,YP2093和YP2095引物(10 μmol·L)各1.0 μL,ddHO 13 μL。PCR扩增程序:95 ℃预变性5 min;95 ℃变性30 s,60 ℃退火1 min 30 s,72 ℃延伸25 s,30个循环;72 ℃延伸10 min, 4 ℃保存。克隆片段连接到pMD19-T载体后转化大肠杆菌DH5α,用载体通用引物YP0085(5′-ATCGGTGCGGGCCTCTT-3′)和YP0086(5′-GGCACCCCAGGCTTTACAC-3′)进行重组克隆,每个品种随机挑选5个克隆进行测序分析。
1.4 表达模式分析
用RT-qPCR的方法进行实时定量表达分析,反应体系为20 μL,包括cDNA模板1 μL,2×TransStart Tip Green qPCR SuperMix 10 μL,uni prime (10 μmol·L) 0.4 μL,specific prime (10 μmol·L) 0.4 μL,RNase-free水 8.2 μL。其中,uni primer为全式金试剂盒提供的通用引物,specific primer为YP0949(5′-GGCGAGGGACATACACTGTACA-3′)。做荧光定量时,使用试剂盒中的通用引物是由于miRNA的特殊性,miRNA序列很短,miR154成熟序列仅为22 nt,所以在反转录时进行了Ploy(A)加尾,在测定的表达量时,上游使用特异引物YP0949(5′-GGCGAGGGACATACACTGTACA-3′),下游使用通用引物。荧光定量PCR仪型号为Bio-Rad伯乐CFX Connect。每个反应设定3次重复,采用两步法PCR,反应程序: 94 ℃预变性30 s,94 ℃变性5 s,60 ℃退火30 s,40个循环。内标为snRNA U6,引物为YP1303(5′-CCTTCGGGGACATCCGATAAA-3′)和YP1304(5′- ATTTGGACCATTTCTCGATTTGTGC-3′)。实时定量数据分析采用2-△△法计算,并用Excel 2003作图。
2 结果与分析
2.1 小麦Ta-miR154的特征分析
用Ensembl Plants对miR154序列进行比对分析,发现基因位于第7同源群染色体上,将其在A、B、D基因组上的3个部分同源基因分别命名为、和。采用特异引物YP2093和YP2095对中国春和矮抗58两个小麦品种的基因组DNA进行扩增,两个品种均获得了约1 000 bp和1 500 bp的两个片段(图1A)。将上述两个片段分别连接到PMD19-T载体,连接产物转化大肠杆菌后,每个片段随机挑选5个重组克隆,利用A、B、D基因组特异引物进行鉴定并测序。测序结果显示,中国春三个部分同源基因、和间的序列存在较大差异。与(993 bp)相比,(1 011 bp)和(1 529 bp)分别存在70(63个SNP和7个InDel)和55(50个SNP和5个InDel)个差异位点;矮抗58和中国春两个品种间的miRNA前体区域序列高度保守,其中、、和分别存在12、15和5个SNP差异;在生成的茎环结构区域两个品种间序列完全一致(图1B),和分别仅存在1个SNP差异,而与另两个部分同源基因间存在4个SNP差异,特别是第32位的C/T变异使得其无法生成。RNA发卡结构分析显示,3个部分同源基因产生的miRNA前体均可形成典型的茎环发夹结构,但由于和间2个单核苷酸的差异导致发夹结构顶端的2个折叠环大小存在一定差异(图1C)。

A: Ta-MIR154的克隆结果,M: DL2000;1:中国春;2:矮抗58。B: Ta-MIR154三个部分同源基因间的序列比对。C:RNA二级结构分析,红色圆圈标记区域为成熟miRNA对应区域。A: Cloning results of Ta-MIR154, M: DL2000; 1: Chinese Spring; 2: Aikang 58.B: Sequence alignment among the three homologues of Ta-MIR154 genes.C: Secondary RNA structures of Ta-MIR154 gene,and sequences marked with red circles are regions corresponding to the mature miRNAs.图1 小麦 Ta-MIR154基因的克隆及序列特征分析Fig.1 Cloning and sequence characterization of Ta-MIR154 gene in wheat
2.2 小麦不同组织中 Ta-MIR154的表达模式 分析
对的组织表达特性进行分析,结果(图2A)发现,不同组织的表达模式存在明显差异。总体来看,在成熟种子中的表达量最高,其次为节间和根系组织,而在穗下节、节和叶片中的表达量较低;在两个品种间的表达水平呈现相似的变化趋势,但矮抗58的表达水平略高于中国春。进一步分析两品种籽粒发育过程中的表达模式,发现在籽粒发育早期的表达量均较低,而在授粉后20 d表达量迅速上升,授粉后30 d表达量达到最大值。
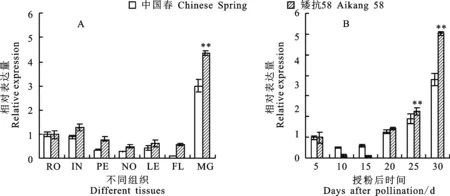
A: 不同组织中 Ta-MIR154的表达分析;B:籽粒形成过程中 Ta-MIR154的表达分析。RO:根系;IN:节间;PE:穗下节;NO:节;LE:叶;FL:旗叶;MG:成熟籽粒。图柱上的**表示品种间差异极显著(P<0.01)。A: Expression of Ta-MIR154 in different wheat tissues;B:Expression of Ta-MIR154 during grain development.RO:Root;IN:Internode; PE: Peduncle;NO:Node;LE:Leaf;FL:Flag leaf;MG:Mature grain.** on the columns indicate significant difference between varieties at 0.01 level.图2 小麦不同组织中 Ta-MIR154的相对表达量Fig.2 Expression of Ta-MIR154 in different wheat tissues
2.3 干旱胁迫对小麦 Ta-MIR154相对表达量的影响
为探讨对干旱胁迫的响应模式,利用qRT-PCR技术分析PEG6000(20%)处理条件下小麦叶片和根系中的表达模式。结果(图3)显示,随着干旱胁迫时间的延长,中国春小麦叶片中的相对表达量总体上呈上升趋势,特别是胁迫6 h后,上调表达幅度逐渐增加,胁迫24和48 h时的相对表达量分别约为胁迫0 h的6倍和14倍,差异均达极显著水平;矮抗58叶片中的相对表达量受胁迫诱导后快速上升,且整体上其表达水平要高于中国春(图3A),胁迫2、24和48 h的表达量与0 h之间差异均达极显著水平,推测在叶片中的积累与其抗旱调节能力的增强有关。随着干旱胁迫时间的延长,中国春根系中的相对表达量呈先上升后下降的变化趋势,胁迫12 h时的相对表达量最高,约为胁迫0 h的8倍,胁迫12和24 h时的表达量与0 h之间差异均达极显著水平;而矮抗58根系中的相对表达量呈先降低后上升的变化趋势,且整体表达水平低于中国春(除胁迫48 h外)(图3B),胁迫48 h的表达量与0 h之间差异达极显著水平。在所检测的两个品种间呈现出不同的干旱响应模式,可能与品种的抗旱特性差异有关。

A: 叶片中 Ta-MIR154的表达分析;B:根系中 Ta-MIR154的表达分析。**表示该品种 Ta-MIR154的相对表达量在该胁迫时间与0 h之间差异达极显著水平(P<0.01)。图4同。A: Expression of Ta-MIR154 in leaves; B:Expression of Ta-MIR154 in roots.** indicates significant difference of the expression of Ta-MIR154 in the variety between the corresponding stress time and 0 h(P<0.01).The same in figure 4.图3 干旱胁迫条件下 Ta-MIR154的表达分析Fig.3 Expression of Ta-MIR154 under drought stress
2.4 盐胁迫对小麦 Ta-MIR154相对表达量的 影响
盐胁迫条件下,两个小麦品种间叶片和根系中的表达模式均存在较大差异(图4A)。随盐胁迫时间的延长,中国春叶片中的相对表达量呈先上升后下降的变化趋势,胁迫24 h时的表达量最高,约为胁迫0 h的17倍,胁迫2~24 h的表达量与0 h之间差异均达极显著水平;而矮抗58叶片中的相对表达量在各胁迫时间与0 h之间均无显著差异。中国春根系中的相对表达量在各胁迫时间与0 h之间也均无显著差异;而矮抗58根系中的相对表达量呈升-降-升的变化趋势,胁迫2 h时的表达量最高,约为胁迫0 h的12倍,差异达极显著水平(图4B)。推测在小麦响应盐胁迫过程中发挥一定的功能,且其参与的盐胁迫调控机制在品种间存在一定差异。

图4 盐胁迫条件下 Ta-MIR154的表达分析Fig.4 Expression of Ta-MIR154 under salt stress
3 讨 论
小麦是典型的异源六倍体植物,含有A、B和D部分同源的3个基因组,大多数基因在小麦基因组中均存在3个部分同源的基因成员。因此,小麦基因组中广泛存在基因冗余现象,而且部分同源基因间往往具有功能互补的特性。本研究中,、和基因序列间存在一定差异,特别是在成熟miRNA对应位置由于一个核苷酸的差异导致其不能生成;而和在miRNA前体区域高度相似,而且miRNA前体序列都可形成典型的茎环结构,推测和两个基因均具有生成的能力。不过,由于和两个基因在miRNA前体区域存在两个碱基的差异,且导致RNA折叠后顶端两个环状结构发生微小的变化,这是否会影响到成熟的生成,仍需进一步证实。
植物在抵御逆境胁迫过程中涉及到一系列的生理响应和基因表达调控过程,已有研究表明,依赖于miRNA表达的基因在植物对逆境胁迫响应过程中发挥重要调控功能,Liu等研究还发现,依赖于miRNA的调控机制可能与小麦的胁迫记忆和跨代效应有关。本研究发现,对干旱和高盐胁迫的响应模式在两个小麦品种中不同,这可能与两个品种具有不同的抗逆特性有关。对基因组序列分析也发现,该基因上游调控区包含有防御和应激反应元件(TC-rich repeats)、脱落酸响应元件(ABRE)和胁迫诱导转录因子MYB,进一步说明参与渗透胁迫响应过程。另外,小麦籽粒成熟后期实际上也是一个干物质积累和籽粒脱水的调控过程,小麦籽粒脱水特性也直接关系到籽粒机收效率和安全贮藏问题;本研究发现,在籽粒成熟期迅速上调表达,推测其可能与籽粒脱水调控相关。在渗透胁迫响应过程中的调控功能仍有待进一步研究。
猜你喜欢
今日农业(2022年1期)2022-11-16
今日农业(2022年2期)2022-11-16
广西植物(2022年8期)2022-09-07
中国农学通报(2022年12期)2022-06-01
中国农学通报(2022年13期)2022-05-31
作文·小学低年级(2021年10期)2021-01-25
中学生物学(2019年7期)2019-10-17
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
科学种养(2017年6期)2017-06-13
科技资讯(2016年32期)2017-03-31
