
苏门白酒草对草甘膦等除草剂的多抗性检测及防治药剂筛选
2022-08-10 01:55郭文磊于超杰张泰劼田兴山
农药学学报 2022年4期
郭文磊, 于超杰, 张 纯, 张泰劼, 田兴山
(广东省农业科学院 植物保护研究所/农业农村部华南果蔬绿色防控重点实验室/广东省植物保护新技术重点实验室,广州 510640)
苏门白酒草Conyza sumatrensis是菊科白酒草属草本植物,原产于南美洲,1850 年传入中国,2000 年以后在中国华南地区广泛分布[1]。苏门白酒草植株高大,繁殖能力强,且种子具有冠毛,易于随风传播,已被中国等多个国家列为最具威胁的入侵性植物之一[2]。在华南地区,苏门白酒草可在果园、茶园等造成危害,常形成单优势群落[3]。长期以来,使用草甘膦 (glyphosate)、百草枯 (paraquat) 等灭生性除草剂是防治苏门白酒草最方便、经济、快捷的方法。
草甘膦通过抑制5-烯醇丙酮酰-莽草酸-3-磷酸合成酶 (5-enolpyruvylshikimate-3-phosphatesynthase, EPSPS) 的活性而杀死杂草,EPSPS 活性被抑制后,莽草酸途径无法正常进行,进而导致苯丙氨酸、酪氨酸和色氨酸等芳香族氨基酸及大量次级代谢物质无法正常合成[4]。百草枯是联吡啶类除草剂,该类除草剂作用于光系统Ⅰ,充当电子传递受体,可迅速造成细胞死亡及植株枯死[5]。
草甘膦和百草枯均具有广泛的杀草谱,且除草活性优异,均为近几十年来全球使用最广泛的除草剂品种之一。由于长期重复使用相同的除草剂,使得杂草持续面临巨大的选择压力,最终导致抗药性种群的产生。最早被发现对草甘膦产生了抗药性的是澳大利亚的瑞士黑麦草种群[6]。1980 年,日本和台湾均发现了抗百草枯的白酒草属杂草,截至目前,全球分别已有55 种和32 种杂草对草甘膦和百草枯产生了抗性,其中多种杂草对两种除草剂具有多抗性[7]。在中国,目前已分别报道了小飞蓬Conyza canadensis和牛筋草Eleusine indica对草甘膦的抗性[8-9],而已报道对百草枯产生抗性的有反枝苋Amaranthus retroflexus、日本看麦娘Alopecurus japonicas、牛筋草、通泉草Mazus fauriei及硬草Sclerochloa dura等多种杂草[7,10-11]。因安全性风险问题,中国自2020 年9 月26 日起全面禁止了百草枯各种剂型产品的销售和使用,但与百草枯作用机理相同的另一联吡啶类除草剂敌草快 (diquat) 仍可继续使用。敌草快对阔叶杂草防效优异,具有见效快、传导性强、耐低温等优点,是百草枯被禁用之后的重要替代产品之一。据报道,对百草枯产生抗药性的杂草很可能对敌草快产生一定水平的交互抗性[12]。
本研究团队曾在广州市一处果园发现,喷施草甘膦后,果园内仍有部分苏门白酒草植株存活,疑似苏门白酒草已对草甘膦产生抗药性,另据果园管理者反映,该果园曾有十多年的百草枯使用历史,百草枯对苏门白酒草已基本失效。因此,本研究主要目的是明确该苏门白酒草种群对草甘膦和百草枯的抗性水平,以及对敌草快是否存在交互抗性,并就其抗草甘膦的机制进行初步探讨,以期筛选出合适的替代防治药剂。
1 材料与方法
1.1 供试材料
杂草种子:苏门白酒草Conyza sumatrensis种子于2017 年9 月在广东省不同地区采集。其中,疑似抗性种群 (GZ-R) 采集自广州市某荔枝园(N:23°23′30.36′′;E:113°25′17.66′′ ) 喷施草甘膦后仍存活的植株;敏感对照种群 (QY-S) 采集于广东省清远市一未使用过除草剂的荒地 (N:23°46′29.02′′;E:113°1′48.99′′ )。每个种群的种子均采自5 株以上植株,混合均匀后于室温干燥条件下保存。
除草剂:41% 草甘膦异丙胺盐 (glyphosate isopropylamine salt) 水剂 (AS),美国孟山都公司;20%百草枯 (paraquat) 可溶胶剂 (WSG),南京红太阳生物化学有限责任公司;20%敌草快 (diquat)AS,深圳诺普信农化股份有限公司;200 g/L 草铵膦 (glufosinate-ammonium) AS,浙江永农作物科学有限公司;70%苯嘧磺草胺 (saflufenacil) 水分散粒剂 (WG),巴斯夫欧洲公司;480 g/L 灭草松(bentazone) AS,江苏绿利来股份有限公司;20%氯氟吡氧乙酸 (fluroxypyr) 乳油 (EC),江苏中旗科技股份有限公司;56% 2 甲4 氯钠 (MCPA-sodium)可溶粉剂 (SP),重庆树荣作物科学有限公司。
主要仪器:ASS-4 型自动控制农药喷洒系统,国家农业信息化工程技术研究中心研制;T100 梯度PCR 仪和PowerPac Basic 电泳仪,美国Bio-Rad 公司;5824R 型台式冷冻离心机,德国Eppendorf 公司;勤翔1800 系列凝胶成像系统,上海勤翔科学仪器有限公司;JXESTPRP-24L 型全自动样品快速研磨仪,上海净信实业发展有限公司。
DP305 植物基因组DNA 提取试剂盒,天根生化科技 (北京) 有限公司。
1.2 试验方法
1.2.1 抗性水平测定 采用整株剂量反应法[13]。分别将GZ-R 种群和QY-S 种群的苏门白酒草种子均匀撒入盛有湿润壤土 (pH 6.5,有机质含量2.5%)的塑料盘 (23 cm × 23 cm × 3.5 cm) 中进行育苗,覆细土0.3 cm,将塑料盘放入玻璃温室 (自然光照,温度20~30 ℃,相对湿度70%~80%) 中进行培养。待幼苗长至3~4 真叶期时,将其移栽至盛有上述壤土、直径15 cm 的塑料花盆中,每盆6 株,置于玻璃温室中继续培养。
待植株长至5~6 真叶期时,采用ASS-4 型自动控制农药喷洒系统进行茎叶喷雾处理。配Teejet 9503EVS 除草剂专用扇形喷头,喷雾压力300 kPa,喷液量450 L/hm2。根据预试验结果,草甘膦施药剂量 (有效成分用量,下同) 分别设为:GZ-R种群,0、10.1、30.4、91.1、273.3、820.0 和2 460.0 g/hm2;QY-S 种群,0、3.4、10.1、30.4、91.1、273.3 和820.0 g/hm2。百草枯施药剂量:GZ-R 种群,0、19.8、59.3、177.8、533.3、1 600.0和4 800.0 g/hm2;QY-S 种群,0、2.2、6.6、19.8、59.3、177.8 和533.3 g/hm2。并采用相同方法验证GZ-R 种群对敌草快是否存在交互抗性。敌草快施药剂量为:GZ-R 种群,0、3.7、11.1、33.3、100、300 和900 g/hm2;QY-S 种群,0、1.2、3.7、11.1、33.3、100 和300 g/hm2。每处理3 次重复,试验共进行2 次。于施药后21 d,检查并记录每盆中苏门白酒草的存活情况:若植株保持绿色,且有新叶片长出,视为存活;若植株完全干枯或个别叶片虽保持绿色但无新叶片长出,则视为死亡。最后剪取地上部分植株,置于80 ℃干燥箱中烘干72 h,称量干重。
1.2.2 苏门白酒草EPSPS2 基因片段测序 据González-Torralva 等[14]的研究报道,EPSPS2 基因的Pro-106-Thr 突变 (以最早报道EPSPS基因抗性突变的牛筋草氨基酸序列为参照序列) 可导致苏门白酒草对草甘膦的抗性。为验证GZ-R 种群对草甘膦的抗性是否同样与EPSPS2 基因的突变有关,笔者对GZ-R 和QY-S 种群的EPSPS2 基因片段进行了扩增和测序。
于苏门白酒草长至4~5 叶期时,从GZ-R 和QY-S 种群中分别随机选取10 株进行基因组DNA 提取,提取方法按照DP305 试剂盒操作说明进行。参考González-Torralva 等[14]和Beres 等[15]的论文以及GenBank 数据库中3 种白酒草属植物小飞蓬 (AY545667.1)、苏门白酒草 (AY834205.1)及香丝草Conyzabonariensis(MG205557.1) 的EPSPS2 基因序列设计引物,正向引物序列为5’-GGACTACTGTTGTAGACAACTTA-3’,反向引物序列为5’-GAAACCCCAAACCGTTCC-3’,片段长度1 347 bp,包含了报道较多的EPSPS2 基因106 个氨基酸位点。PCR 扩增反应体系总体积50 μL,包括基因组DNA 2 μL,正向引物F 和反向引物R 各2 μL,2 ×EasyTaq®PCR SuperMix 25 μL,Nuclease-free Water 19 μL。PCR 反应条件:94 ℃预变性5 min;94 ℃变性30 s、58 ℃退火30 s、72 ℃延伸60 s,共进行35 个循环;最后于72 ℃终延伸7 min。PCR 反应结束后,每个样品取5 μL 于1.5% 的琼脂糖凝胶中进行电泳,将扩增成功的样品送至生工生物工程 (上海) 股份有限公司进行测序。每种群至少检测10 个样品。
1.2.3 防治药剂筛选 为筛选合适用于防控苏门白酒草抗性种群的除草剂,本研究测定了草铵膦、苯醚磺草胺、灭草松、氯氟吡氧乙酸和2 甲4 氯钠5 种除草剂对不同生长期苏门白酒草的防除效果。
试验采取分批培养、1 次施药的原则。选取苏门白酒草GZ-R 和QY-S 种群种子,分别在试验开始的第1 天、第10 天和第15 天,按照1.2.1 节中的方法进行育苗,每批幼苗于2~3 叶期进行移栽,待最后种植的1 批苏门白酒草植株长至4~5 叶期时统一进行茎叶喷雾处理,此时第1 批和第2 批植株分别处于10~12 叶期和6~7 叶期。每种除草剂各设1 个剂量处理,剂量设置参考中国农药信息网[16]中每种有效成分单剂登记剂量的上限,即草铵膦1 800.0 g/hm2、苯醚磺草胺78.8 g/hm2、灭草松1 440.0 g/hm2、氯氟吡氧乙酸210.0 g/hm2、2 甲4 氯钠1 008.0 g/hm2,以喷清水的处理作为空白对照。于施药后21 d,按照1.2.1 节中的方法检查并记录每盆中苏门白酒草的存活情况及地上部分干重。
1.2.4 数据处理 参考Yu等[17]的方法,利用SigmaPlot 12.5 软件对抗性水平测定试验数据进行回归分析。其中生长抑制中量 (GR50) 利用四参数双逻辑非线性回归模型 (1) 进行拟合处理得到。

式 (1) 中:y1为相对干重,即某一处理下苏门白酒草地上部分干重相对于空白对照的百分比;x为除草剂剂量;C表示剂量反应下限;D表示剂量反应上限;b为斜率。
致死中量 (LD50) 利用指数型衰减曲线模型 (2)进行拟合处理后得到。

式 (2) 中:y2为某一处理下苏门白酒草的存活率;x为除草剂剂量;y0为剂量反应下限,y0+a表示剂量反应上限;b为斜率。
抗性倍数(RI)计算公式见式 (3)。

式 (3) 中:GR50(LD50)R为抗性种群的GR50或LD50值;GR50(LD50)S为敏感种群的GR50或LD50值。
防治药剂筛选试验中,苏门白酒草的死亡率和干重抑制率计算公式见式 (4)。
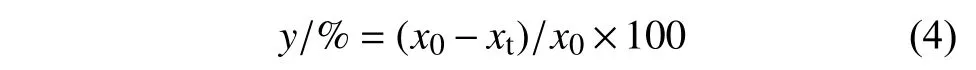
式 (4) 中:y表示杂草死亡率或干重抑制率;x0表示空白对照组杂草存活数或地上部分干重;xt表示药剂处理组杂草存活数或地上部分干重。
采用SPSS 21.0 软件,对每种药剂处理下不同时期苏门白酒草抗性和敏感种群的死亡率和干重抑制率分别进行单因素方差分析,采用LSD 法进行差异显著性分析。
2 结果与分析
2.1 苏门白酒草对草甘膦、百草枯及敌草快的抗性水平
就干重抑制率指标而言,苏门白酒草GZR 和QY-S 种群对供试不同除草剂的剂量反应曲线如图1 (a~c) 所示。其中,草甘膦对GZ-R 和QY-S种群的GR50值分别为143.1 和61.4 g/hm2,据此算得GZ-R 种群对草甘膦的抗性倍数为2.3;与对草甘膦的抗性相比,GZ-R 种群对百草枯的抗性水平更高 (图1 b),其抗性倍数高达35.1 (表1)。值得注意的是,两个苏门白酒草种群对敌草快的剂量反应也存在明显差异,GZ-R 种群对敌草快的抗性倍数达7.8 (表1),表明苏门白酒草GZ-R 种群在对百草枯产生高抗性的同时对敌草快已产生中等水平的交互抗性。
针对存活率指标,则苏门白酒草GZ-R 和QY-S种群对不同除草剂的剂量反应曲线如图1 (d~f)所示。与GR50值相比,不同除草剂对苏门白酒草GZ-R 和QY-S 种群的LD50值均明显提高,同时其抗性倍数也表现出一定变化。其中,GZ-R 种群对草甘膦的抗性倍数提升至7.2,对百草枯的抗性倍数提升至72.3,对敌草快的抗性倍数则与干重抑制率结果相比变化不大 (表1)。

图1 苏门白酒草不同种群对草甘膦、百草枯和敌草快的剂量反应曲线Fig. 1 The dose-response curve of different Conyza sumatrensis populations to glyphosate, paraquat and diquat

表1 草甘膦、百草枯和敌草快对苏门白酒草不同种群的生长抑制中量 (GR50) 和致死中量 (LD50)Table 1 The 50% growth reduction (GR50) and 50% mortality (LD50) of glyphosate, paraquat, and diquat to different C. sumatrensis populations
2.2 苏门白酒草EPSPS 基因片段测序结果
分别将测序得到的苏门白酒草GZ-R 和QYS 种群基因片段序列与已报道的苏门白酒草EPSPS2基因 (AY834205.1) 进行比对,发现其相似度达99% 以上,表明该片段即为苏门白酒草EPSPS2基因片段。进一步比对GZ-R 和QY-S 种群的EPSPS2 基因片段序列、测序峰图和翻译所得氨基酸序列发现,与敏感种群QY-S 相比,抗性种群GZ-R 在EPSPS2基因106 位点的碱基发生了CCA 至ACA 的替换,导致氨基酸由脯氨酸突变为苏氨酸 (Pro-106-Thr) (表2)。该突变已在抗草甘膦的苏门白酒草[14]、牛筋草[18]、瑞士黑麦草Lolium rigidum[19]及Digitaria insularis[20]等杂草中被报道,因此本研究中苏门白酒草GZ-R 种群对草甘膦的抗性很可能与该突变有关。

表2 苏门白酒草抗性与敏感种群EPSPS2 基因序列比对分析Table 2 Sequence alignments and deduced amino acids of the resistant and susceptible C. sumatrensis populations
2.3 多抗性苏门白酒草替代防除药剂筛选
苏门白酒草GZ-R 和QY-S 种群对5 种候选替代药剂的敏感性较为一致 (表3)。在同一除草剂处理下,相同叶龄GZ-R 和QY-S 种群植株的死亡率和干重抑制率均无显著性差异,说明已对草甘膦和百草枯产生抗性的GZ-R 种群可能尚未对灭草松、氯氟吡氧乙酸、2 甲4 氯钠、草铵膦和苯嘧磺草胺等除草剂产生交互抗性。

表3 候选5 种茎叶处理剂对不同生长期苏门白酒草的防除效果Table 3 The control efficacies of five candidate post-emergence herbicides on C. sumatrensis at different leaf stages
随着叶龄增大,灭草松、氯氟吡氧乙酸和2 甲4 氯钠处理组苏门白酒草的死亡率和干重抑制率均呈下降趋势:在3 种除草剂推荐剂量下,4~5 叶期时施药,GZ-R 和QY-S 种群的死亡率均为100%,干重抑制率在90%以上;于10~12 叶期时施药,GZ-R 和QY-S 种群死亡率和干重抑制率分别下降至44.4%~72.2%和48.1%~74.1%。与上述3 种除草剂相比,草铵膦和苯嘧磺草胺对苏门白酒草两个种群均表现出更好的防除效果:采用推荐剂量的草铵膦和苯嘧磺草胺于不同叶龄期施药,GZ-R 和QY-S 种群死亡率均为100%,干重抑制率在87.9%~93.7%之间。
3 讨论与结论
随着不同作用类别除草剂的连续使用,杂草对两种甚至多种不同类别除草剂的多抗性现象愈发普遍,在澳大利亚甚至已发现瑞士黑麦草种群对6 种不同作用类别的除草剂产生了抗性[7]。我国目前已报道的主要为小麦田或水稻田杂草对ALS 和ACCase 抑制剂类除草剂的多抗性,如小麦田看麦娘Alopecurus aequalis[21]、日本看麦娘[22]、菵草Beckmannia syzigachne[23]对甲基二磺隆 (mesosulfuron-methyl) 和精噁唑禾草灵(fenoxaprop-P-ethyl) 的多抗性,水稻田稗Echinochloacrus-galli对氰氟草酯 (cyhalofopbutyl)、五氟磺草胺 (penoxsulam) 和二氯喹啉酸(quinclorac) 的多抗性[24-25],Deng 等[26]则发现,江苏省稻田牛筋草种群对草甘膦和氰氟草酯产生了多抗性。与ALS 和ACCase 抑制剂类除草剂相比,我国目前针对草甘膦和百草枯等灭生性除草剂抗性的研究相对较少,主要集中在牛筋草对草甘膦[27-28]或百草枯[11,29]的抗性以及小飞蓬对草甘膦[8,30]的抗性方面,而美国和澳大利亚已分别报道了17 种和20 种草甘膦抗性杂草[7]。这可能与不同国家除草剂的使用频率和使用年限等客观因素有关,但另一方面也提醒研究者需加强对草甘膦等灭生性除草剂的抗性监测。本研究中,苏门白酒草GZ-R 种群对百草枯已产生高水平抗性,对草甘膦也产生了中等水平抗性,这是在我国首次报道阔叶杂草对草甘膦和百草枯产生了多抗性。
本研究中发现,苏门白酒草GZ-R 种群在对百草枯产生高抗性的同时还对敌草快产生了中等水平的交互抗性。Vaughn 等[31]曾报道,香丝草对百草枯的抗性倍数已达100 以上,但对敌草快的抗性倍数小于10。Koschnick 等[32]和Weaver 等[33]则报道,少根紫萍Landoltia punctate、小飞蓬等杂草对百草枯和敌草快均具有高抗性。因此,虽然目前GZ-R 种群对敌草快的抗性水平仍较低,但不排除有继续升高的风险,故不建议使用敌草快防除已对百草枯产生抗性的杂草。
草甘膦和百草枯对苏门白酒草两个种群的LD50值均比GR50值明显升高,其中抗性种群GZ-R 升高的幅度更大,草甘膦和百草枯对GZ-R种群的LD50值较其GR50值分别提升了8 倍和4 倍,因此采用LD50值计算出的抗性倍数也分别比采用GR50值计算的抗性倍数提高了2 倍和1 倍(表1)。Yu 等[17]的研究表明,野胡萝卜Raphanus raphanistrum抗性种群对氯磺隆 (chlorsulfuron) 的LD50值 (82.0 g/hm2) 是其GR50值 (4.3 g/hm2) 的19 倍,以GR50值计算出的抗性倍数为6.9,而以LD50计算出的抗性倍数在131 以上。本研究中,草甘膦和百草枯对苏门白酒草GZ-R 种群的LD50值分别为 (1 371.2 ± 231.1) 和 (2 132.1 ± 50.4) g/hm2(表1),均达到或超过了其推荐剂量,据此可以推断其LD90值必然远超推荐剂量。事实上,杂草在一定剂量范围内虽然生长受到抑制,生物量明显降低,但假如其并未死亡,仍可产生大量可育的种子。因此,在抗性水平测定试验中,同时测定杂草在不同剂量下的死亡率可能比单独测定干重或鲜重抑制率更具有实际意义。
本研究发现,苏门白酒草GZ-R 种群EPSPS2基因发生了Pro-106-Thr 突变,该突变之前在苏门白酒草[14]、牛筋草[18]、瑞士黑麦草[19]等杂草中已有报道,所引起的抗性倍数一般在2~7 之间,因此认为该突变很可能是导致GZ-R 种群对草甘膦产生中等水平抗性的重要原因。值得注意的是,杂草对草甘膦产生抗性后,若提高草甘膦剂量继续使用,可能会导致其抗性程度进一步发展,如由Pro-106-Thr 突变和转运减少等非靶标抗性机制共同介导的瑞士黑麦草对草甘膦的抗性倍数在10~12 之间[19],而EPSPS基因102 位和106 位双突变的牛筋草对草甘膦的抗性倍数高达180[34]。
一般而言,随着叶龄增大和植株生长,杂草对除草剂的耐药性会明显增强。本研究表明,在推荐剂量下,灭草松、氯氟吡氧乙酸和2 甲4 氯钠对4~5 叶期苏门白酒草具有较好的防效,但对6~7 叶期和10~12 叶期苏门白酒草的防效明显下降。Weaver 等[33]的研究表明,乙氧氟草醚(oxyfluorfen) 对生长56 d 小飞蓬的ED50值 (相当于本研究中的GR50值) 是对35 d 小飞蓬ED50值的6.9 倍,而2 倍推荐剂量的乙氧氟草醚对生长70 d 小飞蓬并无明显作用。因此,在使用除草剂防除果园苏门白酒草时,应尽量选择在其生育期早期以及天气等外界条件合适的情况下施药,以实现在减少除草剂用量的同时保证良好的防除效果。本研究中,虽然草铵膦和苯嘧磺草胺对各叶龄的抗性苏门白酒草均具有较好的防效,但在生产中防除抗性苏门白酒草时,仍建议选择不同作用机制的除草剂混剂或混配后使用[35],尽量避免同一年度内多次使用相同作用机制的药剂。
本研究仅进行了几种药剂对不同叶龄苏门白酒草的室内生物测定试验,后续还需进一步结合各作物田/果园已登记药剂情况开展田间试验进行验证。另外,果园生草不仅可控制田间杂草,还可以调节果园生态环境[36-37],因此可探索结合果园生草等生物防治方法控制苏门白酒草,以尽量减少除草剂的使用。
猜你喜欢
当代水产(2022年7期)2022-09-20
方圆(2021年21期)2021-11-20
农村百事通(2020年23期)2020-12-28
植物保护(2019年2期)2019-07-23
小天使·二年级语数英综合(2019年2期)2019-01-12
今日农药(2017年9期)2017-09-30
儿童故事画报(2016年1期)2016-03-09
山东农药信息(2013年12期)2014-01-15
现代农业科技(2009年19期)2009-03-20
