
木质纤维生物质半纤维素分子模拟应用研究进展
2022-05-21 06:12林琦璇刘昕昕李理波任俊莉
生物质化学工程 2022年3期
林琦璇, 刘昕昕, 李理波, 彭 锋, 任俊莉*
(1.华南理工大学 轻工科学与工程学院,制浆造纸工程国家重点实验室,广东 广州 510640; 2.华南理工大学化学与化工学院,广东 广州 510640; 3.北京林业大学 材料科学与技术学院,北京 100083)
近年来,清洁生产和可持续发展面临的压力越来越大,可再生的木质纤维生物质资源作为一种化石资源的替代和补充,其重要性日益凸显[1-2]。木质纤维生物质中主要含纤维素、半纤维素和木质素,可用于制备生物能源、化学品及材料。目前相关研究主要基于实验,随着研究的深入,研究者们希望从分子层面了解物质的物理和化学性质。为此,研究者们尝试结合分子模拟技术来强化对生物质利用的研究。分子模拟通常可分为两大类:理论计算和经验计算。理论计算主要指量子力学模拟,经验计算主要指分子力学模拟。量子力学计算方法包括分子轨道理论方法、从头算方法、密度泛函理论方法。分子力学主要包括分子动力学、蒙特卡洛、分子力学、布朗动力学、介观动力学、耗散动力学等[3-4]。目前分子模拟在纤维素、木质素和半纤维素方面有初步的探索,并给出了有关的理论支撑。本文主要介绍有关半纤维素的分子模拟,包括常用的分子模拟软件、半纤维素在细胞壁中的物理形态、半纤维素转化、半纤维素基材料等,并对分子模拟在半纤维素研究的发展应用进行了展望,以期为半纤维素的进一步研究和利用提供指导。
1 分子模拟软件介绍
为了使运算更加高效,人们开发出适合于前沿硬件技术的全新动力学策略和代码,编写出一系列并行效率出色的分子动力学模拟软件。目前在生物质模拟过程中使用最广泛的主要有CHARMM、ADF、AMBER、CPMD、HyperChem、GROMACS、Materials Studio、Gaussian等模拟软件。
1.1 CHARMM
CHARMM(Chemistry at HARvard Macromolecular Mechanics)是一种用于分子动力学的分子力场,同时,采用这种力场的分子动力学软件包也采用了这个名称。CHARMM是一个被广泛承认并应用的分子动力学模拟程序,其主要用于生物学分子的模拟,包括蛋白质、多肽、脂类、核酸、碳水化合物和小分子配体。该软件提供了大量的计算工具,包括多种构象和路径采样方法、自由能估计器、分子最小化、动力学和分析技术以及模型构建能力。CHARMM可以使用多种不同的能量函数和模型来执行,包括混合量子力学-分子机械力场、具有显式溶剂和各种边界条件的全原子经典势能函数、隐式溶剂和膜模型[5]。
1.2 ADF
ADF(Amsterdam Density Functional)程序是在70年代初开发的,当时名为HFS(Hartree-Fock-Slater),在后续的不断补充和改进过程中,阿姆斯特丹和卡尔加里的理论化学研究小组做出了突出的贡献,目前ADF已经发展成为一个先进的量子化学程序包[6]。ADF程序包可安装在Linux、Windows、Mac OS X和一些Unix平台中。ADF可以计算基态和激发态能量、谐波振动频率,还可以优化结构、处理溶剂化问题、进行过渡态搜索和分析分子性质(如NMR自旋耦合或NMR自旋耦合)。所使用的方法主要是DFT,同时也允许用户使用QM/MM方法来处理更大的系统[7]。
1.3 AMBER
AMBER(Assisted Model Building with Energy Refinement)通常包含两个部分:一个是用于模拟生物分子的一组分子力场,为经验力场;另一个是由美国加州大学开发并维护的力场和代码独立的分子模拟软件。Amber软件包由Amber Tools和Amber两个部分组成,适用于蛋白质、核酸、脂类和糖类等生物分子的模拟。与同类软件相比,Amber具有一些特有的优势:1) Amber 软件功能强大,模拟物质种类众多,除常规分子动力学软件可模拟的蛋白质、脂质、核酸分子外还可进行糖类及小分子物质的模拟;2) Amber软件包中提供生成小分子力场参数文件的程序,从而保证Amber对小分子物质模拟的正确性和精确性[8]。
1.4 CPMD
CPMD(Car-Parrinello Molecular Dynamics)是由Car和Parrinello提出的从头算分子动力学方法[9],该方法利用了赝势、平面波基矢和密度泛函理论(DFT)对原子(分子)间作用力进行计算,在CPMD官网可以免费下载CPMD程序进行使用。与传统第一原理的计算方法相比,CPMD方法可以同时处理离子和电子的系统,不需要在每一步中重复自洽求解KS方程组,就能同时获得离子轨迹与相应电子基态,减少很多计算量[10]。
1.5 HyperChem
HyperChem软件包是HyperCube公司开发的32位Windows界面程序,是常用的分子设计和模拟软件,质量高、灵活易操作,可进行量子力学、分子力学、分子动力学计算。HyperChem软件包功能非常强大,主要由这样几个模块组成:① 构造分子;② 优化分子结构;③ 研究分子反应;④ 观察轨道和电子图谱;⑤ 评估化学反应路径和化工机械装置;⑥ 研究分子动力学[11-12]。HyperChem的主要功能:1) 强大的量子化学、分子力学计算功能;2) 可研究分子的HOMO-LUMO能隙、电离势、电子亲和力、偶极矩、电极化率、电子能级等各项特性;3) 构造分子模型。
1.6 GROMACS
GROMACS是一个分子动力学软件,它最初是在荷兰格罗宁根大学生物物理化学系开发的,现在由全球大学和研究中心的贡献者维护。GROMACS是一个缩写词,源自于格罗宁根化学模拟机器(GROningen MAchine for Chemical Simulations)。GROMACS是可用的最快和最流行的软件之一,可以在中央处理器(CPU)和图形处理器(GPU)上运行。它是基于GNU通用公共许可证(GPL)下发布的免费开源软件。从GROMACS 4.6版本开始,GROMACS开始基于GNU宽通用公共许可证(LGPL)下发布[13]。GROMACS具有免费性、迅速性、灵活性等优点。
1.7 Materials Studio
Materials Studio可以进行分子动力学模拟、蒙特卡洛模拟以及量子力学模拟,是一个多功能的软件。Materials Studio是由美国Accelrys公司开发的专门用于材料领域的模拟软件。Materials Studio最大的优势在于集建立三维模型、结构优化、计算模拟和结果分析功能于一体。Materials Studio具有大家熟悉的Microsoft标准用户界面,能应用多种数值算法以及多种算法综合应用,无论是进行简单的结构优化,还是做复杂的量子力学和动力学计算,都可以通过简单的操作来获得可靠的数据,是一个强有力的模拟工具[14]。
1.8 Gaussian
Gaussian是一个功能强大的商业量子化学计算软件包,不具有免费性,适用于多个系统平台(Windows、Mac、Linux、Unix),最初是由John Pople在1970年发布,目前最新的版本是Gaussian16。其主要功能为对分子结构和能量、过渡态的能量和结构、化学键和反应能量、分子轨道、原子电荷和电势、红外和拉曼光谱等的计算[15]。其计算结果通过Gauss View可视化软件配合使用,在有机化学、结构化学、物理化学、生物化学等领域获得广泛的应用[16]。
2 半纤维素大分子形态及其与纤维素结合方式理论模拟
2.1 半纤维素的构象解析
在植物细胞壁中,半纤维素与纤维素、木质素之间紧密连接,存在多种连接方式[17]。目前研究者主要集中于模拟半纤维素大分子形态及其与纤维素结合方式,这些研究能够为提取利用植物三大素提供理论依据。
半纤维素包括木聚糖和木葡聚糖等。影响半纤维素大分子构象的最重要因素是主链的柔韧性,这在很大程度上取决于糖苷键的构象自由度。有研究表明,主链不同组成的半纤维素多糖具有不同的构象,其侧链基团及周围环境也会对其构象造成影响[18]。
木聚糖是植物中最常见的半纤维素,侧链基团含有阿拉伯糖、葡萄糖醛酸、乙酰基中的一种或多种。Martinez-Abad等[19]利用GROMACS软件模拟研究发现,木聚糖在水溶液中的首选构象是31螺旋构象。木聚糖与纤维素相互作用时会发生构象变化,由灵活的31螺旋构象转变为平面的、相对刚性的21构象。这一变化直观地促进了木聚糖与平坦的纤维素表面的紧密相互作用,甚至可以达到共结晶。侧链的存在能够影响木聚糖链的构象[20],天然木聚糖链被预测为具有中等延伸性的半柔型(延伸构型),而改性木聚糖则表现为柔性线圈(卷曲构型)。
木葡聚糖是一种半纤维素多糖,存在于所有高等植物的初生细胞壁中。由于木葡聚糖不易结晶,因此难以在原子水平上确定其构象。Levy等[21]对木葡聚糖的静态和动态平衡构象进行了理论势能计算,结合NMR结果,证明了木葡聚糖的扭曲构象与该分子在溶液中的构象一致。Umemura等[22]利用AMBER软件对溶于显式水分子中的木葡聚糖低聚物进行分子动力学模拟,研究了水溶液中侧链折叠到主链上的机理,发现木葡聚糖侧链在主链上的折叠机制取决于主链的状态,揭示了木葡聚糖可以通过主链构象控制侧链折叠,这种能力可能在木葡聚糖水-醇溶液的胶凝过程中起作用。
2.2 半纤维素与纤维素的相互作用
2.2.1木聚糖与纤维素表面的分子吸附作用 在模拟体系中,吸附在纤维素表面的木聚糖经过充足时间平衡之后,接着在纤维素质心和木聚糖质心之间距离的表面法向分量上施加谐波电位,将木聚糖缓慢地从纤维素表面上拉离,直到木聚糖与纤维素表面完全分离,这个过程所需的自由能称为吸附自由能。自由能计算结果表明:侧链基团影响了木聚糖与纤维素表面之间的相互作用,通过α-(1→2)连接的取代基可增强木聚糖与纤维素之间的相互作用[19]。Mazeau等[23]利用Materials Studio软件模拟了气相中木聚糖在纤维素上的吸附过程,在分子动力学模拟过程中,木聚糖首先被纤维素表面吸引,吸附在该表面上,然后逐渐展开,最后以稳定的位置、方向和构象固定在纤维素表面上。木聚糖片段通过调节其构象与纤维素相互作用,侧链(如阿拉伯糖和葡萄糖醛酸)的存在会明显干扰木聚糖骨架的平衡取向。
2.2.2木葡聚糖与纤维素表面的分子吸附作用 木葡聚糖是双子叶植物和非淀粉单子叶植物的初生细胞壁中的主要半纤维素。显微镜技术显示木葡聚糖位于微原纤维之间并覆盖在微原纤维上[24-26]。
Zhao等[27]利用CHARMM软件模拟研究了表面疏水性对木葡聚糖吸附到纤维素微纤维上的影响,研究表明,(XXXG)3型木葡聚糖(X代表带有木糖侧链的葡萄糖单元,G代表葡萄糖单元)与纤维素微纤维(100)和(200)疏水表面的结合比与(110)、(010)和(1-10)亲水表面的结合更有利(图1)。这是因为在纤维素微纤维的疏水性表面上,(XXXG)3能够处于一个较为稳定的平坦构象中,其在纤维素(100)表面上的构型接近纤维素微纤维链本身的构型。
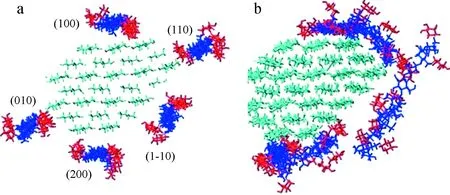
图1 木葡聚糖(XXXG)3在纤维素微纤维表面上吸附前(a)后(b)的结构模拟[27]
Zhang等[28]利用GROMACS软件研究了侧链变化对木葡聚糖在纤维素上的吸附的影响,研究发现:木葡聚糖与纤维素的相互作用受到木葡聚糖链长的影响,每个重复单元的平均相互作用能随链长的增加而降低。Hanus等[29]研究发现:木葡聚糖能够较为稳定地吸附在纤维素微纤维表面,主要由范德华力和静电吸附起作用,与静电能量相比,范德华能量通常较弱,当半乳糖和岩藻糖加入侧链时,木葡聚糖和纤维素之间的总相互作用能增加,半乳糖的添加起到更大的作用,但是较长的侧链增加空间位阻,会导致吸附受阻。此外,创建并优化了一个木葡聚糖-纤维素网络模型(图2),该模型与实验数据一致,但是由于技术原因(计算能力),与自然界中观察到的20~40 nm相比,10 nm的纤维间距离相当短,尽管如此,上述模型可能距离成功探索细胞壁成分结合原理又近了一步。
2.3 半纤维素在植物细胞壁中的作用
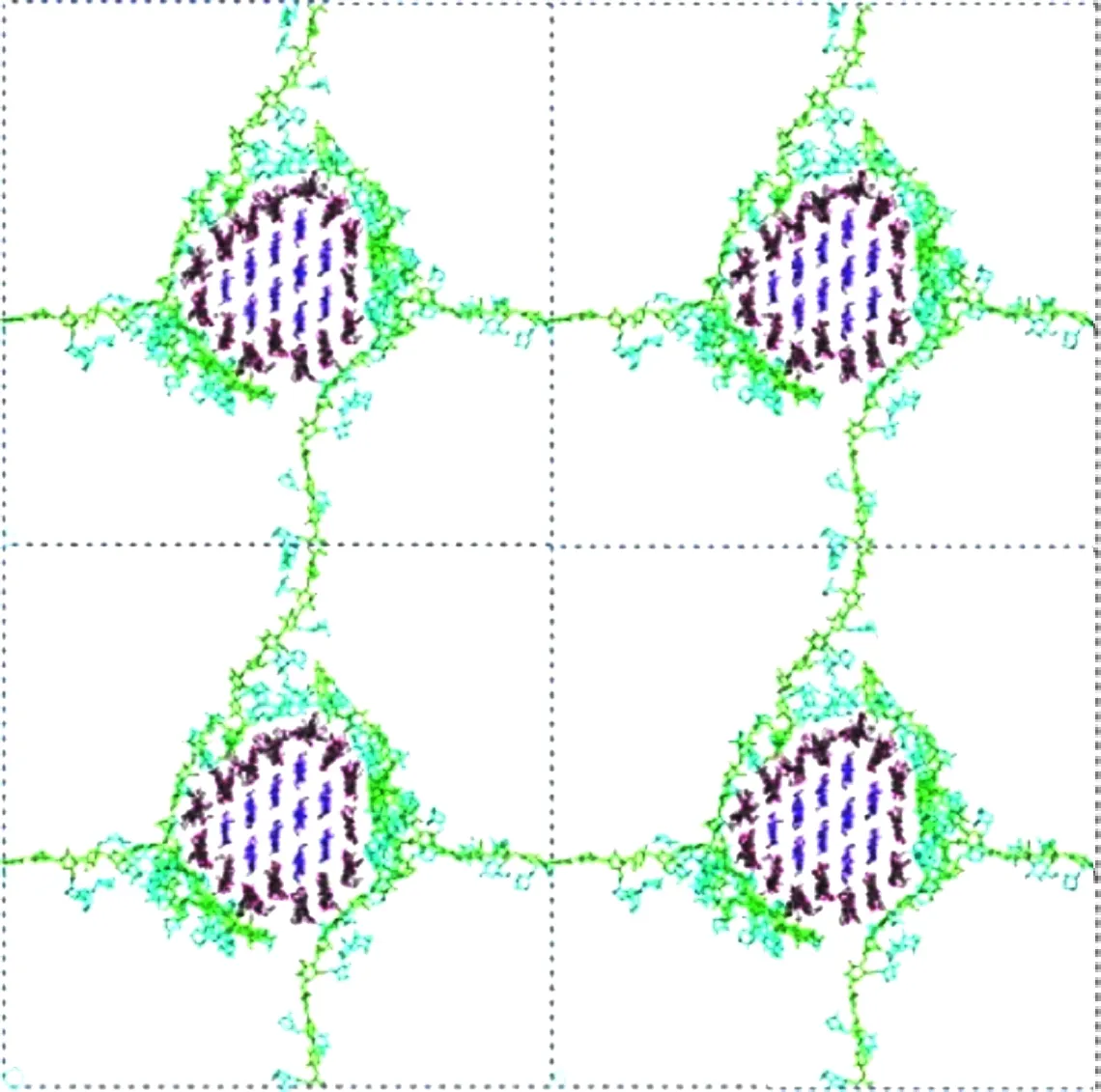
图2 木葡聚糖(外部)-纤维素(内部)网络模型[29]Fig.2 Network model of xyloglucan(outside)-cellulose(inside)[29]
植物细胞壁由纤维素、半纤维素和木质素组成,组成结构非常复杂。开发生物质资源需要破坏细胞壁原始结构,提取和分解组分,用以生产生物燃料和其他有价值的化学品。不同种类植物和同一植物的不同部位的细胞壁,其顽固性均不相同,细胞壁的顽固性与三大组分的组成结构有关。因此,充分了解细胞壁内的组分间的化学相互作用是有效控制木质纤维生物质降解性难易的基础。然而,由于细胞壁的高度复杂性,目前仍然缺乏这些结构的分子水平表征。
Silveira等[30]利用CHARMM软件研究了半纤维素成分对其与纤维素原纤维有效相互作用的影响,并揭示了半纤维素在原代细胞壁稳定性中的重要作用。研究发现:不同的半纤维素支链基团对细胞壁强度的相对贡献不同,如图3所示。半纤维素支链基团阿拉伯糖和葡萄糖醛酸主要作为氢键供体与纤维素亲水面的氧原子相互作用,而葡萄糖醛酸根阴离子充当氢键受体在纤维素表面与伯醇基团强烈相互作用;另外,乙酸根阴离子与葡萄糖醛酸根阴离子的分布相似,它们能够与羟基配位,对半纤维素与纤维素相互作用起主要贡献。这些结果表明:在植物中,半纤维素支链基团以互补的方式与纤维素表面相互作用,以稳定细胞壁的微观结构。
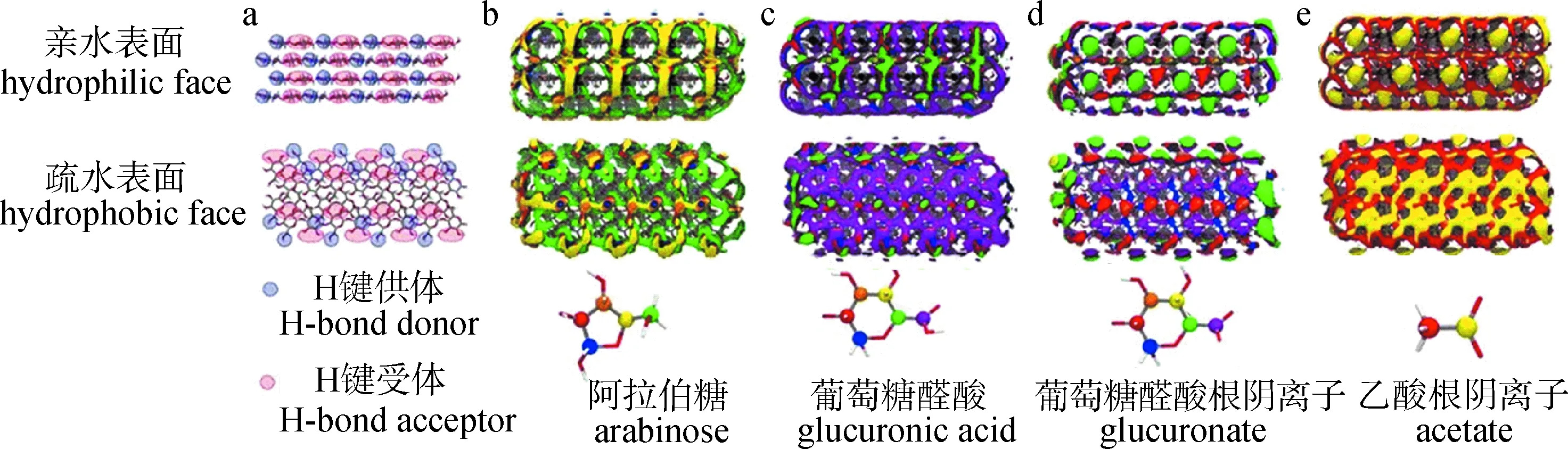
图3 纤维素纳米微晶的氢键供体和受体位点(a)及半纤维素支链基团的碳原子在纤维素纳米
植物细胞壁具有很强的抗应力性,同时对水合反应敏感,在水分含量较高的环境中可以显著膨胀。细胞壁中的纤维素对水分不敏感,而半纤维素是细胞壁中含量第二的组分,因此半纤维素与水的相互作用是一个关键因素。半纤维素在植物初生细胞壁的膨胀行为中起着重要的作用,分子动力学模拟为了解半纤维素与水的相互作用提供了手段。Kulasinski等[31]利用GROMACS软件研究了由结晶纤维素和无定形半纤维素组成的木材微原纤维的两相模型,对纤维素微纤维-半纤维素系统的模型进行水分吸附,模拟所得的吸附数据点与实验数据吻合较好。研究发现:水分子不会进入纤维素结晶区域,水分子优先占据结晶纤维素和半纤维素之间的界面,随着水分含量的增加,半纤维素吸附水分量明显增加,同时,半纤维素的平均密度降低,意味着半纤维素发生溶胀。溶胀可以理解为主要由氢键断裂引起的聚合物链有效体积的增加,可以认为结晶纤维素不会随水分含量而改变其机械性能,但半纤维素剪切模量降低了一个数量级以上,在这种情况下,随着水分含量的增加,木材的剪切刚度下降,导致结构变弱。
木材具有低密度、高刚度和高强度的优点,产生这种机械性能的关键是次生细胞壁,它在支撑木材抵抗重力方面发挥了重要作用。近二十年来,人们对木材细胞壁的详细组成和微观结构进行了大量的研究,木材特殊的力学性能吸引了研究者的兴趣,其潜在的变形机制能够为新材料的设计提供灵感。实际中观察到半纤维素链段是以不同的方式与纤维素微纤维结合,包括通过单桥结合、环结合和相邻纤维素微原纤维束之间的随机自由链端结合(图4)。Zhang等[32]利用AMBER软件研究了在施加一定剪切力下半纤维素链的形状对半纤维素-纤维素复合材料强度的影响,发现纤维素与半纤维素的接触面积、氢键以及半纤维素链主链上的共价键是决定复合体系界面强度的控制参数,其中随机结合模型能够承受的最大剪切力最小,桥结合模型能承受的最大剪切力较大,环结合模型相对于桥结合模型,纤维素与半纤维素之间的有效接触面积减少,导致模型能承受的最大剪切力降低。
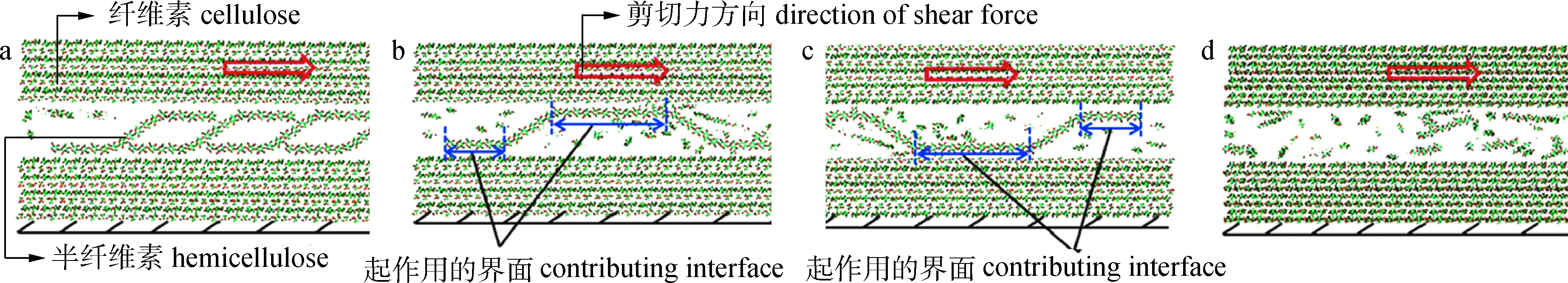
结合模型binding mode:a.桥bridge; b.环loop; c.翻转的环‘flipped’ loop; d.随机random图4 纤维素-半纤维素复合材料的初始原子构型的侧截面视图[32]
3 半纤维素转化过程模拟
3.1 半纤维素热解的分子模拟研究
在生物质热解过程中,通常认为纤维素、半纤维素和木质素3大组分是独立进行热解的。生物质热解反应过程是剧烈且极为复杂的热化学转化过程,包含分子键断裂和结合,分子结构异构化和裂化后生成的小分子聚合等过程,同时化学结构及特性上的差异使得三大素的热解规律不同,因此,针对这一过程的研究中出现了许多研究手段和方法,其中分子模拟方法能够从分子层面上剖析生物质三大组分热解过程、主要产物形成机理及中间产物的演化过程等[33]。
对生物质热解过程进行分子模拟时,一般是将三大素分开进行[3,34-35]。纤维素、半纤维素、木质素的热解模拟过程都分为3个阶段:低温、中温和高温阶段,不同物质每个阶段的温度范围不同。黄金保等[35]以聚合度为10的木聚糖为半纤维素模型化合物,运用HyperChem软件对其热解过程进行了分子动力学模拟,结果表明:温度上升到450 K时,开始有羟基键断裂,随温度继续升高,550 K时侧链苷键和主链苷键开始断裂,整个分子发生解聚,形成各种糖类单体,糖苷键断裂的同时发现环状单体内部键断裂形成各种分子碎片。模拟中,由于断裂的分子碎片没有排出,仍然留在模拟盒里,随温度的升高,这些分子基团继续分解,产生大量更小的分子碎片。因此,控制热解温度可以获得不同的热解产物。若为了获取生物油,快速热解试验的温度必须控制在中温阶段,且分解出来的产物必须快速分离和冷却;而为了获取气体产物,热解温度就必须控制在高温阶段。
3.2 半纤维素直接液化的分子模拟研究
生物质液化除了干燥木质纤维的快速热解之外,还包括溶剂中生物质的直接液化。直接液化技术是在适当的压力和温度下,将生物质加一定的催化剂和溶剂放在高压釜中,通入惰性气体或氢气,将生物质直接液化的技术,产物包括生物油或小分子化学品(糖类、呋喃类、酸类、醇类、醛类等)。
3.2.1半纤维素在水热条件下的形态 生物质转化过程中的预处理方法最初的目的是为了去除半纤维素和木质素,以提高纤维素酶水解效率。随着三大素综合利用的理念逐渐深入人心,研究者们开始调整预处理方式以使得三大素组分能够同时得到有效的利用。共溶剂增强木质纤维分馏是其中一种预处理方法,该过程是将生物质原料在有机溶剂-水共溶剂体系中进行水热转化,产物包括半纤维素来源的糠醛、纤维素来源的五羟甲基糠醛和乙酰丙酸,同时超过90%的木质素被提取出来,最终以固体产物形式产出。这种处理过程同时利用了三大组分,提高了生物质原料的整体产品产量[36]。由于生物质原料结构组分复杂,无法作为一个整体进行模拟,因此Smith J.C.团队利用GROMACS软件分别模拟了水热条件下纤维素、半纤维素和木质素在水-四氢呋喃共溶体系中的行为[37-39]。四氢呋喃-水共溶剂体系在不同的温度下表现出双相和单相的体系转变,在333~418 K之间是双相体系,温度高于418 K或低于333 K时,体系为单相。一般的水热温度在445~475 K之间,高于418 K,木聚糖处于四氢呋喃-水混溶体系中,相对于纯水条件,木聚糖溶解度降低。在单相四氢呋喃-水体系中,由于木聚糖在其中的溶解度降低而导致糠醛形成缓慢。
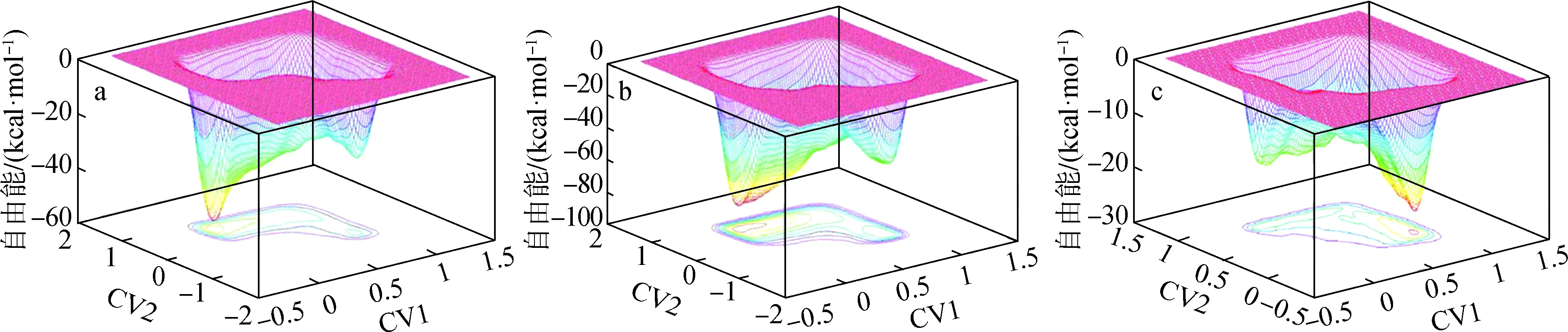
3.2.2半纤维素水解 植物三大素中,半纤维素的热稳定性最差,因此在直接液化过程中,半纤维素最早被溶出,通过水解反应进一步生成水溶性聚糖、寡糖和单糖。半纤维素的水解反应是化学反应,化学反应是复杂的动力学过程,涉及化学键的断裂和形成以及电子的转移。Dong等[40]利用CPMD软件研究气相和水溶液中酸催化木二糖水解反应的自由能势能面(图5)。自由能势能面上有两个最小值,一个对应于反应物状态,一个代表质子化已完成且醚键中的C—O键断裂的反应产物状态,这些位置之间的自由能差能够表示反应发生的难易程度,同时也能确定过渡产物的结构状态和自由能大小。水对溶液中的反应的壁垒起着至关重要的作用,质子部分去溶剂化及其向醚键合位点的迁移、醚键的质子化以及随后的C—O键的断裂是限速步骤,质子的部分去溶剂化对整个反应作用很大,说明酸浓度和质子运输将在木聚糖水解中起关键作用。
连接方式/反应相态connection mode/reaction phase state:a.β-1,4/气相β-1,4/gas phase;
3.2.3木糖降解反应 木聚糖型半纤维素链中最主要的单体为木糖。在对生物质原料进行预处理获取糖类的过程中,单糖很容易发生降解,造成单糖得率下降。糠醛是用途广泛的大宗生物质基化学品,而木糖脱水制备糠醛是半纤维素利用过程中重要的反应,但是在这一过程中,木糖会发生其他的降解反应而生成副产物。因此,理解木糖的降解机理十分重要。木糖在酸性介质下降解的第一步是糖环上的OH基团的质子化。Qian等[41]利用CPMD软件模拟了真空中木糖的降解行为,发现只有木糖C2—OH质子化后生成了2,5-酸酐中间体,最终才会生成糠醛。C1—OH和C4—OH质子化不会引起木糖的降解,质子对C3—OH的攻击导致木糖分子的裂解,这可能是其他降解产物如甲酸产生的原因。考虑到溶剂可能对反应造成影响,Qian等[42]对水溶液中木糖的降解行为也进行了研究。在稀酸环境下,糖环上羟基的质子化是糖降解的限速步骤。在模拟系统中,H2O、糖环上的氧和乙醇等共溶剂会与糖环上的羟基竞争质子。此外,溶剂水分子与糖环上OH基团之间的氢键相互作用能够破坏糖环上C—C、C—O键的相对稳定性,从而改变反应途径。水分子也可以很容易地从反应中间体中获取质子,从而终止环的闭合反应。在水溶液中,木糖C2—OH质子化之后,C3—C4键发生断裂,而在真空中,是C1—O5键发生断裂,这是由于羟基与水的氢键作用,使得C3—C4键比C1—O5键弱,说明了溶剂水对反应造成了影响。
高斯量子计算可用来获得反应物、过渡态和产物的能量,从而可以获得反应能与势垒。Nimlos等[43]利用Gaussian和CPMD软件进行研究,发现C2—OH质子化木糖分子降解形成糠醛是容易发生的,虽然C1—OH和O5质子化后有生成糠醛的可能性,但是这两个过程的反应势垒太高,无法与C2—OH质子化形成糠醛的过程竞争。木糖的5个氧原子中,O1、O2、O3、O4、O5的质子亲和力(PA)值分别为186.7、 191.3、 188.8、 187.2、 189.5 kcal/mol,O2的PA值最高,O1的PA值最低,说明O2最可能受到质子的攻击,而O1受到质子攻击的可能性较小(图6)。
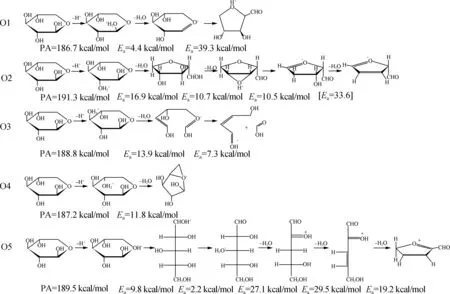
图6 木糖不同氧原子质子化的反应路径(Ea值是过程需要克服的最高活化能)[43]
Lin等[44-45]利用GROMACS软件研究了木糖降解为糠醛过程中的溶剂效应,对单相体系和两相体系中的木糖和糠醛进行了分子动力学模拟。单相体系中,发现不同溶剂在木糖与糠醛周围的分布方式与氢键结合不同,导致木糖向不同的反应路径转化,γ-戊内酯和γ-丁内酯可以提供糠醛保护壳,在糠醛生产中起积极作用。两相体系中,有机溶剂与木糖、水、糠醛之间的不同亲和力导致木糖和糠醛周围溶剂分布的不同。2-丁醇和四氢呋喃与木糖之间有更强的亲近力,二氯甲烷、甲基异丁基酮和二甲基四氢呋喃能够阻止更多水分子靠近糠醛,其中,二氯甲烷(DCM)/水体系中木糖和糠醛分别被严格限制在水相和有机相中(图7),有利于减少副反应的发生[46]。

图7 木糖(绿)/糠醛(蓝)在二氯甲烷-水体系Z轴上的运动轨迹[46]
3.3 生物质气化
糠醛是生物质(主要是半纤维素)解聚过程中的主要中间产物。目前,糠醛制氢已引起研究人员的关注。在适当的超临界水反应条件下,糠醛可转化为气体产物,例如H2、CO和CH4,但在一些条件下,糠醛被转化为焦油或焦炭。因此,糠醛在超临界水中的气化机理很重要。Jin等[47]利用ADF软件研究了超临界水中的糠醛转化机理,揭示了糠醛在超临界水中原子级的分解途径并获得产物分布。开环反应是热化学过程中的速率决定步骤,开环反应发生在杂环中的氧原子O6和连接醛基的碳原子C4之间形成的键。然后在C2—C3键处发生断裂,糠醛结构分解为两个片段。之后,这两个碎片在超临界水气化过程中分解为H2、CH4、CO、CO2等小分子。超临界水分子可以形成簇并向糠醛分子片段提供OH自由基,之后,超临界水簇中的H自由基会相互组合或和碎片中释放出来的H自由基组合产生H2。
蒽是生物质超临界水气化过程中重要的中间产物,也可以用于气化制氢,开发价格相对较低的金属是研究的重点。Jin等[48]利用ADF软件研究了Ni和Cu作用下蒽和糠醛催化气化的分子动力学,模拟结果表明:在超临界水气化过程中,Cu和Ni明显增加了H自由基的生成。Ni催化剂降低了H2的产量,显著提高了CO的产量,最终提高了合成气的产量。
4 半纤维素基材料
近年来,随着半纤维素分离纯化及改性技术的不断发展,其在造纸、食品、包装、能源、化工、环保和生物医药等领域表现出了潜在的应用价值。由于半纤维素具有无定形结构,与纤维素相比,其易在普通溶剂中溶解和化学改性,具有较好的加工性能。并且,半纤维素的主链和侧链上含有大量羟基、羰基、羧基和乙酰基等,可以利用这些基团对其进行酯化、醚化、氧化及交联等改性,改变其部分理化性质如溶解性、热稳定性和生物活性等,进而获得不同性质和不同功能的半纤维素基材料,如半纤维素基膜材料、水凝胶、医用材料、吸附材料和承重材料等[49]。目前关于分子模拟应用于半纤维素复合材料的研究较少,主要集中于木葡聚糖材料。
4.1 蒙脱土-木葡聚糖纳米复合材料
自然界中大部分承重材料是纳米结构的复合材料,其中蒙脱土-木葡聚糖纳米复合材料,在潮湿状态下具有较好的机械性能和氧气阻隔性能,这是因为湿润条件下木葡聚糖对蒙脱土有强烈物理吸附。为了提高人造纳米复合材料在这方面的性能,从实验和理论两方面对仿生复合材料进行研究具有重要意义。Wang等[50]为了更好地理解耐湿粘土纳米复合材料的材料设计原理,利用GROMACS软件对蒙脱土-木葡聚糖纳米复合材料模型进行分子动力学模拟,模拟结果显示:相对于酶促修饰的木葡聚糖,天然木葡聚糖对蒙脱土表现出更强的结合亲和力,酶促修饰的木葡聚糖相对于天然木葡聚糖,缺少了支链上的半乳糖基团,使得两种分子在蒙脱土表面构象不同,天然木葡聚糖相比于酶促修饰的木葡聚糖更加平坦,而酶促修饰的木葡聚糖在蒙脱土表面较为扭曲,因此吸附变得不利(图8(a))。这些结果有助于将来对具有耐湿性的粘土-多糖纳米复合材料的复杂多糖结构进行选择和分子剪裁。Wang等[51]进一步构建了两层蒙脱土夹层中间插入4条木葡聚糖链的模型(图8(b)),研究了木葡聚糖-蒙脱土纳米复合材料的尺寸稳定性溶胀与水分含量的关系。研究发现:木葡聚糖和蒙脱土相互作用密切,两者之间形成大量氢键,木葡聚糖同时与两层蒙脱土保持相互作用,即使在高湿度的环境下也能与蒙脱土板之间形成物理交联,因此在较高湿度下,木葡聚糖-蒙脱土纳米复合材料也能够保持较好的机械性能。
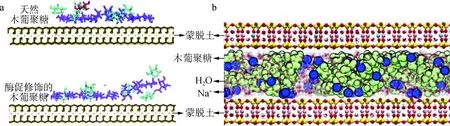
图8 木葡聚糖吸附在蒙脱土表面(a)及葡聚糖-蒙脱土模型(b)
4.2 木葡聚糖凝胶
木葡聚糖在水中溶解后,加入约20%(质量分数)醇溶剂,如甲醇、乙醇或丙醇,能够形成凝胶。小角度X射线散射研究表明:木葡聚糖凝胶化涉及到缩合和可溶性链域,醇可能在木葡聚糖周围聚集并在凝胶的缩合区域中充当黏合剂,但该机理尚未在分子水平上揭示。Umemura等[52]利用AMBER软件进行了木葡聚糖低聚物在水、水/甲醇和水/乙醇溶液中的分子动力学模拟,阐明了木葡聚糖的溶剂化和凝胶化机理(图9):木葡聚糖在水中分散,当加入醇时,醇分子会附着在木葡聚糖上,并通过减缓木葡聚糖的膨胀收缩运动,使其在某些区域聚集。在链的聚集过程中,黏合剂醇可以介导木葡聚糖之间的相互作用。木葡聚糖网络中含有丰富的水,而加入酒精会破坏和改变其氢键网络。这种水分子被醇分子通过氢键作用包围,并与醇保留在木葡聚糖的网状结构中。这种状态的木葡聚糖,即含有丰富水分的网状物,呈现出凝胶状态。
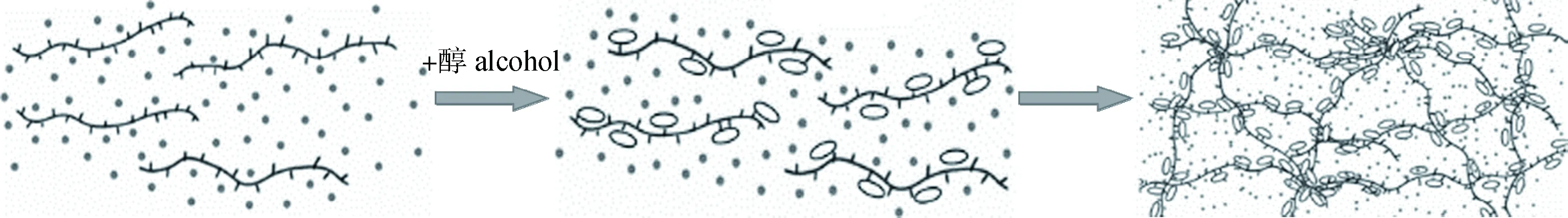
图9 木葡聚糖凝胶化机理示意图[52]
5 结语及展望
木质纤维生物质具有复杂的成分及结构,研究者们对其物理化学性质、转化制备化学品、改性制备材料等方面进行了大量的实验研究。分子模拟是实验方法的重要辅助手段,能够提供原子和分子水平上的见解,对物质的物理化学性质和反应过程具有更加直观的指导意义。虽然分子模拟主要是对实验起到补充作用,一般认为模拟结果是间接的而实验结果是直接的,但是在一些实验无法进行的情况下,分子模拟可能是更好的或唯一的选择。目前关于半纤维素的分子模拟研究还较为缺乏,一方面是由于分子模拟应用于生物质研究还不久,另一方面是由于半纤维素本身结构的复杂性。目前半纤维素分子模拟应用存在许多空白领域,包括半纤维素在细胞壁中所起的作用、半纤维素液化生产生物油、木糖异构化生产木酮糖、半纤维素与木质素之间的结合方式、其他的半纤维素基材料等,需要进一步的探索与研究。将不同尺度的分子模拟方法(量子力学方法、全原子力场的方法和粗粒度方法)应用于关于半纤维素的研究中是必要的,这种多尺度的方法不仅能够研究反应途径、过程能量、电子结构,还能够研究物质间相互作用、溶剂效应、大分子结构等,无论是化学反应过程还是物理作用过程都能够提供更深层次的解释。
猜你喜欢
阅读(中年级)(2022年9期)2022-10-08
精细石油化工(2022年5期)2022-09-21
林产化学与工业(2022年4期)2022-09-13
红蜻蜓·低年级(2022年5期)2022-05-11
红蜻蜓·低年级(2022年2期)2022-03-19
红蜻蜓·低年级(2021年12期)2021-12-19
粮食加工(2021年5期)2021-11-01
石油沥青(2021年6期)2021-02-10
湖南饲料(2019年5期)2019-10-15
太原理工大学学报(2019年4期)2019-08-05
