
4种化学物质对HaCaT细胞氧化应激相关基因表达的影响
2022-06-10 13:55段惠娟孙照刚郝卫东魏雪涛褚洪迁
癌变·畸变·突变 2022年3期
段惠娟,孙照刚,郝卫东,魏雪涛,褚洪迁,*
(1.北京市结核病胸部肿瘤研究所耐药结核病研究北京市重点实验室,北京 101149;2.首都医科大学附属北京胸科医院转化医学研究室,北京101149;3.北京大学公共卫生学院毒理学系,北京 100191;4.食品安全毒理学研究与评价北京市重点实验室,北京 100191)
氧化应激是指体内活性氧(reactive oxygen species,ROS)大量产生,超岀了机体清除能力,破坏了机体正常的氧化/还原平衡状态,造成蛋白质、脂质、核酸等生物大分子物质氧化损伤和功能紊乱,影响机体正常代谢过程而形成的一种异常应激状态。它是引起机体衰老、炎症和多种慢性疾病如糖尿病、高血压等的主要原因之一[1-2]。当细胞受到外源或内源刺激时,产生的ROS 会激活多条信号通路。Kelch 样ECH 相关蛋白1(Kelch like ECH associated protein 1,Keap1)-核转录因子E2 相关因子2(nuclear factor erythroid 2 related factor 2,Nrf2)-抗氧化反应元件(antioxidant response element,ARE)信号通路是一个综合的氧化还原反应体系,有多个激活途径维持细胞的氧化还原平衡及代谢,能调节机体1%~10%的基因[3-4]。Keap1-Nrf2-ARE 信号通路与氧化应激相关的多种疾病,包括癌症、阿尔茨海默病、帕金森病、糖尿病等均有相关性[5]。抗氧化剂可影响氧化应激相关基因的表达,进而作用于Keap1-Nrf2-ARE 信号通路发挥抗氧化作用。
姜黄素(curcumin)、槲皮素(quercetin)、叔丁基对苯二酚(tert-Butylhydroquinone,tBHQ)是食品药品中经常添加的抗氧化剂,研究表明上述3 种化学物质是Keap1-Nrf2-ARE 信号通路的小分子激活剂,可与Keap1 半胱氨酸上的巯基通过氧化或烷基化形成共价加合物,诱导Keap1 和Nrf2 解离,抑制Nrf2 通过泛素化降解,进而促进Nrf2 核转位[6]。但是,上述物质引起的具体哪种氧化应激相关基因发生改变尚未见报道。本研究利用实时荧光定量PCR(quantitative realtime PCR,qPCR)的方法,以过氧化氢(hydrogen peroxide)为对照、筛选槲皮素、姜黄素及叔丁基对苯二酚对84种氧化应激相关基因表达的影响,拟为抗氧化剂的添加提供依据。
1 材料与方法
1.1 主要材料与仪器
过氧化氢、槲皮素、姜黄素、叔丁基对苯二酚均购自美国BD公司,DMEM高糖培养液、胰酶、DMSO均购自Sigma Aldrich 公司,胎牛血清购自BD Pharmingen 公司。RNA 提取及反转录试剂盒购自QIAGEN 公司。人类氧化应激PCR 芯片(RT2ProfilerTMPCR Array Human Oxidative Stress Plus) 测定由QIAGEN 公司完成。姜黄素先溶于DMSO 配成储备液,再用新鲜的DMEM完全培养基稀释成不同的浓度使用,其余3 种化学物质用新鲜的DMEM 完全培养基稀释得到。
i-MARK 型酶标仪(美国Bio-Rad 公司);RNA 质量及浓度测定仪(美国Thermo公司);CO2培养箱和荧光定量PCR(美国Thermo 公司);细胞计数仪NucleoCounter(丹麦ChemoMetec公司)。
1.2 细胞系及芯片
HaCaT 细胞购自美国典型培养物保藏中心(American Type Culture Collection,ATCC)。所需培养液为DMEM 高糖完全培养液(含0.37%碳酸氢钠,10%灭活的胎牛血清,2 mmol/L左旋谷氨酰胺,100 U/mL青霉素及100 μg/mL链霉素)。细胞贴壁培养在75 cm2培养瓶,于37 ℃、CO2体积分数为5%的培养箱中培养。
人类氧化应激PCR芯片用来探查84个氧化应激相关基因的表达(表1)。过氧化物酶在此阵列上,包含谷胱甘肽过氧化物酶(GPx)和过氧化还原酶(TPx)。阵列上还包括ROS代谢相关基因,如氧化应激应答基因及参与过氧化物代谢的基因(如SOD等)。还包括其他由16个试验确定的生物标志基因。每个阵列上均设有对照,用ΔΔCT方法计算相对表达量。使用qPCR 技术检测氧化应激基因的表达,基因的表达超过3 倍认为有意义。
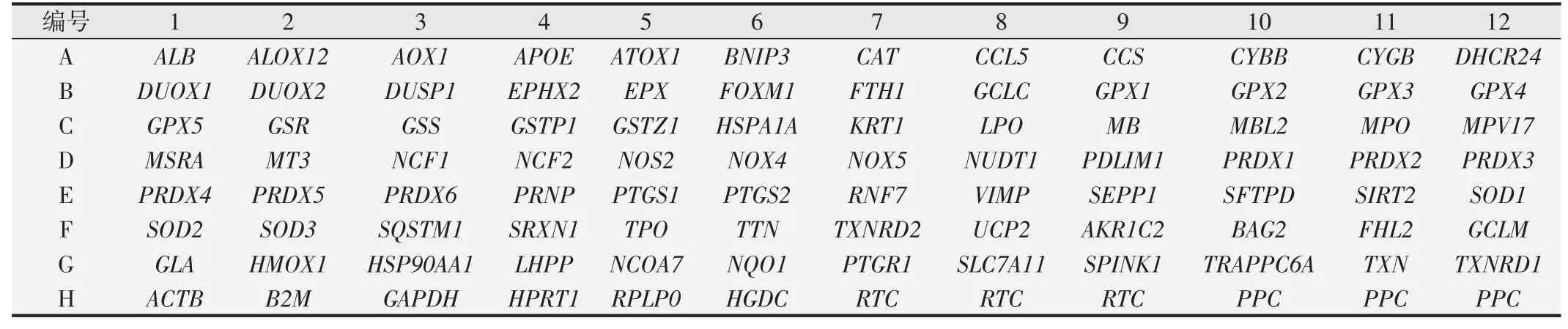
表1 人类氧化应激相关基因
1.3 方 法
1.3.1 四甲基噻唑蓝(MTT)法检测4 种化学物对HaCaT细胞存活率的影响取对数生长期的HaCaT细胞,使用胰酶分离后进行细胞计数,使用完全DMEM培养基调整细胞浓度为2×105/mL,于96 孔板中加入100 μL HaCaT细胞悬液和100 μL不同浓度的过氧化氢及受试物。过氧化氢的终浓度为0、0.125、0.25、0.5、1、2.5、5、7.5、10 mmol/L,姜黄素的终浓度为0、0.625、1.25、2.5、5、10、20、40、80 μmol/L,槲皮素的终浓度为0、300、400、500、600、700、800、900、1 000 μmol/L,叔丁基对苯二酚的终浓度为0、1.25、2.5、5、10、25、50、75、100 μmol/L。每个剂量设6 个平行孔,将培养板置于CO2体积分数为5%、37 ℃培养箱20 h 后,每孔加入20 μL MTT(5 mg/mL,用PBS 配制成pH=7.4),继续培养4 h,1 000 r/min 离心5 min 后取上清,加入150 μL DMSO,振荡混匀10 min后,于570 nm波长处测吸光度值,按下列公式计算细胞存活率。

1.3.2 RNA 提取及质量分析取对数生长期细胞,制成终浓度为2×105/mL 的细胞悬液,将细胞接种在6 孔板内(4×105个/孔),在37oC 培养箱内培养过夜。分别用过氧化氢(0.5 和1.25 mmol/L),姜黄素(5 和20 μmol/L),槲皮素(200 和500 μmol/L)及叔丁基对苯二酚(10和50 μmol/L)刺激HaCaT 细胞4 h,24 h后重复1 次,将细胞用PBS 洗两次,用胰酶消化,转移到EP 管内,300 g 离心5 min,弃掉上清。加入1 mL TRIzol,反复吹打几次,将液体转移到新的EP 管中。每管加入0.2 mL氯仿:异戊醇混合液,剧烈混合。室温孵育3 min后,4 ℃、12 000 g离心5 min,将上层水相转移到新的EP 管内,每管加入0.53 mL 无水乙醇。然后用RNeasy Mini Kit(QIAGEN)纯化RNA,将RNA 置于-80 ℃保存。合成cDNA 之前使用Agilent RNA 600 Nano Kit检测RNA的质量及浓度。
1.3.3 cDNA合成及qPCR根据RT2First Strand 试剂盒(QIAGEN)步骤要求,每管使用0.5 μg 的RNA 作为模板合成cDNA。合成cDNA 之后,进入Real-time PCR 步骤(每个样本重复3 次),在5 mL 管内配制反应液,2×RT2SYBR Green Mastermix 1 350 μL,cDNA合成反应液102 μL,RNase-free water 1 248 μL,总体积为2 700 μL。小心将RT2Profiler PCR Array(96孔板)从密封袋中取出,每孔加入25 μL上述PCR混合液,室温下1 000g离心1 min 去除气泡,然后在Applied Biosystems 7500 Fast Real Time PCR System中进行反应。反应条件为:95 ℃、10 min,然后95 ℃、15 s,40个循环,最后60 ℃、1 min,得到每孔的CT值。根据2-ΔΔCT计算基因相对表达量。
1.4 统计学分析
应用GraphPad Prism 8.0.1、SPSS 22.0 和Microsoft Office Excel 2010 对数据进行统计及作图,用配对样本t检验对细胞活率进行统计分析。
2 结 果
2.1 过氧化氢、槲皮素、姜黄素、叔丁基对苯二酚对HaCaT细胞的毒性
见图1。结果显示,过氧化氢浓度>1 mmol/L时,HaCaT 细胞的存活率在80%以下(图1A)。姜黄素浓度>10 μmol/L 时,HaCaT 细胞的存活率在80%以下(图1B)。而槲皮素在浓度为1 000 μmol/L 仍未显示出明显的细胞毒性(图1C)。叔丁基对苯二酚在浓度>50 μmol/L时,HaCaT细胞的存活率在80%以下(图1D)。

图1 不同浓度的4种化学物对HaCaT细胞存活率的影响
2.2 RNA浓度测定及质量分析
如图2 所示,以槲皮素为例,可以看出,RNA 条带及吸收峰完整(图2A和C),RNA质量得分均为10分(图2B),可用来进行后续试验。
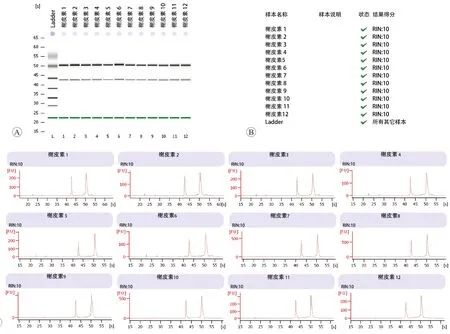
图2 RNA质量分析
2.3 4 种化学物质对HaCaT 细胞氧化应激相关基因表达的影响
给予不同浓度的过氧化氢、姜黄素、槲皮素及叔丁基对苯二酚处理HaCaT细胞4 h及24 h后,均能引起HaCaT细胞氧化应激相关基因的表达发生改变。如图3 和表2 所示,过氧化氢0.5 mmol/L 处理HaCaT 细胞4 h 后不会引起HaCaT 氧化应激相关基因的表达发生改变,1.25 mmol/L 处理HaCaT 细胞4 h 引起ALB、CCL5、SOD3、HMOX1、SPINK1共5 种基因表达上调。24 h 时,0.5和1.25 mmol/L处理HaCaT细胞均能引起基因DUOX2和AKR1C2基因表达升高;此外,1.25 mmol/L 处理HaCaT 细胞24 h 后还可引起HMOX1和TXNRD1基因表达上调,CCL5基因表达下调。

图3 H2O2对HaCaT细胞氧化应激相关基因表达的影响
如图4 和表2 所示,姜黄素5 和20 μmol/L 处理HaCaT 细胞4 h后均引起HSPA1A、NOS2和HMOX1基因表达的上调,不同的是,5 μmol/L处理细胞后表达上调的基因还包括CCL5、CYGB、EPHX2、EPX、GCLC、GPX2、GPX3、KRT1、SRXN1、SLC7A11及TXNRD1共11种基因,未引起任何氧化应激相关基因表达降低;20 μmol/L 处理后表达上调的基因还包括ALB、AOX1、APOE、DUSP1、LPO、MB、MPO、TTN及SPINK1,并且引起GSR基因表达下调。5 μmol/L处理HaCaT 细胞24 h 后引起HMOX1、NQO1及SLC7A11共3 种基因表达上调,DHCR24、KRT1及SEPP1共3种基因表达下调。由于使用20 μmol/L 姜黄素处理24 h 后细胞存活率较低,提取的RNA 浓度少,所以导致24 h的数据缺失。

图4 姜黄素对HaCaT细胞氧化应激相关基因表达的影响
如图5 和表2 所示,槲皮素200 和500 μmol/L 处理HaCaT 细胞4 h 均能引起ALB、NOS2、HMOX1共3种基因表达上调,此外,500 μmol/L 处理4 h 后还能引 起DUSP1、GPX3、NCF1和PTGS2基 因 上 调,SEPP1基因表达下调。200 和500 μmol/L 处理HaCaT细胞24 h后均引起BNIP3和PTGS1共2种基因表达上调,MPO和SEPP1共2 种基因表达下调。此外,200 μmol/L 处理HaCaT 细胞24 h 后还可引起EPX基因表达下调,500 μmol/L处理HaCaT细胞24 h后还可引起CYGB、PTGS2和SPINK1共3 种基因表达上调,KRT1和MB共2种基因表达下调。

图5 槲皮素对HaCaT细胞氧化应激相关基因表达的影响
如图6 和表2 所示,叔丁基对苯二酚10 和50 μmol/L 处 理HaCaT 细 胞4 h 均 引 起EPX、GCLC、SRXN1、HMOXN1、SLC7A11及TXNRD1共6种基因表达上调,50 μmol/L 处理后还引起EPHX2、GSR、AKR1C2、GCLM、NQO1基因表达上调,均未引起任何氧化应激相关基因表达降低。叔丁基对苯二酚10和50 μmol/L处理HaCaT细胞24 h后,均引起AKRIC2和SLC7A11基因表达上调。此外,10 μmol/L处理后表达上调的基因还包括ALB、EPX、MB,50 μmol/L 处理后表达上调的基因还包括CYBB、GCLC、SRXN1、GCLM、HMOX1、NQO1。10 μmol/L处理后引起AOX1基因表达下调,50 μmol/L 处理后引起CYGB、EPX、KRT1、MPO及NCF1共5种基因的表达下调。

表2 4种化合物不同浓度下对基因表达的影响
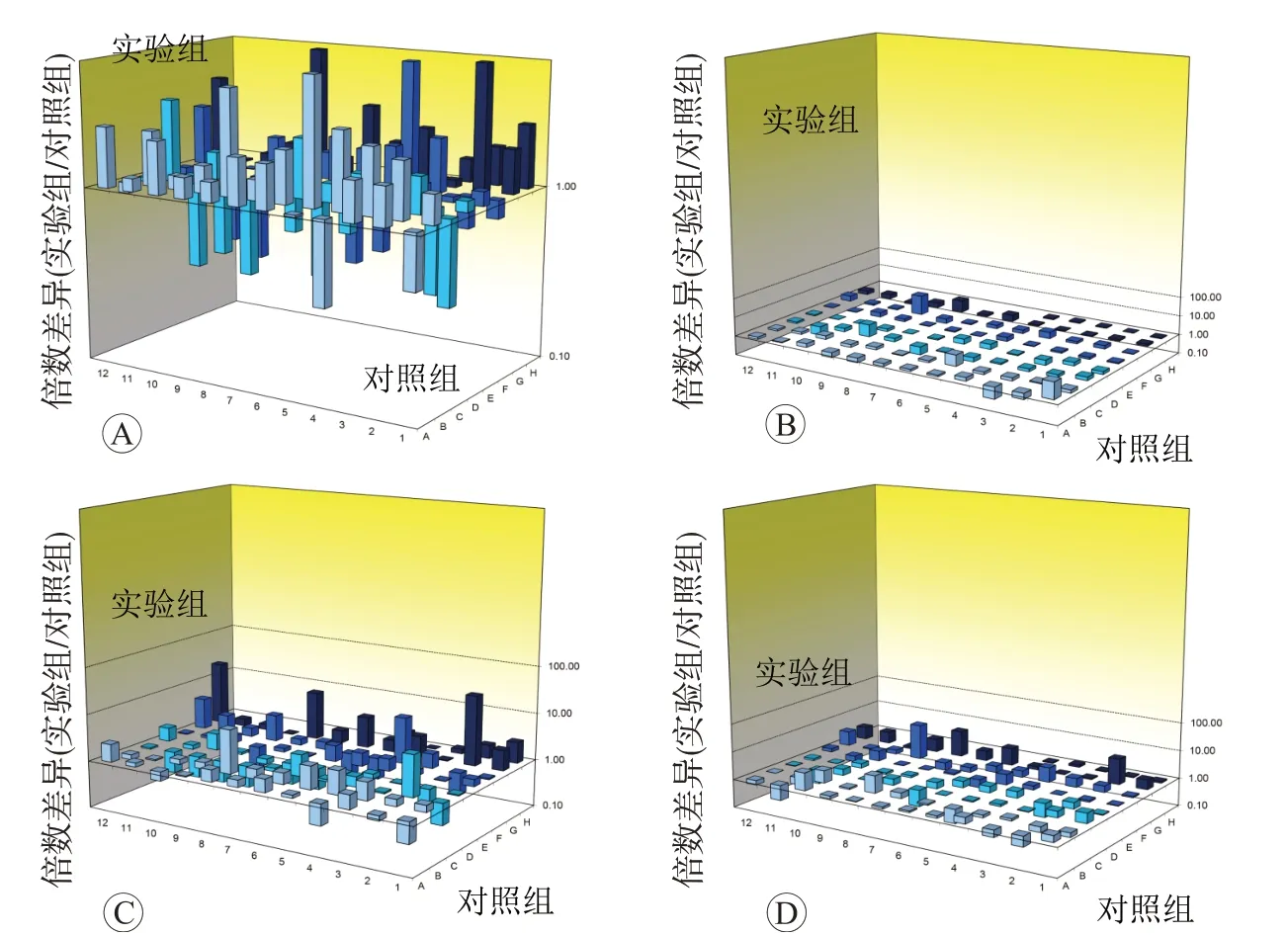
图6 叔丁基对苯二酚对HaCaT细胞氧化应激相关基因表达的影响
3 讨 论
人体表皮经常暴露于大量的物理或化学因子,能够引起细胞的氧化应激反应[1]。ROS 是氧化应激最重要的调节因子,并且能够引起细胞的氧化损伤[3]。ROS是小的生物分子,是需氧有机体氧代谢的副产物,在体内能够持续产生。尽管细胞产生ROS能够产生毒性作用,如细胞的坏死和凋亡,但是ROS也在细胞信号转导过程中起着重要作用[5]。在正常生理状态下,由于体内抗氧化机制的存在,ROS 的产生处于平衡状态。在病理状态下,由于ROS的产生增多或是抗氧化系统的受损导致ROS 的产生达到较高水平[2,7-9]。ROS能够影响细胞的氧化还原平衡状态,最终导致生物大分子如蛋白质、脂质和核酸的改变。研究表明转录因子Nrf2 能够调节氧化应激相关基因的表达[4,10],但涉及哪些基因及可诱导基因表达的程度还不清楚[11]。有研究表明植物来源及人工合成的一些化合物能够有效激活Nrf2。姜黄素、槲皮素、叔丁基对苯二酚3 种化学物质作为Keap1-Nrf2-ARE 信号通路的小分子激活剂,被认为是低活性的抗氧化剂,然而,这些化合物在较高浓度时会对细胞产生不利影响,而且其引起的具体氧化应激相关基因的改变目前尚无报道。因此,本研究基于HaCaT细胞,研究过氧化氢、槲皮素、姜黄素及叔丁基对苯二酚对细胞氧化应激相关基因表达的影响,关于Nrf2途径对减少氧化应激的影响的研究将有助于确定每种化合物的安全剂量,拟为抗氧化剂的添加提供依据。研究结果表明,不同浓度以及不同处理时间,上述4 种化合物可引起HaCaT 细胞氧化应激相关基因表达上调和下调。
过氧化氢作为信号转导分子在免疫细胞活化、血管重塑、细胞凋亡等生物学过程中起着重要的作用,同时也是体内几种经典的酶(葡萄糖氧化酶、乙醇氧化酶、胆固醇氧化酶等)催化的生物化学反应的产物[12]。本研究结果显示,过氧化氢浓度为≤1 mmol/L 时,HaCaT 细胞的存活率在80%以上。细胞在正常状态下也能产生低浓度的过氧化氢,在过氧化氢酶的作用下分解成水和氧。过氧化氢能够激活Nrf2,进而发挥抗氧化作用[13]。过氧化氢本身作为ROS,能够直接诱导氧化应激的发生。本研究结果显示,过氧化氢0.5 mmol/L 处理HaCaT 细胞4 h 后不会引起HaCaT 氧化应激相关基因的表达发生改变,1.25 mmol/L处理HaCaT细胞4 h 引起ALB、CCL5、SOD3、HMOX1、SPINK1共5种基因表达上调。24 h 时,0.5和1.25 mmol/L处理HaCaT 细胞均能引起基因DUOX2和AKR1C2表达升高;此外,1.25 mmol/L过氧化氢处理HaCaT细胞还可引起HMOX1和TXNRD1基因表达上调,ALOX12、AOX1及CCL5基因表达下调。
姜黄素和槲皮素均属于植物多酚类,是天然的抗氧化剂,可增强机体抵抗ROS 或活性氮自由基(reactive nitrogen species,RNS)的能力,均与Keap1-Nrf2-ARE信号通路相关[14]。在临床上,姜黄素被用来降低术后炎症反应。除了氧自由基清除作用,姜黄素还能降低促炎因子的表达(如TNF-α、IL-β、IL-6)[15]。同时,姜黄根来源的姜黄素能够激活Nrf2,参与调节氧化应激反应[16]。本研究结果显示,与过氧化氢相比,不同浓度姜黄素处理HaCaT 细胞4 h 后不仅可引起ALB、CCL5、HMOX1和SPINK1共4 种 基 因 上 调外,还 可 引 起HSPA1A、NOS2、CYGB、EPHX2、APOE、LPO等多种基因的上调,并且引起GSR基因表达下调。与本研究一致的是,张春云等[17]使用姜黄素对四氯化碳(CCl4)诱导鲫鱼肝损伤的作用时,发现姜黄素处理后的鲫鱼肝切片中,ALB的含量显著增高。但也有研究表明[18],姜黄素受浓度影响,具有亲氧化剂和抗氧化剂双向调控性,但促氧化剂的作用剂量目前还未见报道,需进一步研究。有研究表明槲皮素具有抗氧化,清除自由基,抑制脂质过氧化的功能,具有抗肿瘤等作用。槲皮素也是通过Nrf2 来发挥抗氧化作用[19]。本研究中,槲皮素200 和500 μmol/L 处理HaCaT 细胞4 h 均能引起ALB基因表达上调,但与过氧化氢相比,500 μmol/L 处理4 h 后还能SEPP1基因表达下调,24 h后可引起KRT1基因表达下调。
叔丁基对苯二酚与上述3 种化合物一样,能够通过激活Nrf2,控制氧化应激相关基因的表达,降低ROS,促进抗氧化酶的活力,进而调节氧化应激反应,活化Nrf2 信号通路[20-22]。叔丁基对苯二酚10 μmol/L 处理HaCaT 细胞24 h 能引起ALB、HMOX1等基因表达的上调,与姜黄素和槲皮素有相同的效应。刘辉等[23]对增强心肌细胞抗氧化应激能力机制的研究结果表明,叔丁基对苯二酚处理后的细胞,Nrf2的蛋白表达明显增强,NQO1和GCLC的表达也呈一致性升高,这与本研究结果一致。
Keap-Nrf2-ARE 信号通路对抗氧化防御机制非常重要,激活Keap-Nrf2-ARE系统,对开发抗氧化药物具有重要意义。本研究显示,HMOX1基因在四种化合物处理后均上调表达,其中姜黄素和叔丁基对苯二酚引起的基因表达量上调明显。HMOX1作为一种细胞保护酶,具有抗炎和抗氧化特性,可在多种有害环境刺激和疾病状态下诱导产生,已有研究证明Nrf2 是HMOX1启动子激活的重要转录因子,HMOX1启动子被促氧化刺激激活,主要是通过Nrf2转录因子。氧化应激介导的HMOX1启动子激活诱导lacZ基因表达[24]。HMOX1基因转录可能会受到热疗、炎性细胞因子或特定的磷酸化依赖性信号级联的刺激[25]。此外,研究表明,BNIP3 蛋白通常在正常细胞和组织中低表达,包括骨骼肌、心肌细胞、大脑和心脏的细胞和组织,并通过参与多种细胞信号通路参与多种细胞功能[26],但本研究结果显示,BNIP3基因在榭皮素的作用下上调表达明显。
综上所述,姜黄素、槲皮素及叔丁基对苯二酚作为Nrf2的激活剂,研究其对84种氧化应激相关基因表达的影响,为抗氧化剂的添加提供了实验支持。本研究结果表明过氧化氢、姜黄素、槲皮素及叔丁基对苯二酚分别处理HaCaT细胞后,引起部分氧化应激相关基因表达水平升高,部分表达水平降低。而且处理浓度越高,处理时间越长,对HaCaT细胞氧化应激相关基因表达水平的影响越大,为抗氧化剂的添加提供一定的理论依据。
猜你喜欢
新农业(2022年16期)2022-11-07
食品科学(2022年18期)2022-10-09
湖北农业科学(2022年16期)2022-09-29
中国药房(2022年8期)2022-04-27
家庭百事通·健康一点通(2021年1期)2021-02-24
现代养生·上半月(2021年1期)2021-01-28
分析化学(2020年7期)2020-07-14
文萃报·周五版(2018年25期)2018-08-13
文萃报·周二版(2018年26期)2018-08-04
新教育时代·教师版(2017年29期)2017-09-08
