
全身糖脂代谢的关键调节剂:法尼酯X 受体(FXR)
2022-07-27 09:35王定保丁雪莲蒋祖环叶啟发
武汉大学学报(医学版) 2022年5期
王定保 丁雪莲 蒋祖环 李 玲, 杨 岚 叶啟发,3
1孝感市第一人民医院临床中心实验室 湖北 孝感 432100;2武汉大学中南医院/武汉大学肝胆疾病研究院/武汉大学移植医学中心/移植医学技术湖北省重点实验室 湖北 武汉 430071;3中南大学湘雅三医院/卫生部移植医学工程技术研究中心 湖南 长沙 410013
1 法尼脂X 受体的结构及表达
法尼脂X 受体(farnesoid X receptor,FXR)是核受体超家族和胆汁酸(bile acid,BAs)受体成员之一。FXR 作为单体与类视黄醇X 受体(retinoid X receptor,RXR)结合形成异源二聚体,并结合FXR应答元件,调控众多下游基因的转录。
1995 年,Jiang 等[1]通过质粒转染、序列分析,首次在大鼠的cDNA 序列中分离出一个属于NR1H 核受体亚族的新成员,后被编码为NR1H4,并发现法尼酯可增强其转录活性,因此命名法尼酯受体。后期研究证实胆汁酸代谢产物与其在生理状态下实现结合,在这一过程中胆汁酸是其内源性配体,故又命名胆汁酸受体[2]。
FXR 位 于12 号 染 色 体 长 臂2 区3 带1 亚 带(12q 23.1),长约90 kb,包含11 个外显子。FXR 包括N-端配体非依赖的激活功能结构域1(activation function domain,AF1)、DNA 结 合 结 构 域(DNA binding domain,DBD)、铰链区(hinge)、配体结合域(ligand binding domain,LBD)4 个部分。其中,位于C-端的LBD 包含配体依赖的激活功能结构域2(activation function domain,AF2),参与配体结合和二聚化过程(图1)。
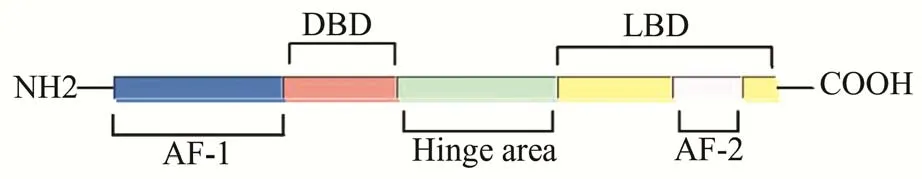
图1 FXR 的结构
研究表明,FXR 主要在肝、肠、肾和肾上腺中大量表达[3,4],通过复杂的信号通路网络,FXR 与机体内相关激素或各代谢途径限速酶相互作用,参与调控机体胆汁酸代谢、脂质代谢、葡萄糖代谢等多种代谢途径,与多种疾病的发生发展有关[5,6],因此,FXR 有望成为相关疾病诊断和治疗的新靶点。
2 FXR 参与正常生理代谢的过程
2.1 FXR 在脂质代谢中的作用Feng 等[7]发现,高甘油三酯血症患者胆汁酸代谢功能障碍,而FXR在胆汁酸代谢调节中起到重要作用,因此我们推测FXR 与甘油三酯代谢有关。鹅去氧胆酸(chenodeoxycholic acid,CDCA)是FXR 的配体,其在临床中常作为胆固醇结石溶解剂发挥作用,可减少胆汁中胆固醇的浓度,亦提示FXR 在脂质代谢中的作用。
2.1.1FXR 调节甘油三酯水平 肝是合成甘油三酯(triglyceride,TG)的主要场所,肝TG 水平由脂肪酸氧化水解和甘油、脂肪酸合成的动态平衡调控。脂肪酸β 氧化主要由过氧化物酶体增殖物活化受体α (peroxisome proliferator-activated receptor-α,PPARα)控制。而甘油、脂肪酸合成TG 的过程受固醇 调 节 元 件 结 合 蛋 白-1(sterol regulatory element-binding protein-1,SREBP-1)调 控。Sagar等[8]证明,FXR 上调PPARα 表达,促进脂肪酸β 氧化分解,使TG 合成原料减少,且通过激活小二聚体伴侣(small heterodimer partner,SHP)介导的信号通路抑制SREBP-1 的表达[9],进一步抑制甘油三酯合成。
Panzitt 等[10]报道,脂肪组织中FXR 通过抑制肿瘤坏死因子α(tumor necrosis factor alpha,TNF-α)、过氧化物酶体增殖物活化受体γ(peroxisome proliferator-activated receptor-γ,PPARγ)、CCAAT/增强子结合蛋白(CCAAT/enhancer binding protein,CEBPα)及脂肪酸结合蛋白(fatty acid binding protein,FABP)的表达,促进脂肪储存,降低血液中TG 浓度。
2.1.2FXR 调节游离脂肪酸水平 研究者对FXR基因敲除小鼠和野生型小鼠同时给予高脂饮食喂养,发现FXR 基因敲除小鼠循环游离脂肪酸(free fatty acids,FFA)水平更高,这是由于FXR 缺陷小鼠中脂蛋白脂肪酶(lipoprtein lipase,LPL)活性高于野生型小鼠[11],从而使乳糜微粒(chylomicron,CM)中TG 及磷脂水解增多,故产生的FFA 增高,提示FXR 在降低循环游离脂肪酸中的作用。在正常非肥胖但无糖尿病大鼠中,FXR 激活能改善其体内的游离脂肪酸分布也得到证实[10]。此外,FXR 还调节多个脂质代谢相关蛋白的基因表达[12],如抑制磷脂转移蛋白(phospholipid transfer protein,PLTP)和载脂 蛋 白(apolipoprotein,apo)C-Ⅱ,以 及apo A-Ⅰ,C-Ⅲ,载 脂 蛋 白E 和 对 氧 磷 酶1(paraoxonase 1,PON1)等,间接调节FFA 水平(图2)。
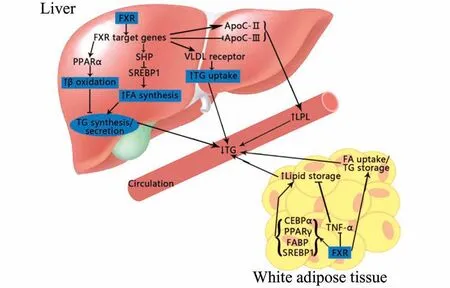
图2 FXR 对脂质代谢的影响
2.2 FXR 在糖代谢中的作用
2.2.1FXR 在糖异生和葡萄糖水平正常化中的作用 胆固醇-7a-羟化酶(cholesterol-7a-hydroxylase,CYP7A1)和糖皮质激素受体(glucocorticoid receptor,GR)是糖异生过程中的关键基因[13]。FXR 激活引起FGF19(成纤维细胞生长因子)表达水平的升高,FGF19 分泌进入门静脉循环,随后在肝脏中与FGFR4 受体结合,活化MAPK 信号通路,进而引起CYP7A1 表达的下调[14],另外FXR 也抑制GR 的表达[15],由此推断FXR 通过抑制糖异生来调节血糖水平。
磷酸烯醇丙酮酸羧激酶(phosphoenolpyruvate carboxykinase,PEPCK)和 葡 萄 糖-6-磷 酸 酶(G6Pase)是肝脏糖异生过程中的主要限速酶,PEPCK 和G6Pase 过表达引起循环葡萄糖水平升高和葡萄糖不耐受,而HNF-4α 可以在转录层面调控编码这两种基因。FXR 通过SHP-LRH1 途径抑制HNF-4α 的表达,在转录水平上抑制糖异生来使血糖水平正常化[14]。
2.2.2FXR 在糖原合成中的作用 葡萄糖通过糖原合酶的连接作用,实现糖原合成及葡萄糖储存的过程。糖原合酶激酶-3β(glycogen synthase kinase-3β,GSK-3β)的磷酸化及其失活直接影响糖原合酶活性(糖原合酶磷酸化后则失去活性)。且GSK-3β 主要通过胰岛素受体-胰岛素受体底物(insulin receptor substrate,IRS)-Akt 途径被胰岛素灭活,而FXR 可诱导Akt 磷酸化[16]。由此我们推测,FXR 可 能 通 过IRS-Akt 途 径 灭 活GSK-3β,减 少 糖原合酶磷酸化,升高其活性来增加葡萄糖糖原合成,降 低 血 浆 葡 萄 糖 浓 度。Katafuchi 等[17]发 现,FXR 治疗能增加小鼠中糖原合成和储存。正常喂养的FXR 基因敲除小鼠对比同等喂养条件野生小鼠,表现明显餐后高血糖,可能与糖异生作用增强和糖原合成抑制有关;而禁食FXR 基因敲除小鼠出现低血糖,提示FXR 可能有助于空腹条件下血糖生成,提示FXR 在血糖浓度调节中可能表现双重作用,取决于机体的病理生理条件,最终保证血糖水平平衡。此外,肝是糖原合成与分解的主要场所,FXR 基因敲除小鼠中肝细胞凋亡增加,血糖调节能力减弱,而经FXR 激动剂CDCA 和GW4064 处理后HepG2 细胞凋亡减少,进一步验证了FXR 在维持体内葡萄糖稳态中的作用[18](图3)。
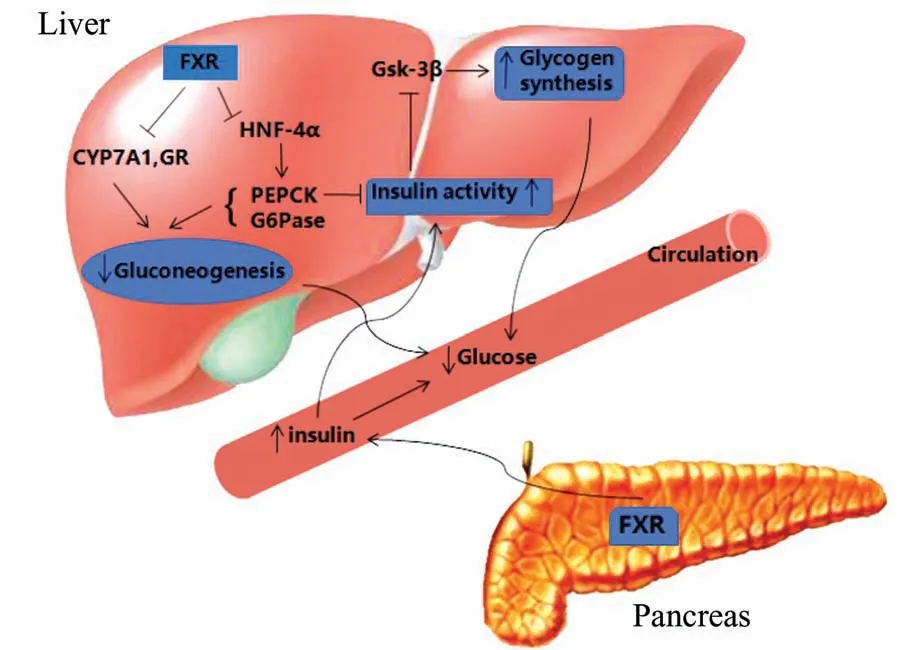
图3 FXR 对葡萄糖代谢的影响
3 FXR 与代谢性疾病的关系
FXR 作为一种重要核受体,它在全身代谢尤其是糖、脂代谢中均发挥了重要作用,因此,FXR 与某些代谢性疾病的发生发展密切相关。
3.1 糖尿病2 型糖尿病作为一种常见的代谢性疾病,其重要特征包括胰岛素抵抗、脂质代谢紊乱等,而FXR 调控胆汁酸代谢与上述特征均有紧密联系。研究表明,胆汁酸(bile acid,BA)治疗有利于缓解2 型糖尿病[19]。Wang 等[20]指出,糖尿病啮齿动物模型中FXR 表达降低。高葡萄糖浓度刺激FXR 表达,而高胰岛素抑制后者的表达[21],提示FXR 在维持机体血糖浓度平衡中的作用。早期有研究发现,FXR 基因敲除小鼠的胰岛素敏感性降低,其FXR缺乏是胰岛素抵抗发生及发展为糖尿病的关键[22]。与此发现一致的是,用FXR 特异性激动剂治疗野生型和两种糖尿病模型(ob/ob 和db/db)小鼠能改善它们的胰岛素敏感性并降低其血浆葡萄糖水平[22]。葡萄糖转运蛋白(glucose transporter 4,GLUT4)是存在于肌和脂肪组织中的转运载体,其作用是以胰岛素依赖方式促进细胞摄取利用血糖,而前者的表达受FXR 调控,若FXR 数量或功能异常,则GLUT4 无法响应胰岛素分布于脂肪和肌组织细胞膜,即引起胰岛素抵抗,阻碍血中葡萄糖进入细胞。
靶组织中脂质含量增加可能是引起胰岛素敏感性降低的关键因素。FXR 基因敲除的小鼠显示脂肪组织异常,外周胰岛素抵抗,PPARα 和β/Δ 激活,以及肝脏和骨骼肌代谢受损[23],而PPARα 过表达进一步导致了胰岛素抵抗增加。缺少FXR 时,肝脏葡萄糖和TG 输出的增加引起骨骼肌中的胰岛素抵抗,进而调节全身糖脂代谢。Kong 等[24]在体内和体外均证明了β 细胞中FXR 能增加了胰岛素释放并保护脂毒性。另有研究报道,FXR 激活能引起极低密度脂蛋白(very low density lipoprotein,VLDL)输出率下降[25],降低脂质水平,从而引起循环葡萄糖水平总体下降,有利于对抗胰岛素抵抗,达到治疗糖尿病的目的。
3.2 非酒精性脂肪肝非酒精性脂肪肝(non-alcoholic fatty liver disease,NAFLD)是一种常见肝脏疾病,主要特征为肝细胞内脂肪滴的积累,并可演化为非酒精性脂肪肝炎(non-alcoholic steatohepatitis,NASH)甚至肝细胞癌[25]。NAFLD 患者FXR 水 平 较 低,TG 合 成 和 肝X 受 体(LXR)及SREBP-1 表达增加,这些因素与患者肝细胞脂肪变性程度有关[26]。最新研究发现,FXR 激活抑制细胞内PERK/EIF2α/ATF4 和CHOP 信号转导,降低了肝脂肪变性中的肝细胞ER 应激,有效减弱蛋氨酸和胆碱缺乏的L-氨 基 酸(methionine-choline-deficient,MCD)饮食和高脂肪饮食(high-fat diet,HFD)喂养小鼠中NAFLD 和代谢紊乱的进展[27]。以上研究均从不同角度证实FXR 在脂肪变性及NAFLD 的发展中的作用。
3.3 肥胖症肥胖症是以脂肪过剩为主要特征的代谢疾病。FXR 作为核受体,在脂肪细胞分化和脂质储存中发挥重要作用。脂肪组织的代谢变化与肥胖及FXR 参与的胰岛素抵抗过程密切相关。胰岛素抵抗的发生发展与脂肪因子的严重改变相关,其特征在于促炎细胞因子的释放增加和来自脂肪组织的抗炎细胞因子释放减少。Seok 等[28]报道,内源性FXR 的配体去氧胆酸(CDCA)可抑制促炎因子[TNF-α,单核细胞趋化蛋白-1(monocyte chemotactic protein 1,MCP-1)、白细胞介素-6(interleukin,IL-6)、脂肪因子Chemerin、纤溶酶原激活物抑制剂1 型(plasminogen-activator inhibitor type 1,PAI-1)、视黄醇结合蛋白4(retinol binding protein 4,RBP4)、抵抗素、脂肪因子vaspin 等的表达。因此,FXR 激动可能通过调节脂肪因子改善肥胖症中的胰岛素抵抗。
4 结语与展望
近期研究报道,FXR 除了作为调节全身能量代谢的传感器的作用外,它还影响各种器官的完整性维持,包括肝脏、肠道、肾脏、白色脂肪组织(white adipose tissue,WAT)和胰腺,可能与调节它们的代谢功能密切相关[29]。FXR 还通过胆汁酸和成纤维细 胞 生 长 因 子-15/19(fibroblast growth factor,FGF-15/19)促进器官间信号传递,特别是肠肝循环,实现对全身组织代谢的广泛影响。
鉴于FXR 对代谢的改善作用,近十年来科学家为发现和优化FXR 激活的潜在候选药物投入大量研究,其中FDA 已批准奥贝胆酸(obeticholic acid,OCA)作为第一个成功上市的FXR 受体激动剂用于治疗原发性胆汁性肝硬化。但存在较大的安全问题,在某些条件下还发现了FXR 活化后不良影响,其NASH 适应证正处于三期临床试验验证阶段[30]。FXR 参与体内代谢机制的因子繁多,机制复杂,作用多方面,可能与组织特异性、疾病类型和状态,病理刺激和营养状态有关。对FXR 的研究应扩大样本量,科学、合理地选取观测指标,探寻FXR 在不同病理生理状态下的用法和剂量,以求做到个体化用药,并结合临床试验实现其临床意义。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
现代临床医学(2021年6期)2021-11-20
昆明医科大学学报(2021年1期)2021-02-07
科学导报(2019年45期)2019-09-23
保健与生活(2019年23期)2019-09-10
家庭医学(2018年2期)2018-07-14
黄河黄土黄种人(2017年11期)2017-11-27
家庭百事通·健康一点通(2017年8期)2017-08-18
成都体育学院学报(2017年1期)2017-02-21
现代养生·下半月(2016年6期)2016-10-21
