
含笑内酯的药理作用及机制研究进展Δ
2022-07-29 02:29莫俊俏黎咏嫦海南省第五人民医院药学部海口57000海南科技职业大学健康科学学院海口577广西中医药大学药学院南宁50200
中国药房 2022年14期
林 景,莫俊俏,周 欣,宋 艳,黎咏嫦(.海南省第五人民医院药学部,海口 57000;2.海南科技职业大学健康科学学院,海口 577;.广西中医药大学药学院,南宁 50200)
含笑内酯是一种来自天然植物的愈创木烷型倍半萜内酯(化学结构见图1),在木兰科植物黄兰的根皮、台湾含笑的根皮中均有分布;含笑内酯也可用小白菊内酯为原料通过化学反应得到,其活性与小白菊内酯相当,但稳定性更高、毒性和生产成本更低,更具有作为临床药物的潜力[1-2]。现代药理学研究表明,含笑内酯在抗肿瘤、抗炎、免疫调节、神经保护等方面疗效显著[3-9]。为了解含笑内酯的药理学研究进展,笔者在中国知网、万方数据、维普网、PubMed、Web of Science 等数据库中组合查询2016年1月-2022年6月发表的相关文献,对含笑内酯的药理作用及其机制进行了梳理和归纳,以期为其相关新药研发和临床应用提供参考。
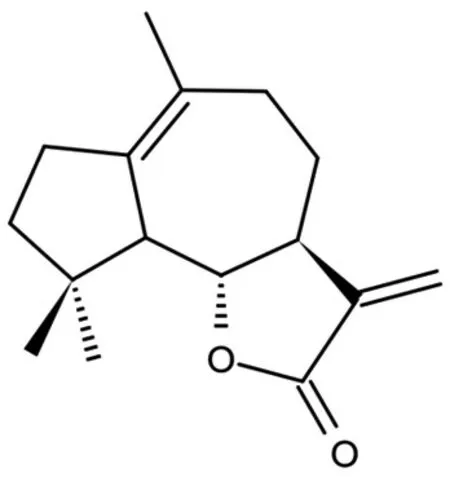
图1 含笑内酯的化学结构图
1 抗肿瘤作用
1.1 抑制肿瘤细胞增殖
研究表明,有氧糖酵解的代谢重编程是癌症的重要标志之一,在糖酵解的最后一步,丙酮酸激酶催化磷酸烯醇-丙酮酸转化为丙酮酸是癌症代谢重编程中至关重要的一步[10]。通常情况下,丙酮酸激酶的M2 亚型(the M2 isoform of pyruvate kinase,PKM2)在单体、二聚体和四聚体形式之间处于平衡状态[11]。Li 等[5]研究发现,含笑内酯可通过共价结合半胱氨酸残基选择性地激活PKM2,促进丙酮酸激酶形成四聚体,降低PKM2的赖氨酸乙酰化和核易位,进而显著抑制白血病细胞的增殖和生长。
相关机制研究表明,含笑内酯还可以从多条途径抑制肿瘤细胞的异常增殖[7,12-14]。例如,白细胞介素6(interleukin 6,IL-6)/信号传导与转录激活子3(signal transducer and activator of transcription 3,STAT3)通路在胃癌的发生和发展中起着重要作用,含笑内酯可通过阻断IL-6/STAT3通路来抑制胃癌AGS和N87细胞增殖[7]。另外,含笑内酯能抑制乳腺癌TNBC 细胞的增殖,并可通过降低细胞内谷胱甘肽水平,增强TNBC 细胞对顺铂的敏感性;同时,含笑内酯对肺癌H1299细胞和Calu-1细胞在辐射和缺氧暴露后缺氧诱导因子1α(hypoxia-inducible factor,HIF-1α)的表达也有明显的抑制作用,其能通过抑制HIF-1α通路使p53 基因缺陷的肺癌细胞对辐射更敏感,从而抑制肺癌细胞异常增殖[12-14]。
1.2 诱导肿瘤细胞凋亡
含笑内酯主要通过线粒体相关途径诱导肿瘤细胞凋亡。据报道,肌动蛋白可通过细胞骨架驱动的运输系统促进线粒体从细胞质中招募各种促凋亡蛋白,从而促进细胞凋亡的启动[15]。Yu等[16]发现,含笑内酯可通过促进线粒体活性氧(reactive oxygen species,ROS)和胱天蛋白酶3(caspase-3)的活化来诱导肝癌细胞凋亡;同时该研究还发现,经含笑内酯处理后的肝癌细胞中线粒体的聚集和F-肌动蛋白纤维的扰动均发生在线粒体ROS活化和肝癌细胞凋亡之前,这表明F-肌动蛋白纤维的扰动可能与线粒体受损和肝癌细胞凋亡有关,这可能是含笑内酯诱导肝癌细胞凋亡的潜在机制之一。Jia 等[6]发现,含笑内酯在抑制人乳腺癌MCF-7 细胞的凋亡过程中伴随着线粒体裂变增加和动力相关蛋白1(dynamin-related protein 1,Drp1)的上调——在含笑内酯的作用下,Drp1的表达水平增加,导致线粒体分裂及ROS生成增加、线粒体膜电位丧失、细胞色素C释放增加,进而放大了乳腺癌细胞的凋亡信号,这表明含笑内酯诱导乳腺癌细胞凋亡的机制可能与Drp1作用于ROS-线粒体凋亡途径有关。还有一些研究发现,含笑内酯还可通过下调肿瘤标志物糖类抗原199(carbohydrate antigen 199,CA199)、CA242、癌胚抗原(carcinoembryonic antigen,CEA)、铁蛋白和甲胎蛋白(alpha fetoprotein,AFP)的表达水平,进一步诱导肝癌H22细胞凋亡;此外,含笑内酯还可通过抑制核因子κB(nuclear factor-kappa B,NF-κB)信号通路、提高caspase-9 蛋白表达水平来诱导卵巢癌HeyA8细胞凋亡[17-18]。
1.3 诱导肿瘤细胞自噬
自噬是机体利用溶酶体降解自身产生的有害物质并维持细胞稳态的自我清洁修护过程[19]。齐春胜等[20]发现,含笑内酯可提高人结肠癌HCT116细胞的自噬率,显著上调STAT3 的磷酸化水平;同时,该研究采用拉马克遗传算法计算出含笑内酯与STAT3 蛋白在体内可直接结合,进一步证实含笑内酯可通过调控STAT3 通路来诱导人结肠癌HCT116细胞发生自噬。
1.4 抑制肿瘤细胞的侵袭和转移
肿瘤细胞的侵袭和转移是一个多步骤、多因素参与的复杂过程。Ji 等[21]发现,含笑内酯可选择性地清除白血病细胞,上调核因子E2 相关因子2(nuclear factor E2-related factor 2,Nrf2)、白血红素加氧酶1(heme oxygenase 1,HO-1)等转录因子的水平,通过抑制NF-κB 信号通路和增加ROS 的产生来抑制白血病细胞的转移。此外,Alwaseem等[22]使用一种新的指纹识别策略诱导了含笑内酯支架上多个碳氢键位置的选择性羟基化,结果发现,羟基化后的含笑内酯可通过靶向重组人过氧化物酶和重组人含硫氧还蛋白结构域蛋白12(thioredoxin domain-containing protein 12,TXNDC12)来抑制白血病细胞的扩散和转移,表现出良好的抗白血病细胞活性。
综上,现有研究表明,含笑内酯对于白血病细胞、胃癌细胞、乳腺癌细胞、肺癌细胞、肝癌细胞等有较强的抑制增殖和促凋亡作用,其作用机制包括抑制肿瘤细胞增殖、诱导肿瘤细胞凋亡、自噬以及抑制肿瘤细胞侵袭和转移;同时,含笑内酯能够调控PKM2/STAT3/NF-κB 等多条通路,提高肿瘤细胞中与机体生长凋亡调控相关的蛋白表达水平,进一步诱导肿瘤细胞凋亡。其良好的抗肿瘤活性对于肿瘤预防以及靶向治疗具有重要意义,值得进一步研究。
2 抗炎作用
2.1 NF-κB信号传导途径
NF-κB是B淋巴细胞的细胞核中的转录因子,能够控制DNA 转录、细胞因子产生和细胞存活。通常情况下,NF-κB 在大多数细胞中均表现为未激活,在细胞质中多与NF-κB 抑制蛋白(inhibitor of NF-κB,IκB)结合;当受到外界刺激后,其可介导多种细胞因子参与炎症反应[23]。研究发现,含笑内酯能够抑制强直性脊柱炎(ankylosing spondylitis,AS)模型小鼠血清中肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)、IL-1 β、IL-6、IL-18 和γ-干扰素(interferon-γ,IFN-γ)的含量,降低caspase-1 p10、IL-1β p17、核苷酸结合寡聚化结构域样受体蛋白3(nucleotide-binding oligomerization domain-like receptor protein 3,NLRP3)和caspase-3的蛋白表达水平;同时抑制磷酸化p65(p-p65)的表达,增加磷酸化IκBα(p-IκBα)的表达[24]。这表明,含笑内酯能够通过抑制NLRP3炎症小体激活和调节NF-κB信号通路、维持Th1/Th2 的平衡来缓解AS。脂多糖(lipopolysaccharide,LPS)是革兰氏阴性菌细胞壁外壁的组成成分,能够通过增加TNF-α和IL-6 等炎症细胞因子的分泌引发炎症反应[25]。Lei等[8]研究发现,含笑内酯能够显著抑制LPS诱导的NF-κB 信号通路激活,降低NLRP3 和caspase-1 的表达水平,减少IL-1β和IL-18 的分泌;同时,含笑内酯还能抑制线粒体ROS(mROS)激动剂鱼藤酮诱导的NLRP3 炎症小体激活,这表明含笑内酯可以通过抑制mROS/NF-κB/NLRP3 通路来抑制LPS 诱导的大鼠肾小管上皮细胞炎症反应。
2.2 PI3K/AKT信号传导途径
磷脂酰肌醇-3-羟激酶(phosphatidylinositol-3-kinase,PI3K)/蛋白激酶B(protein kinase B,Akt)信号通路参与调控细胞凋亡和细胞自噬,在细胞分化、增殖、凋亡及葡萄糖转运等多种细胞转录和迁移过程中发挥关键作用,与炎症相关疾病有着密切联系[26-27]。结核分枝杆菌(Mycobacterium tuberculosis,MTB)在结核病活动期能够引起患者肉芽肿性肺部病变和全身炎症反应,研究表明,含笑内酯可减少MTB 所致小鼠单核巨噬细胞白血病细胞中炎症细胞因子(IL-1β、TNF-α)的异常分泌,显著抑制诱生型一氧化氮合酶(inducible nitric oxide synthase,iNOS)和环氧合酶2(cyclooxygenase 2,COX2)的激活以及随后一氧化氮(NO)的产生,其机制可能与通过PI3K/AKT/NF-κB通路来调控MTB诱导的炎症反应、抑制NLRP3炎症小体的激活有关[28]。Qin等[29]的研究进一步证实了含笑内酯能够抑制LPS 造成的小鼠急性腹膜炎,能降低IL-6、TNF-α、IL-1β、单核细胞趋化蛋白1(monocyte chemotactic peptide-1,MCP-1)、IFN-β 和IL-10 的分泌,其抗炎作用可能是通过抑制PI3K/Akt/p70S6K 和NF-κB 信号通路的激活来发挥的。此外,含笑内酯在金黄色葡萄球菌引起的小鼠腹膜炎中也能够发挥显著的抗炎作用,可抑制巨噬细胞和树突状细胞中炎症细胞因子(IL-6、TNF-α)和趋化因子(MCP-1、IFN-γ)的表达,通过调控PI3K/Akt和NF-κB信号通路来降低金黄色葡萄球菌引起的炎症反应,进而改善肝、肾等器官损伤[30]。
可见,含笑内酯在治疗AS 等炎症相关疾病上具有良好的疗效,能够通过调控NF-κB、PI3K/Akt 等多条炎症相关信号通路来减轻机体的炎症反应,提示含笑内酯具有开发成为抗炎药物的潜力。
3 免疫调节作用
类风湿关节炎(rheumatoid arthritis,RA)是一种慢性自身免疫性疾病,其特征是关节滑膜炎和周围关节的侵蚀性破坏[31]。补体组分5a(complement component 5a,C5a)在先天性免疫反应中可与巨噬细胞、中性粒细胞、T细胞等免疫细胞表面结合,通过丝裂原活化的蛋白激酶途径启动G 蛋白偶联信号,从而诱导炎症细胞因子(如TNF-α、IL-6)的合成[32]。巨噬细胞集落刺激因子(macrophage colony-stimulating factor,M-CSF)由成骨细胞、滑膜细胞和牙周成纤维细胞产生,其与NF-κB配体受体活化相结合,可调节破骨细胞分化,参与RA等加重骨质流失的慢性炎症过程[33]。有学者发现,含笑内酯能恢复关节炎模型小鼠M-CSF、金属蛋白酶组织抑制因子1(tissue inhibitors of metalloproteinase 1,TIMP-1)和C5/C5a的水平,并可以增加B 淋巴细胞趋化因子(B lymphocyte chemoattractant 1,BLC)的表达水平,表明含笑内酯可通过恢复C5/C5a、M-CSF、TIMP-1、BLC 等因子的水平来发挥免疫调节作用[34-35]。
4 神经保护作用
阿尔茨海默病(Alzheimer disease,AD)是最常见的中枢神经系统退行性疾病,由淀粉样前体蛋白异常加工而来的β淀粉样蛋白(amyloidβ-protein,Aβ)的积累是AD患者脑部常见的病理特征。小胶质细胞的活化与帕金森、AD 等神经退行性疾病的发病机制密切相关[36]。现代研究表明,含笑内酯能明显减少转基因AD 模型小鼠的海马齿状回、CA1区和皮质区的Aβ斑块数量、斑块面积以及小胶质细胞的数量,且能抑制小胶质细胞的异常活化和聚集;同时,也能减少胶质纤维酸性蛋白星形胶质细胞的数量、抑制星形胶质细胞的活化,并能够明显改善转基因AD模型小鼠的认知障碍[4]。
此外,小胶质细胞参与的神经炎症被认为在各种神经退行性疾病中可以诱导脑损伤,抑制小胶质细胞的过度激活可能对神经退行性疾病的治疗有益[37]。Sun等[38]发现,含笑内酯可显著减轻LPS刺激BV2小鼠小胶质细胞引发的神经炎症,降低iNOS、COX-2、TNF-α、IL-6 和NO 的含量,其机制可能与含笑内酯能够抑制IκBα/NF-κB 通路、Akt 通路的激活以及c-Jun 氨基末端激酶(c-Jun amino terminal kinase,JNK)、p38 丝裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)激酶和细胞外信号调节激酶1/2(extracellular signal-regulated kinase 1/2,ERK1/2)的激活有关;同时,含笑内酯能够通过增强Nrf2的活性,显著促进抗氧化蛋白HO-1的表达,这表明含笑内酯可在神经炎症引发的神经退行性疾病中发挥神经保护的作用。
可见,含笑内酯可通过减少Aβ的积累,抑制小胶质细胞活化等作用来改善神经炎症反应,可通过调节IκBα/NF-κB、Akt、JNK、MAPK、ERK1/2通路及增强Nrf2的活性来发挥治疗神经退行性疾病的作用。
5 肝脏保护作用
非酒精性脂肪性肝病(non-alcoholic fatty liver disease,NAFLD)是糖尿病、肝纤维化和心血管疾病发展的新危险因素。在NAFLD 进展过程中,炎症调节异常是导致脂质通量失衡的主要驱动因素,而脂肪酸代谢稳态受损又加重了肝脏和整个系统的炎症反应[39]。有研究表明,含笑内酯可减轻糖尿病模型小鼠的肝脏脂肪变性,降低血脂水平(三酰甘油、总胆固醇)、肝脏转氨酶水平(丙氨酸转氨酶、天冬氨酸转氨酶)以及炎症细胞因子(TNF-α、IL-1β)的异常升高,上调肝细胞过氧化物酶体增殖物激活受体(peroxisome proliferator-activated receptor γ,PPAR-γ)的含量以及降低p-IκBα和p-NF-κB/p65 的表达水平,从而抑制NF-κB 通路,减少肝细胞中TNF-α和IL-1β的产生[40]。
自噬在改善脂质代谢和肝脏脂肪变性中也同样发挥着重要作用[41]。PPAR-γ是一种主要的核受体,能够调节脂质代谢和增加脂质分解,含笑内酯可通过激活腺苷酸活化蛋白激酶(adenosine monophosphate activated protein kinase,AMPK)/雷帕霉素靶蛋白抑制剂(mammalian target of rapamycin inhibitors,mTOR)信号通路来上调PPAR-γ的表达,诱导糖尿病模型小鼠的肝细胞发生自噬[40]。
可见,含笑内酯不仅可以减轻肝细胞的炎症反应和脂肪变性,还可以诱导肝细胞自噬,进一步发挥其肝脏保护作用。
6 肾脏保护作用
肾纤维化是慢性肾脏疾病向终末期肾病进展的最常见特征,这一过程包括肾脏固有细胞损伤、细胞外基质的过度积累以及肌成纤维细胞和成纤维细胞活化增殖等,肾脏固有细胞在损伤过程中发生上皮间充质转化(epithelial-mesenchymal transition,EMT)是肾纤维化的重要机制[42]。现有研究表明,异黏蛋白(metadherin,MTDH)是一种参与肾脏纤维形成的因子,含笑内酯能够特异性抑制MTDH 过表达所诱导的EMT。在2 种肾纤维化小鼠模型(单侧输尿管阻塞模型和缺血再灌注损伤模型)中,含笑内酯能降低模型小鼠纤维连接蛋白和α-平滑肌肌动蛋白(α-smooth muscle actin,α-SMA)的表达水平,并能在一定程度上回调上皮钙黏素(E-cadherin)的表达水平[9,43]。骨形态发生蛋白(bone morphogenetic protein,BMP)信号是另一种参与纤维形成的细胞外信号,MTDH的过表达可通过调控肾小管上皮细胞中BMP/MAPK 通路来发挥促纤维化作用,而含笑内酯可特异性逆转这种作用[9,44]。可见,含笑内酯能够通过抑制MTDH/BMP/MAPK通路来治疗肾纤维化。
糖尿病肾病是全球终末期肾病的主要类型之一[45]。已有研究表明,含笑内酯可以保护瘦素受体基因缺陷小鼠(已被广泛用作糖尿病模型)肾脏免受肾衰竭和炎症的影响,其机制可能与含笑内酯可抑制MTDH的异常上调有关;同时,MTDH 的下调显著抑制了NF-κB信号通路的激活,并降低了其下游炎症细胞因子(如MCP-1、TNF-α、IL-1β、IL-6等)的表达水平;含笑内酯还可通过抑制MTDH 转录和促进泛素介导的降解来下调MTDH的表达[46]。以上研究结果表明,含笑内酯可通过抑制MTDH/NF-κB通路来减轻肾脏炎症,起到预防糖尿病肾损伤的作用。
综上,含笑内酯可通过抑制MTDH/BMP/MAPK、MTDH/NF-κB等信号通路来减轻肾纤维化,减轻肾脏炎症,从而发挥肾脏保护的作用。
7 心脏保护作用
阿霉素是一种蒽环类抗肿瘤药物,临床用于治疗实体肿瘤和造血系统恶性肿瘤[47]。现有研究发现,阿霉素在低剂量时仍可能产生心脏毒性,剂量过高可能会出现充血性心力衰竭,对心脏组织产生有害影响,包括ROS的产生、线粒体的生物能量衰竭、内质网应激、钙稳态受损、NF-κB 信号通路激活等[48-49]。Kalantary-Charvadeh等[50]发现,含笑内酯对阿霉素诱导的心脏毒性模型小鼠具有保护作用,可有效降低肌酸激酶同工酶MB和心肌肌钙蛋白Ⅰ水平,改善心脏组织结构及心脏卒中体积;同时,含笑内酯能够降低NF-κB p65 亚基的活性以及PI3K、磷酸化Akt 和caspase-3 的水平,减少炎症细胞因子的分泌以及氧化应激造成的损伤,其心脏保护作用机制可能与调节PI3K/Akt/NF-κB信号通路有关。可见,含笑内酯在心脏保护作用方面也具有潜在的应用前景。
8 结语
含笑内酯作为一种三环结构的倍半萜内酯类化合物,已被证实具有多方面的药理活性,包括抗肿瘤、抗炎、免疫调节、神经保护、肝脏保护、肾脏保护以及心脏保护等,也因此得到众多科研工作者的研究与关注。有关其药理活性对应的作用机制也得到初步的明确:含笑内酯可通过抑制肿瘤细胞增殖、调节肿瘤细胞凋亡、诱导肿瘤细胞自噬、抑制肿瘤细胞的侵袭和转移等发挥抗肿瘤作用;其可通过调控NF-κB、PI3K/Akt 等多条相关信号通路,降低炎症细胞因子的表达水平;其可恢复机体免疫相关因子表达,发挥免疫调节作用;其可通过减少Aβ的积累、抑制小胶质细胞活化以及调节IκBα/NF-κB、Akt、JNK、MAPK、ERK1/2 通路及增强Nrf2 的活性来发挥神经保护作用;其肝脏保护作用机制包括通过调节AMPK/mTOR 信号通路来上调PPAR-γ的表达等;其肾脏保护作用机制与其可通过调控MTDH/BMP/MAPK、MTDH/NF-κB通路来减轻炎症反应有关;其心脏保护作用机制包括减轻炎症反应,降低氧化应激水平,调控PI3K/Akt/NF-κB信号通路等。
可见,含笑内酯可通过多条途径发挥药理活性,即与多靶点蛋白、多通路、多途径调控有关,如含笑内酯能通过调控PKM2/STAT3/NF-κB多条信号通路,以及参与ROS-线粒体凋亡、自噬等多条途径发挥抗肿瘤作用;可通过参与调控PI3K/Akt/NF-κB 多条通路来抑制机体的炎症反应。目前,对于含笑内酯抗肿瘤作用的研究多集中在体外分子机制方面,缺乏动物体内实验的证实;对于抗炎、神经保护等作用的研究则主要以探究相关信号通路为主,今后可从多靶点角度出发深入挖掘含笑内酯的药效机制。
综上,含笑内酯是一种具有重要开发价值的化合物,后续可继续深入研究其药效机制,以便为其在治疗肿瘤、AS等复杂疾病中的应用提供参考。
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
中华实用诊断与治疗杂志(2022年1期)2022-08-31
九江学院学报(自然科学版)(2022年2期)2022-07-02
现代食品科技(2022年5期)2022-05-30
新世纪智能(数学备考)(2021年10期)2021-12-21
昆明医科大学学报(2021年10期)2021-12-02
中国卒中杂志(2021年7期)2021-11-29
今日农业(2021年2期)2021-11-27
新世纪智能(数学备考)(2020年10期)2021-01-04
今日农业(2020年23期)2020-12-31
