
谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗生理特性的影响
2022-06-14 05:34苗如意杨玉花梁燕萍
中国土壤与肥料 2022年4期
雷 阳,乔 宁,苗如意,杨玉花,梁燕萍
(1.山西农业大学园艺学院,山西 太原 030031;2.山西农业大学农业基因与资源研究中心,农业农村部黄土高原作物基因资源与种质创制重点实验室,山西 太原 030031)
镉(Cd)是植物体内的有害元素,极易被植物吸收,并通过维管束从根转移至地上部[1]。镉元素在植物体内大量富集会引起活性氧(ROS)积累,造成氧化胁迫,影响RNA和蛋白质的正常表达合成,导致植物生长发育迟滞、光合作用受阻[2]。据统计,我国土壤重金属污染情况极其严重,污染超标率高达19.4%,每年因重金属污染造成的粮食损失接近1000万t,其中36.08%的重金属污染为镉含量超标[3-4]。
谷胱甘肽(GSH)是谷氨酸、半胱氨酸和甘氨酸缩合而成的水溶性氨基酸衍生物[5],GSH可直接或间接清除ROS[6],同时也能通过后续反应与镉等重金属离子发生螯合,降低重金属对植物的胁迫[7]。γ-谷氨酰半胱氨酸合成酶(γ-GCS)是植物体内GSH合成路径上游的关键酶,丁硫堇(BSO)是该酶的抑制剂,能够有效减少植物体内GSH的含量,从而降低植物对重金属的耐受性[8]。蔡仕珍等[9]研究表明,外源GSH可有效提高镉胁迫下绵毛水苏的生物量、光能的捕获与转换能力、抗氧化酶活性,有效缓解镉胁迫对植株幼苗生长的抑制作用。杨志峰等[10]研究表明,GSH可有效减轻低温胁迫下叶绿体膜脂的过氧化程度,增强黄瓜幼苗的抗寒性。Zhou等[11]研究表明,GSH可有效缓解盐胁迫对番茄光合作用的抑制,并提高抗氧化物酶活性,而添加BSO则会起到与GSH相反的作用。
近年来,随着我国镉污染的情况不断恶化,科研工作者做了大量关于缓解植物镉胁迫的研究[12-14]。然而,关于GSH和BSO对镉胁迫下辣椒叶片和根部生理影响的研究尚未见报道。鉴于此,本研究针对辣椒生产中出现的镉污染问题,探究了镉胁迫下GSH和BSO对辣椒幼苗各项生理性指标的影响,旨在探讨GSH对辣椒逆境响应的调节机理,丰富辣椒抗逆机理的理论认识,以期为辣椒耐重金属栽培提供理论依据。
1 材料与方法
1.1 试验材料
供试辣椒品种为晋椒503(Capsicum annuumL.cv. Jinjiao 503),由山西省园艺学院辣椒育种团队提供。CdCl2、GSH和BSO均购自Sigma公司。
1.2 试验方法
1.2.1 种子萌发试验
选取均匀、无病害的晋椒503辣椒种子,用灭菌水在室温下浸泡4~5 h。进而用75%乙醇冲洗20 s,15%的NaClO冲洗15 min,灭菌水洗涤3次,放入真空干燥箱40℃烘干30 min,播种于72孔的穴盘中,置于25℃(光/暗,12 h/12 h)光照培养箱中培养。培养15 d后,取长势一致的幼苗,分成对照组(CK,处理液为灭菌水)和4个处理组(表1)移栽至营养箱中。在处理12 d后分别取样测定各指标含量,每处理测定3株,3次重复。

表1 试验处理
1.2.2 幼苗生理指标的测定
叶部和根部Cd含量的测定:称取晋椒503辣椒叶片或根部0.1 g,加入硝酸∶过氧化氢(H2O2)=3∶2的混合溶液5 mL,在消解炉中160℃消解4 h,蒸馏水定容至50 mL,火焰原子吸收分光光度计测定。
叶绿素a、叶绿素b和类胡萝卜素含量的测定:避光条件下取晋椒503辣椒幼苗叶片2 g,采用丙酮溶解法提取,然后分别在470、649和665 nm波长下测定。
GSH、氧化型谷胱甘肽(GSSG)含量、超氧阴离子(O2·-)产生速率和H2O2含量的测定采用Murshed等[15]的方法。根据试剂盒说明书(苏州格锐思生物技术有限公司)测定抗氧化物酶和抗坏血酸(AsA)-GSH循环关键酶活性。过氧化物酶(POD):在470 nm比色,记录10 s和1 min 10 s吸光值。超氧化物歧化酶(SOD):静置30 min,在450 nm比色。过氧化氢酶(CAT):静置30 min,在510 nm比色。抗坏血酸过氧化物酶(APX):在290 nm比色,记录30 s和5 min 30 s吸光值。单脱氢抗坏血酸还原酶(MDHAR):在340 nm比色,记录10 s和5 min 10 s吸光值。脱氢抗坏血酸还原酶(DHAR):在265 nm比色,记录10 s和3 min 10 s吸光值。谷胱甘肽还原酶(GR):在412 nm比色,记录30 s和10 min吸光值。谷胱甘肽过氧化物酶(GPX):在340 nm比色,记录3 s和10 min吸光值。γ-GCS:在412 nm比色,记录1 min吸光值。
1.3 数据处理
采用Excel 2016整理数据和作图;SAS 1.2分析数据,Duncan法进行多重比较。
2 结果与分析
2.1 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗生物量的影响
由表2可知,与CK相比,Cd处理的根长、株高、地上部鲜重和地下部鲜重均有显著(P<0.05,下同)下降,降幅分别为19.57%、14.69%、31.74%和29.04%。在镉胁迫下,喷施谷胱甘肽的处理CdGSH比Cd处理的根长显著升高了22.01%,株高显著提高了6.59%,地上部鲜重显著增加了39.11%,地下部鲜重显著增加了26.47%;喷施谷胱甘肽合成抑制剂BSO可显著降低辣椒地下部鲜重,而根长、株高和地上部鲜重与Cd处理差异不显著;同时喷施GSH和BSO,比单独喷施BSO的处理CdBSO在根长、地上部鲜重和地下部鲜重均有显著提升,而比CdGSH处理均有显著降低。
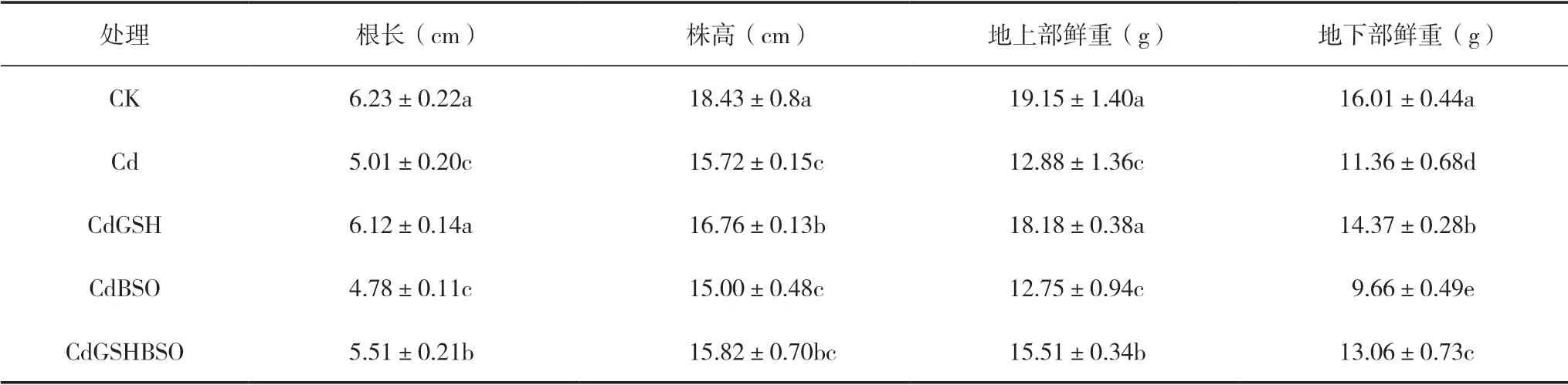
表2 GSH和BSO对镉胁迫下辣椒幼苗生物量的影响
2.2 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗叶片光合色素含量的影响
光合色素的含量可以作为衡量植物对重金属胁迫耐受性的一个重要指标。由表3可知,与CK相比,处于镉胁迫下Cd处理的叶绿素a、叶绿素b、类胡萝卜素含量和叶绿素a/叶绿素b均显著降低,分别降低了48.36%、35.03%、40.13%和18.64%。在镉离子胁迫下,施加GSH与Cd处理相比,辣椒幼苗叶片的叶绿素a含量显著提高了82.65%,叶绿素b显著提高了51.33%,类胡萝卜素含量显著提高了32.19%,叶绿素a/叶绿素b显著提高了20.70%。喷施谷胱甘肽合成抑制剂BOS后,叶部的3种光合色素含量较Cd处理均有显著下降。同时喷施GSH和BSO,比单独喷施BSO的CdBSO处理,叶片的绿素a、叶绿素b、类胡萝卜素含量均有显著提升。
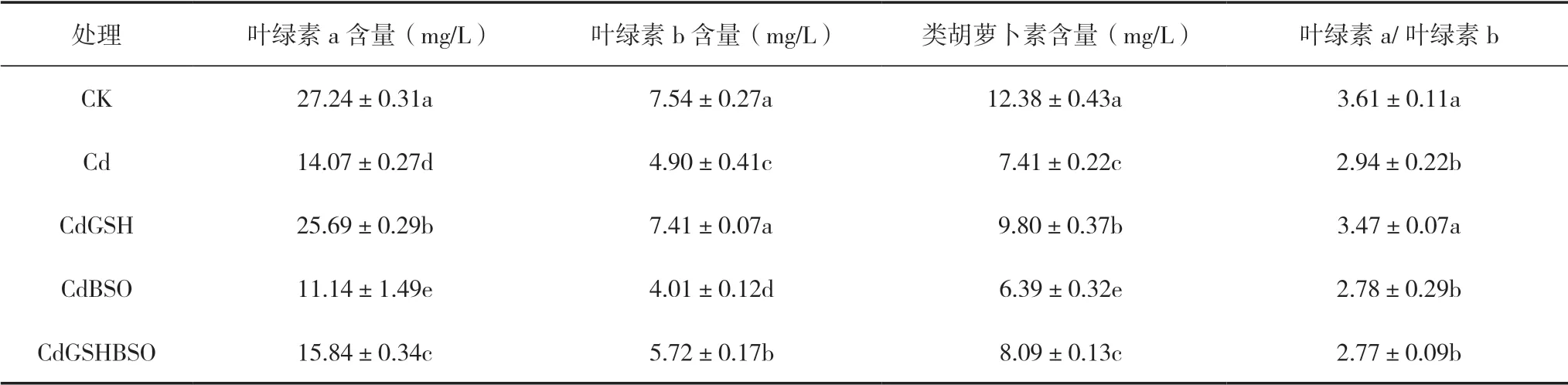
表3 GSH和BSO对镉胁迫下辣椒幼苗光合色素的影响
2.3 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗镉含量和转运系数的影响
由表4可知,各个处理的叶部镉含量均小于根部的镉含量。在镉胁迫下喷施GSH后,辣椒幼苗叶部和根部的镉含量均比Cd处理显著升高,增幅分别为11.26%和33.26%,转运系数有所降低,但差异不显著。在喷施BSO后,叶部镉含量比Cd处理显著下降了12.82%,而根部则降低了10.53%。同时喷施GSH和BSO,比单独喷施BSO的CdBSO处理,叶片和根部镉含量分别显著提升了16.34%和35.56%,而比CdGSH处理叶片和根部镉含量分别显著降低了10.00%和18.94%。

表4 GSH和BSO对镉胁迫下辣椒幼苗镉含量和转运系数的影响
2.4 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗抗氧化酶活性的影响
SOD、CAT、APX和POD是植物重要的抗氧化酶,在植物体清除ROS的过程中起到重要作用。由图1可知,辣椒叶部的4种抗氧化酶活性总体上均高于根部的酶活性。与CK相比,Cd处理根部的SOD活性以及根部和叶部的CAT、APX和POD活性均显著下降。在镉胁迫下,喷施GSH对根部的SOD、CAT、APX和POD活性均显著提升,比Cd处理分别增加了49.63%、68.15%、51.60%和59.72%;喷施GSH对叶部的CAT、APX和POD酶活性均显著提升,相比于Cd处理分别有28.11%、64.86%和40.47%的增幅。在喷施BSO后,辣椒叶部SOD、CAT和POD活性均显著下降。同时喷施GSH和BSO,叶部的CAT、APX和POD酶活性和根部的SOD、CAT、APX、POD酶活性比CdBSO处理均显著提升。
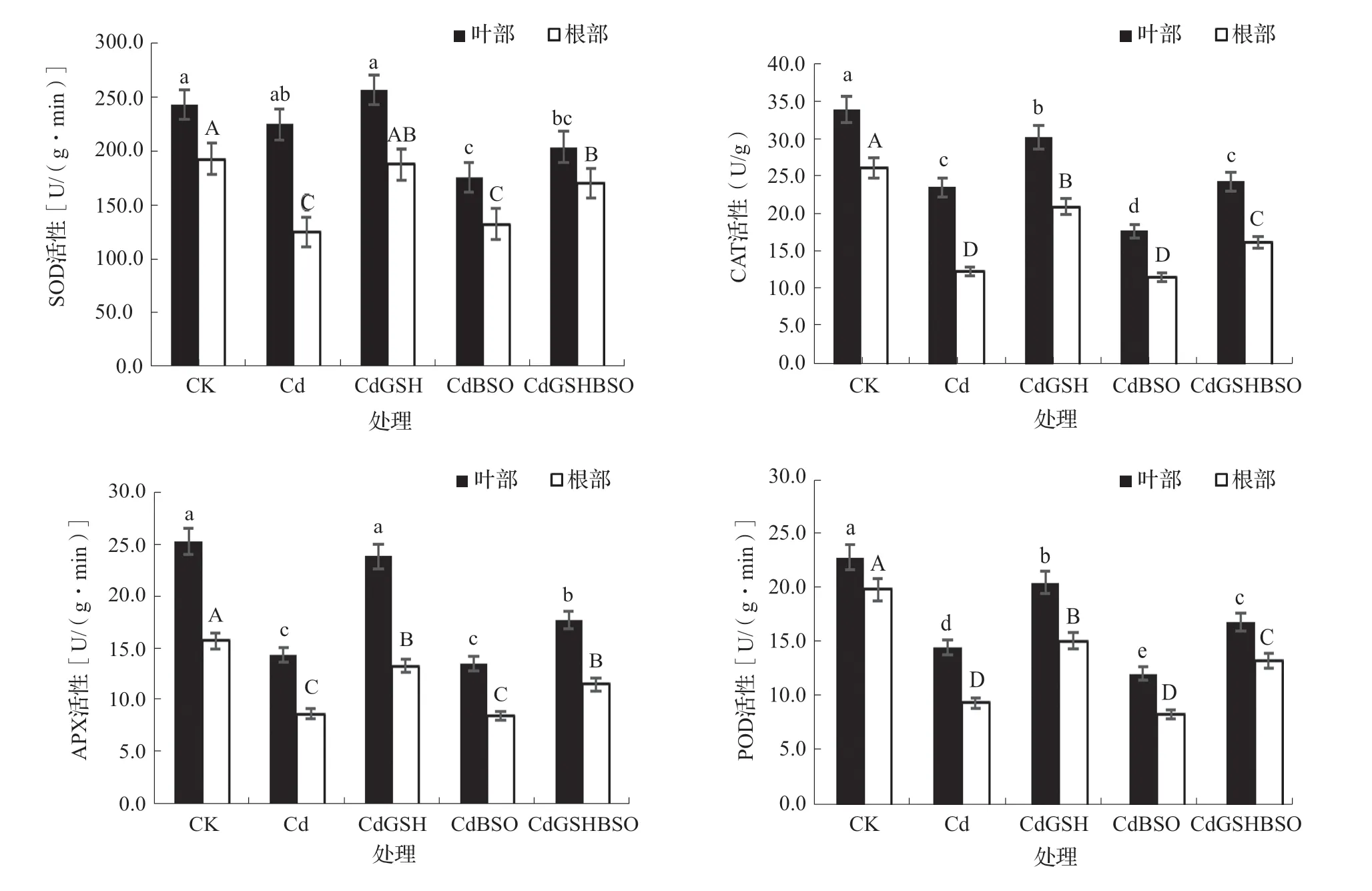
图1 谷胱甘肽和丁硫堇对镉胁迫下辣椒抗氧化物酶活性的影响
2.5 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗AsAGSH循环关键酶活性的影响
AsA-GSH循环是除了抗氧化物酶系统以外,另一套植物体清除ROS的重要系统,GR、DHAR、MDHAR、GPX和γ-GCS是该系统的关键酶。由图2可知,与CK相比,Cd处理根部和叶部的GR、DHAR、MDHAR和γ-GCS活性均显著降低,而GPX活性则显著升高。在镉胁迫下,补充GSH可显著提升GR、DHAR、MDHAR和GPX在辣椒叶片和根部的活性,而γ-GCS活性在叶部降低,在根部则与Cd处理差异不显著。在施加了BSO后,叶部的GR、DHAR、GPX、γ-GCS活性和根部的MDHAR、DHAR、GPX、γ-GCS活 性 与Cd处理相比均显著降低。同时喷施GSH和BSO,叶部和根部的GR、DHAR、MDHAR、GPX酶活性比CdBSO处理均显著提升。
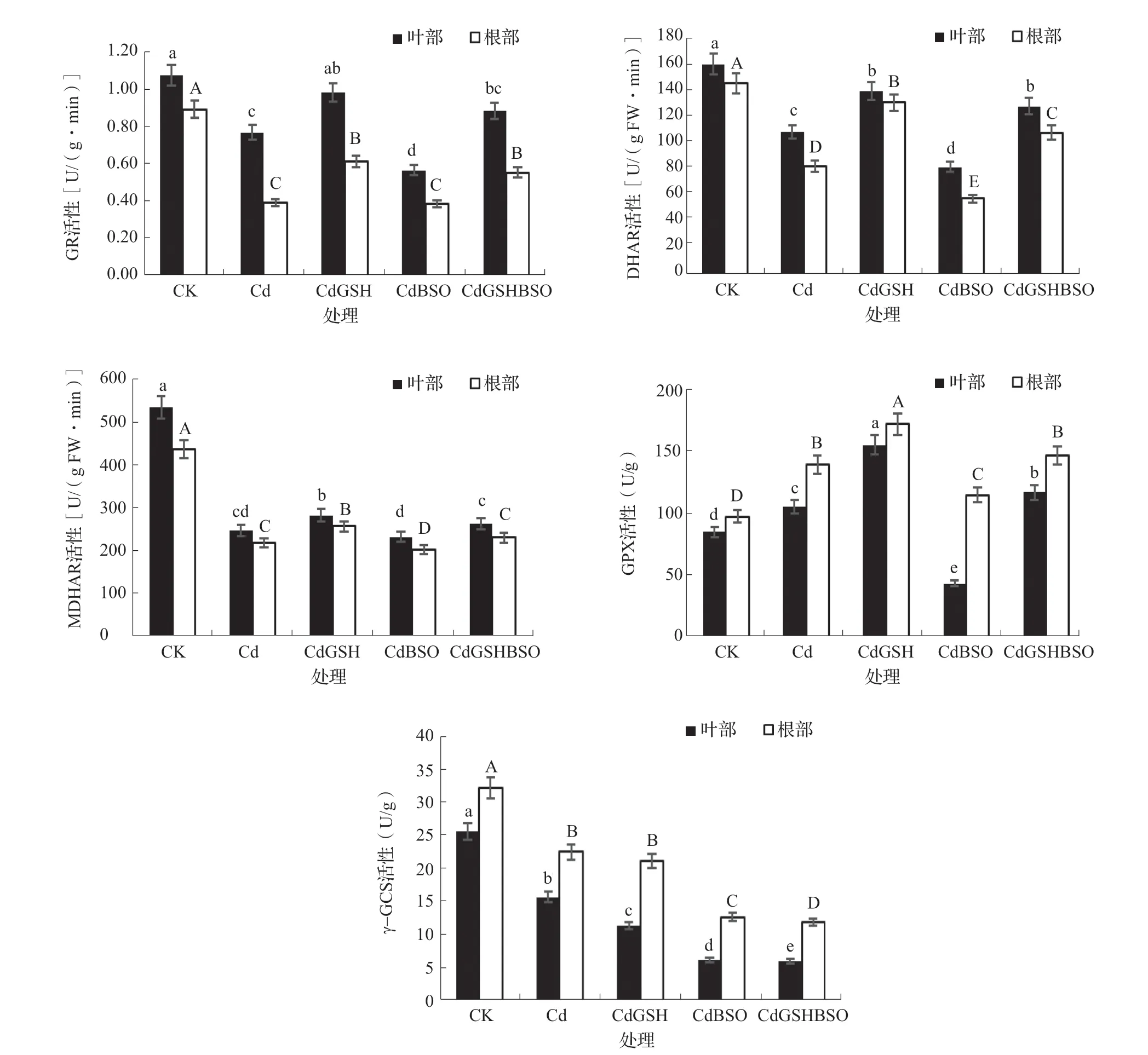
图2 谷胱甘肽和丁硫堇对镉胁迫下辣椒AsA-GSH循环关键酶活性的影响
2.6 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗GSH循环的影响
GSH是植物体内清除ROS的重要还原性物质,GSH/GSSG代表了细胞内还原氧化性物质的能力,与植物抵抗非生物胁迫能力相关。由图3可知,Cd处理比CK处理根部和叶部的GSH、GSSG含量显著升高,叶部GSH/GSSG显著降低,而根部GSH/GSSG变化不显著。相比于Cd处理,CdGSH处理可显著提升镉胁迫下的辣椒叶片和根部GSH、GSSG含量和GSH/GSSG,增幅分别为165.83%、62.62%、37.18%和15.89%、93.78%、40.32%。相比于Cd处理,CdBSO处理可显著降低辣椒叶部和根部GSH、GSSG含量以及叶部的GSH/GSSG。同时喷施GSH和BSO,叶部和根部GSH、GSSG含量和GSH/GSSG比CdBSO处理均显著提高;与CdGSH处理相比,CdGSHBSO处理叶部的GSH、GSSG含量和GSH/GSSG均显著下降,根部只有GSH显著下降。

图3 谷胱甘肽和丁硫堇对镉胁迫下辣椒GSH循环的影响
2.7 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗ROS的影响
O2·-和H2O2是植物体内主要的ROS,O2·-产生速率和H2O2含量可代表植物细胞内ROS的积累程度。由图4可知,与CK相比,Cd处理叶部和根部的O2·-产生速率分别显著下降44.01%和60.63%,H2O2含量在叶部和根部分别显著下降53.83%和119.98%。在镉胁迫下,喷施GSH后,叶部和根部的O2·-产生速率比Cd处理分别降低27.92%和23.99%,H2O2含量分别降低24.99%和31.81%。在喷施BSO后,叶部和根部的O2·-产生速率和根部的H2O2含量均显著升高。同时喷施GSH和BSO,叶部和根部的O2·-产生速率和H2O2含量比CdBSO处理均显著降低,而与CdGSH处理相比则显著上升。
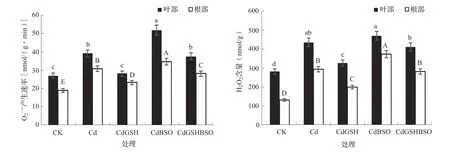
图4 谷胱甘肽和丁硫堇对镉胁迫下辣椒O2·-产生速率和H2O2含量的影响
3 讨论
3.1 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗生物量和光合色素的影响
镉胁迫会导致植物体内过量的ROS迅速积累,影响光合色素的稳定性,阻碍光合作用对能量的转化,破坏营养物质的正常积累,从而严重影响植物生物量的增长[16]。本研究结果表明,在镉胁迫条件下辣椒幼苗根长、株高、地上部鲜重和地下部鲜重以及叶绿素a、叶绿素b和类胡萝卜素含量明显下降,在叶面喷施GSH则可有效提升镉胁迫下辣椒生物量和光合色素的含量,而喷施GSH合成抑制剂BSO则会加重镉胁迫下辣椒生物量和光合色素含量的下降。正面和反面的结果共同说明GSH可以缓解镉胁迫辣椒幼苗生长发育的抑制和对光合色素的降解,有效保护辣椒光合系统,从而提升幼苗对镉胁迫的耐受性。
3.2 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗镉含量和转运系数的影响
GSH可通过聚合反应形成多聚物骨架,进而与重金属阳离子发生螯合形成螯合肽,在重金属吸收和转运方面起着重要作用[17-18]。本研究结果表明,在叶面喷施GSH可增加叶部和根部的镉含量,其中根部镉含量的增加更为明显,因此转运系数反而下降;喷施BSO可同等程度地降低叶部和根部的镉含量,因此转运系数变化不显著。Yuan等[19]的研究表明,对马蔺喷施GSH会显著增加根部和叶部的铅含量,这与本研究结果相一致,但马蔺对铅的转移系数会因GSH而增加,这与本研究结果不同。Nakamura等[20]的研究结果表明,对油菜喷施BSO可降低根中镉含量,但会增加油菜茎叶中的镉含量,也与本研究结果部分不同,这可能是植物种类、重金属种类和施用方式不同导致的。喷施GSH显著增加了辣椒根部和叶部的镉含量,但却没有对辣椒的生长和光合作用造成损害,反而增加了辣椒的生物量和光合色素含量。这可能是由于GSH通过螯合作用将原本可自由移动的镉离子固定为金属螯合肽,螯合物可被转运蛋白运送至细胞外或储存于液泡内。辣椒镉元素的总含量虽然在上升,但是可对植物细胞产生胁迫的活性镉的有效浓度却明显下降,因此减轻了细胞膜的过氧化程度,保护了光合色素的稳定性,维护了营养物质的正常积累。
3.3 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗抗氧化物酶的影响
植物体内清除O2·-和H2O2等ROS的机制可分为抗氧化酶和非酶2个清除系统:抗氧化酶系统的关键酶为POD、SOD、CAT、APX;非酶系统又称为AsA-GSH循环系统[21]。SOD可将O2·-催化分解为H2O2和O2,而H2O2可分别由POD催化还原型辅酶代谢以及APX催化AsA代谢,或者是CAT直接清除,因而抗氧化酶在响应重金属胁迫中具有重要作用[22]。本研究结果表明,镉胁迫会造成辣椒幼苗的抗氧化物酶活性降低,这可能是镉离子替换了抗氧化酶活性中心的镁、铁和锰等金属阳离子,造成部分酶的功能丧失。对镉胁迫下的辣椒幼苗喷施外源GSH后,抗氧化物酶活性均显著提高,而添加BSO则会抑制这些酶的活性,从正反两方面说明GSH可有效保护抗氧化物酶行使正常功能,降低镉胁迫对酶活性的影响。
3.4 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗AsAGSH循环及其关键酶活性的影响
AsA和GSH可直接还原ROS,氧化产物分别为DHA和GSSG。GR则可将氧化态的GSSG还原为GSH[23],MDHAR和DHAR酶可分别将单脱氢抗坏血酸(MDHA)和DHA还原为AsA[24],GPX以GSH为电子供体还原ROS,γ-GCS是植物体内GSH合成路径上游的关键酶[25-28]。本研究结果表明,在镉胁迫下,辣椒根部和叶部的GR、DHAR、MDHAR和γ-GCS活性以及GSH/GSSG均显著降低,而GPX活性以及GSH、GSSG含量则显著上升。同时本研究结果也表明,补充GSH可显著提升辣椒叶片和根部GR、DHAR、MDHAR和GPX的活性以及GSH、GSSG含量和GSH/GSSG,而γ-GCS在叶部的活性则随着GSH的增加而降低;添加BSO可不同程度地降低MDHAR、GR、DHAR、GPX和γ-GCS活性以及GSH、GSSG含量和GSH/GSSG。结果从正反两方面说明GSH可通过激活AsA-GSH循环系统关键酶活性来提升AsA-GSH的循环速率,通过提高GSH循环系统还原态/氧化态比例来加快清除ROS效率。
3.5 谷胱甘肽和丁硫堇对镉胁迫下辣椒幼苗ROS的影响
在非胁迫状态下,植物细胞内的O2·-和H2O2等ROS的产生和清除处于动态平衡状态,而过多的镉离子会打破该平衡状态[29]。本研究结果表明,在镉胁迫下,辣椒幼苗叶部和根部O2·-产生速率和H2O2含量均显著升高,喷施GSH可有效清除辣椒体内的ROS,而BSO则会加剧ROS的积累。结合上文的结果不难得出,镉胁迫下辣椒体内的抗氧化酶和非酶清除系统受到抑制,造成了体内ROS的大量积累,而补充GSH则一方面可直接参与ROS的清除,另一方面可通过提高抗氧化酶系统和AsA-GSH循环系统的ROS清除效率,有效缓解镉离子造成的细胞ROS胁迫效应。
4 结论
本研究以在高浓度镉胁迫下的晋椒503为试验材料,探究镉胁迫下GSH和BSO对辣椒幼苗叶部和根部各项生理性指标的影响。研究结果表明,高浓度镉离子对辣椒的胁迫主要表现在抑制抗氧化酶和AsA-GSH循环关键酶活性而造成ROS过度积累,添加外源GSH一方面可直接清除ROS,另一方面可显著提高POD、SOD、CAT、APX、GR、DHAR、MDHAR和GPX酶活性,提高GSH循环系统还原态/氧化态比例,两个路径共同作用降低了O2·-的产生速率和H2O2的含量,缓解了镉对辣椒幼苗的正常生长和光合作用的抑制效应;而添加GSH合成抑制剂BOS则会逆转上述作用,从反面证明了GSH在辣椒响应镉胁迫的过程中起到重要作用。外源GSH虽然增加了辣椒幼苗镉的总含量,但未对辣椒的各项生理指标产生副作用,这可能是由于GSH与镉离子形成了螯合肽,被转运至细胞外和液泡内,降低了叶绿体内和各个酶活动区域的镉离子浓度,从而缓解了镉对于光合作用和ROS清除系统的抑制作用。而在喷施外源GSH和BSO的条件下,辣椒细胞内重金属螯合肽与活性镉离子的具体变化情况还需要进一步研究。
猜你喜欢
农业灾害研究(2022年2期)2022-05-31
长春中医药大学学报(2022年5期)2022-05-24
现代养生·上半月(2021年10期)2021-09-24
西藏文学(2020年5期)2020-09-21
中国诗歌(2019年6期)2019-11-15
饮食与健康·下旬刊(2019年9期)2019-03-08
分析化学(2018年4期)2018-11-02
饮食与健康·下旬刊(2018年3期)2018-04-11
董事会(2016年8期)2016-05-14
小资CHIC!ELEGANCE(2015年24期)2015-12-12
