
1株鸡滑液囊支原体的分离鉴定与全基因组分析
2022-08-26 08:50许李锋贾晨宇陈福再方梦园刘孝丹陈吉龙李训良
畜牧兽医学报 2022年8期
许李锋,贾晨宇,陈福再,方梦园,刘孝丹,陈吉龙,李训良
(福建农林大学动物科学学院(蜂学学院),福建省畜禽病原感染与免疫学重点实验室,福建省-尼泊尔畜禽重大疫病防控联合实验室,福州 350002)
鸡滑液囊支原体(,MS)属于柔膜体纲支原体属,直径0.2~0.4 μm,以芽生方式或二分裂法繁殖,经姬姆萨染色后可以更好地观察其多形性,在电子显微镜下观察呈圆形或梨形等;MS只有一个血清型,但不同基因型的菌株致病力却存在差异,引起的症状也有所差异,部分菌株可能不产生或很少产生临床症状。MS基因组的特点是G+C含量低,由于其有限的遗传信息,支原体表达少量的细胞蛋白并且缺乏许多酶活性和代谢途径。因此,它们的营养需求很复杂,许多营养素都依赖于宿主。MS主要引起鸡等禽类的上呼吸道疾病、气囊炎、滑膜炎、腱鞘炎和滑囊炎。MS感染与蛋壳顶端异常(EAA)有关,使蛋的品质下降和质量减轻,导致出现异常卵增多,蛋壳易脆等问题。MS感染在中国非常普遍,一年四季均可发生。MS既可以水平传播也可以垂直传播,对任何年龄的鸡都可以造成感染。由于其感染隐蔽,不易根除,造成家禽生长迟缓、饲料利用率下降等,是目前导致养禽业经济损失的重要因素。
为了减少因野生型MS菌株感染引起的疾病和经济损失,可以使用3种不同的措施用以预防和控制:鸡场净化,抗生素治疗和预防接种。鸡场净化主要是检测鸡群MS的感染,通过淘汰患病鸡群,培育无MS感染的种鸡群。但由于中国禽业养殖场的规模和管理级别的不同,加之鸡场净化的成本高昂,很难建立和维护无MS感染的种鸡群,因此这种方法采用较少。而抗生素的频繁使用,不仅容易导致菌株产生耐药性,而且存在药物残留影响食品安全的风险。目前,市场上存在两种预防MS的减毒活疫苗,分别是对温度敏感的弱毒MS-H疫苗株和烟酰胺腺嘌呤二核苷酸(NAD)非依赖性MS1疫苗株。MS-H疫苗株目前已在全世界广泛使用,有效地控制了MS感染,降低了鸡群死亡率,延长了产蛋期,提高了鸡群产蛋量和饲料转化率;MS1疫苗由Nobilis公司生产,目前,在该公司已经找不到该疫苗的相关信息,该疫苗也未在我国农业农村部注册。MS弱毒疫苗虽然可以减轻MS感染引起的临床症状,提高鸡的生产性能,但其也存在一些缺点,它只能应用于无MS感染的鸡场,这在很大程度上限制了其应用。此外,MS弱毒疫苗具有水平传播的风险,并可能在同一鸡场的圈舍中相互传播,导致未接种疫苗的禽感染MS。基因工程疫苗因其安全性高、易量产、生产成本低等优点,已成为未来MS疫苗研发方向,因此筛选具有良好免疫效果的抗原就成为了研制基因工程疫苗的关键因素。
目前,MS基因工程亚单位疫苗的研究还处于起步阶段,因此需要依托大量基因组数据为疫苗免疫保护性抗原的筛选提供信息,然而对MS基因组测序分析的文章仍然较为缺乏。因此本文对分离获得的福建鸡滑液囊支原体菌株MS-FJ01进行全基因组测序分析,利用COG、GO 和KEGG等数据库进行了基因功能注释和富集分析;与GenBank中公布的其他MS分离株的16S rRNA等基因进行进化树分析比对;对该菌株的耐药基因、毒力基因等关键基因进行分析。研究结果为该病原菌致病性相关分子机制的深入解析以及基因工程亚单位疫苗的研发提供科学依据。
1 材料与方法
1.1 主要试验材料
病料来源于福州某鸡场疑似感染MS的病鸡肿胀跗关节腔内容物;改良Frey氏液体培养基和固体培养基购自青岛海博生物公司;0.45 μm过滤器购自Biosharp 公司;SPF雏鸡购自广东新兴大华农SPF实验动物中心;核酸 Marker、2×PCR Master Mix购自Biomed公司;庆大霉素、链霉素、林可霉素、泰乐菌素、替米考星、恩诺沙星、环丙沙星、四环素购自芊守生物科技有限公司。
1.2 鸡滑液囊支原体的分离
采集疑似感染MS病鸡的关节液及内容物,接种于改良Frey氏液体培养基中,置37 ℃恒温振荡培养48 h,待菌液变为黄色时,用0.45 μm过滤器过滤后按1∶10的比例重新接种到新的液体培养基,收取第2代菌液用于病原菌鉴定。
1.3 鸡滑液囊支原体的鉴定
1.3.1 菌落形态观察 取分离的第2代菌液接种于改良Frey氏固体培养基上,并置于体积分数为5%的CO培养箱中,37 ℃条件下培养7 d,于显微镜下观察菌落形态。
1.3.2 菌体L型鉴定 菌体L型鉴定方法参照丁美娟2014年《鸡滑液囊支原体的分离和鉴定研究》。
1.3.3 活菌计数 取15支装有0.9 mL改良Frey氏液体培养基的EP管,在第1管中加入0. 1 mL第2代菌液,充分混匀后从第1管吸出0.1 mL液体加入第2管,再混匀,吸出0.1 mL加入第3管,依次类推,直到第15管。另取2支含1 mL改良Frey氏液体培养基的EP管作为对照,置于37 ℃培养箱中静置培养,做3组重复。2周后观察培养基颜色变化,并判定结果。
1.3.4 人工感染鉴定 取分离的第2代菌液0.5 mL,滴鼻点眼0.4 mL,脚垫内注射0.1 mL接种于5日龄SPF雏鸡,持续观察鸡的生长及发病情况。
1.3.5 分子生物学鉴定 根据GenBank中登录的MS基因全序列,使用Primer 5软件设计合成一对特异性引物,分别为MS-JD506-1:5′-CTTCTATGCTTAAACTTTCC-3′,MS-JD506-2:5′-TAAAGATATTACAACGACAT-3′,预期扩增的目的片段大小为506 bp。并根据文献[15]合成目的片段大小为208 bp的一对引物,MS-208-F:5′-GAAGCAAAATAGTGATATCA-3′,MS-208-R:5′-GTC-GTCTCCGAAGTTAACAA-3′。用MS特异性引物PCR扩增此分离株。采用20 μL PCR体系:2×Taq PCR Master Mix 10 μL,无菌水8 μL,上下游引物各0.5 μL,菌液1 μL。反应条件:95 ℃ 5 min;95 ℃ 30 s,56.5 ℃ 30 s,72 ℃ 30 s,35个循环;72 ℃ 5 min;4 ℃保存。PCR产物经1%琼脂糖凝胶电泳检测并拍照。
1.4 最小抑菌浓度(MIC)测定
参考Hannan推荐的微量稀释法测定分离株MIC。
1.5 MS-FJ01总基因组的提取
取MS-FJ01菌液5 mL接种于含45 mL改良Frey氏液体培养基的离心管中,在37 ℃条件下培养至对数期后,在15 000 ×4 ℃条件下离心30 min 收集MS-FJ01沉淀物,超纯水清洗1遍后,再次离心收集MS-FJ01沉淀物。参照OMEGA细菌基因组抽提试剂盒步骤提取MS-FJ01的总DNA,利用微量紫外分光光度计测定DNA浓度。
1.6 MS-FJ01测序、组装
使用琼脂糖凝胶电泳检测基因组DNA的完整性,将检验合格的基因组DNA随机打断、消化、修复,之后使用0.45X的PB磁珠进行二次筛选纯化,获得测序文库。对文库质量进行检测,结果达到要求后,使用PacBio Sequel Ⅱ 进行文库的测序,然后使用SMRT LINK 8.0软件进行数据处理,并使用Illumina二代测序进行补充和准确性复核。
采用Microbial Assembly(smrtlink8)、HGAP4软件(smrtlink8)和Canu(v1.6)软件对纯三代数据结果进行组装,通过 Glimmer(v3.02)软件对组装结果进行基因模型预测。
1.7 基因功能的注释
利用tRNAscan-SE(v2.0),RNAmmer(v1.2)进行细菌基因组的tRNA、rRNA的预测;使用trf409.legacylinux64软件预测基因组中串联重复序列(TRF);利用MinCED(v0.3.2)对细菌基因组进行成簇规律间隔的短回文重复序列(CRISPR)结构预测;利用软件IslandPath-DIOMB预测基因组的基因岛;采用PhiSpy(v2.3)软件预测基因组中的前噬菌体结构;预测到的编码序列利用NR(ftp://ftp.ncbi.nih.gov/blast/db)、SwissProt(http://www.ebi.ac.uk/swissprot/)、COG(ftp://ftp.ncbi.nih.gov/pub/COG)、KEGG(http://www.kegg.jp/orhttp://www.genome.jp/kegg/)、GO(http://www.geneontology.org)、CARD(http://arpcard.mcmaster.ca)、CAZy(http://www.cazy.org/)、PHI(http://www.phi-base.org/index.jsp)、VFDB(http://www.mgc.ac.cn/VFs/)、TCDB(http://www.tcdb.org/)、RMS(http://rebase.neb.com/rebase/rebase)进行比对,注释功能基因。使用软件EffectiveT3(v2.0.2)进行T3SS效应蛋白的注释。
1.8 比较基因组学
在全基因组测序的基础上,将MS-FJ01的Eno和16S rRNA基因分别登录NCBI,通过BLAST与GenBank中的已知序列比对,选取HN01、G3、A4、WVU1853等十几株相似性高的MS相应基因序列,利用MEGA7软件构建系统进化树。将MS-FJ01的基因信息与NCBI上已公布的15个完整菌株进行比对。使用MUMmer软件对MS-FJ01和中国流行株HN01和5-9基因组进行共线性比较。
2 结 果
2.1 鸡滑液囊支原体的分离鉴定
取过滤后的菌种接种于改良Frey氏固体培养基上,培养7 d后在显微镜下观察可发现细小、光滑、致密的小菌落,表现为“煎蛋样”,半凹陷于培养基内(图1)。菌体L型鉴定的结果显示为阴性;活菌计数不低于10CCU·mL。

图1 鸡滑液囊支原体菌落图Fig.1 Colony diagram of Mycoplasma synoviae in chicken
取过滤后的菌种攻毒SPF雏鸡,25 d后可发现感染鸡出现精神沉郁、跛行、瘫痪、膝关节发炎肿大、关节和气囊内有干酪样沉积物等明显的病变(图2)。
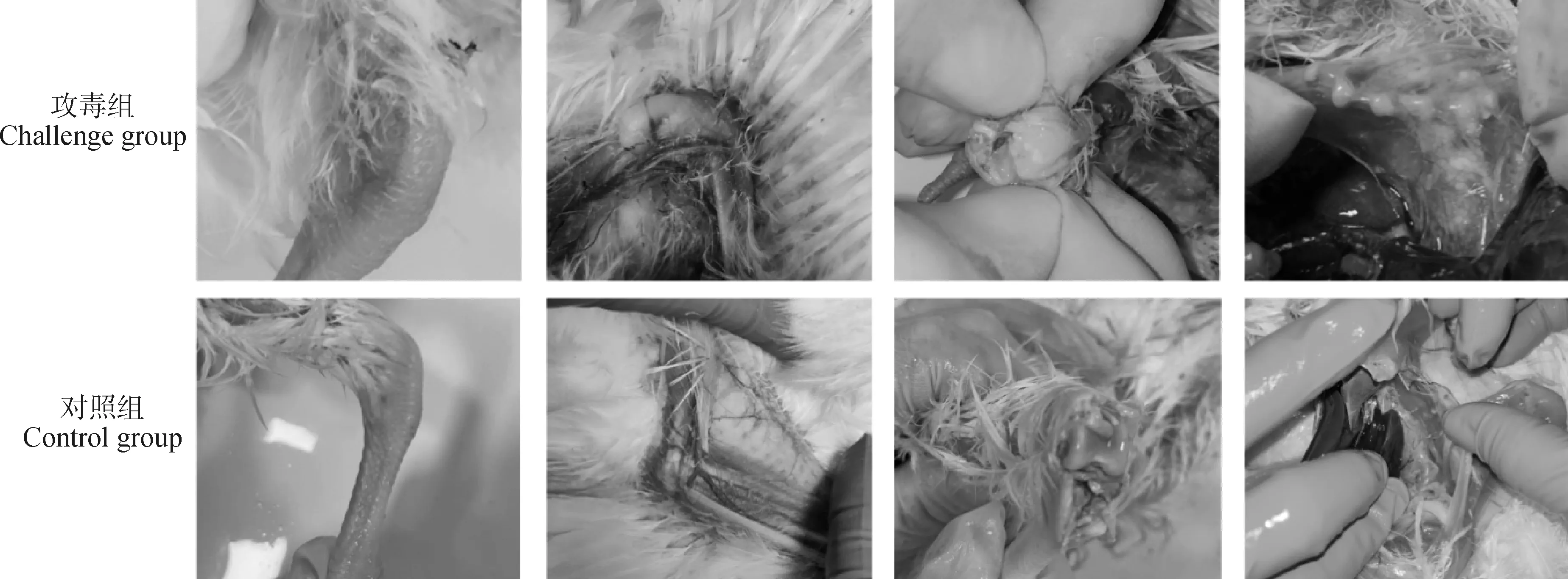
图2 鸡滑液囊支原体攻毒至SPF鸡后出现的病变Fig.2 Pathological changes of Mycoplasma synoviae in SPF Chickens
用MS特异性引物PCR扩增此分离株,能扩增出约506和208 bp的特异性片段(图3)。通过以上各项鉴定结果表明,所分离的菌株符合MS特性。收集60~100 mL培养至对数生长期的菌液,离心收集菌体沉淀物,并进行冻干保存,送往中国典型培养物保藏中心(CCTCC)保藏。所述的鸡滑液囊支原体其分类命名为MS-FJ01,保藏单位为中国典型培养物保藏中心(CCTCC),保藏编号为CCTCC No.: M 2021210。

M.DNA相对分子质量标准;1.本研究设计引物的鉴定结果;2.引用参考文献[15]的引物鉴定结果M.DNA Marker; 1. The identification results of primers designed in this study; 2. The identification result of primers quoted from references [15]图3 鸡滑液囊支原体核酸电泳鉴定图Fig.3 Electrophoretic identification of Mycoplasma synoviae in chicken
2.2 MS-FJ01对抗菌药物敏感性试验
通过微量肉汤稀释法,测得泰乐菌素、替米考星素等8种抗菌药物对MS-FJ01的MIC。试验结果表明,MS-FJ01对氟喹诺酮类耐药,而四环素和泰乐菌素对其则有较好的抑制作用。
2.3 MS-FJ01的基本信息
MS-FJ01总DNA提取浓度≥50 ng·μL,样品纯度:OD/OD=1.85,OD/OD=2.0。使用三代测序得到MS-FJ01的基因组全长为795 381 bp,GC含量为28.39%,编码基因704个,34个 tRNA以及7个rRNA。MS-FJ01基因组的圈图如图4所示。
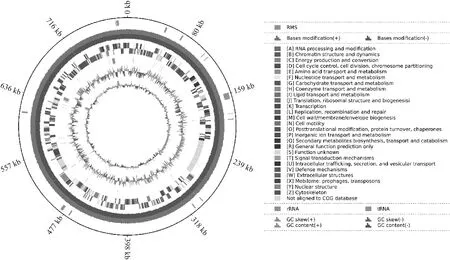
圈图从外到内第1圈为基因组大小;第2圈散在分布的点为限制性修饰酶分布情况;第3圈和第4圈分别为正链和负链的碱基修饰;第5圈和第6圈分别为正链和负链上的CDS;第7圈为tRNA和rRNA的分布;第8圈为GC-skew值,一般前导链GC-skew>0,后滞链GC-skew<0,也可以辅助判断复制起点(累计偏移最小值)和终点(累计偏移最大值),尤其对环状基因组最为重要;最内圈为GC含量,向外部分表示该区域的GC含量高于基因组的平均水平,向内部分表示该区域的GC含量低于基因组的平均水平The first circle of the circle from outside to inside is genome size; The scattered points in the second circle are the distribution of restrictive modification enzymes. The third and fourth circles are the base modification of plus and minus chains respectively. The fifth and sixth circles are CDS on the positive and negative chains respectively; The seventh circle is the distribution of tRNA and rRNA. The eighth circle is GC-skew value, generally leading chain GC-skew>0, lag chain GC-skew<0, which can also assist in determining the starting point (minimum cumulative offset) and end point (maximum cumulative offset) of replication, especially for circular genomes. The innermost circle is GC content, with the outward part indicating that GC content in this region is higher than the average level of the genome, and the inward part indicating that GC content in this region is lower than the average level of the genome图4 MS-FJ01基因组圈图Fig.4 Genome circle of MS-FJ01
本研究预测TRF有79个,TRF的长度变化范围从1~500 bp不等,表现出种属组成特异性,可作为物种的遗传性状、进化关系的研究。预测CRISPR序列2个,CRISPR序列存在于许多细菌和古细菌中,其与CRISPR相关基因构成CRISPR-Cas系统,目前的研究证明,该系统很可能是原核细菌抵御外来入侵者重要的防御系统。
基因岛与多种生物功能相关,因此一直以来都是研究的热点,本研究预测到3个基因岛。前噬菌体序列的存在可能会允许一些细菌获取抗生素抗性,增强对环境的适应性,提高黏附力或使细菌成为致病菌,本研究预测出1个前噬菌体结构。MS-FJ01全基因组数据已收录GenBank,收录号为CP079705。
2.4 基因功能注释分析
为进一步解析MS-FJ01基因组的功能,将其基因组序列在COG、GO和KEGG共3个数据库中进行注释。
2.4.1 COG数据库注释 COG数据库按照基因功能分为25大类,每一个COG类别均由直系同源序列构成。因此,通过序列相似性比对,可以将某个蛋白序列归类于某一COG簇中,从而可以推测该序列的功能。作者将预测基因的蛋白序列比对到COG库。如图5所示,在MS-FJ01基因组中与翻译、核糖体结构和生物起源相关的基因最多,有133个基因;与复制、重组和修复相关的基因次之,有50个基因;与细胞运动和次生代谢产物生物合成、转运和分解代谢相关的基因最少,各仅有1个基因;但仍有7个功能未知的基因。

C. 能量的产生和转化;D. 细胞周期调控、细胞分裂和染色体分区;E. 氨基酸的转运和代谢;F. 核苷酸的转运和代谢;G. 碳水化合物的运输和代谢;H. 辅酶的运输和代谢;I. 脂质运输和代谢;J. 翻译、核糖体结构和生物发生;K. 转录;L. 复制、重组和修复;M. 细胞壁/膜/包膜生物发生;N. 细胞运动;O. 翻译后修饰、蛋白质更新和伴侣蛋白;P. 无机离子的运输与代谢;Q. 次生代谢产物生物合成、转运和分解代谢;R. 一般功能预测;S. 未知功能;T. 信号转导机制;U. 细胞内运输、分泌和囊泡运输;V. 防御机制;X. 噬菌体和转座子C. Energy production and conversion; D. Cell cycle control. cell division, chromosome partitioning; E. Amino acid transport and metabolism; F. Nucleotide transport and metabolism; G. Carbohydrate transport and metabolism; H. Coenzyme transport and metabolism; I. Lipid transport and metabolism; J. Translation, ribosomal structure and biogenesis; K. Transcription; L. Replication, recombination and repair; M. Cell wall/membrane/envelope biogenesis; N. Cell motility; O. Posttranslational modification, protein turnover, chaperones; P. Inorganic ion transport and metabolism; Q. Secondary metabolites biosynthesis, transport and catabolism; R. General function prediction only; S. Function unknown; T. Signal transduction mechanisms; U. Intracellular trafficking secretion, and vesicular transport; V. Defense mechanisms; X. Mobilome: prophages, transposons图5 MS-FJ01基因组COG注释统计Fig.5 COG annotation statistics of MS-FJ01 genome
2.4.2 GO数据库注释 GO数据库将蛋白序列分为3大类:生物过程、分子功能和细胞组分,分别用来描述基因编码的产物所参与的生物过程、所具有的分子功能及所处的细胞环境。如图6所示,MS-FJ01基因组中,在生物过程大类中,与代谢过程(基因数 249)和细胞过程(基因数 237)相关的基因最多。在细胞组分大类中,被注释最多的是细胞(基因数 170)、细胞部件(基因数 170)、膜(基因数 95)以及膜部件(基因数 91)。在分子功能途径大类中,与催化活性(基因数 219)和结合(基因数 228)相关的基因最多。
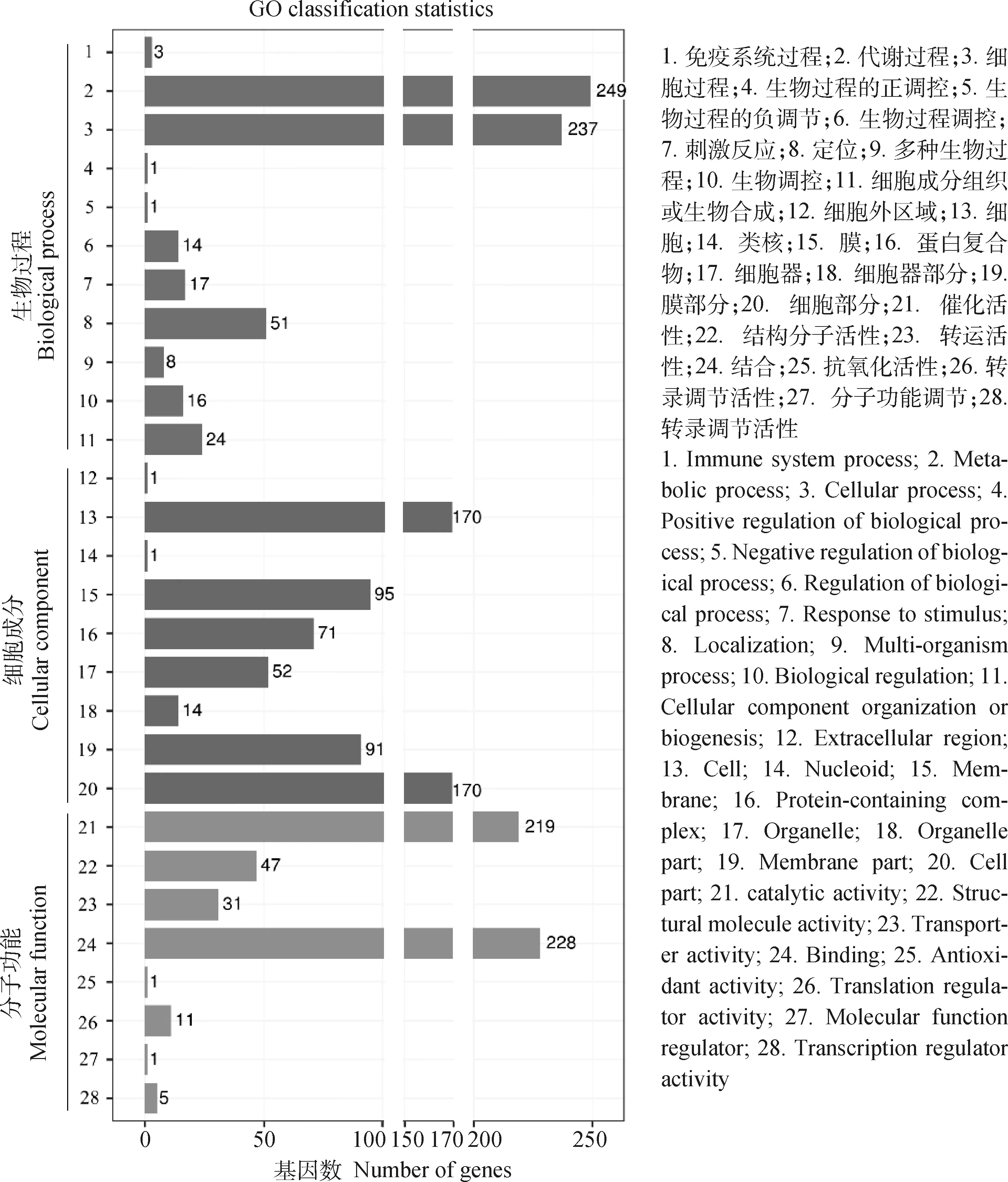
图6 MS-FJ01基因组GO注释统计Fig.6 GO annotation statistics of MS-FJ01 genome
2.4.3 KEGG数据库注释 KEGG是系统分析基因产物和化合物在细胞中的代谢途径以及这些基因产物功能的数据库;它整合了基因组信息、化合物和小分子信息以及生化反应系统等方面数据,主要包括代谢通路、药物、疾病、功能模型、基因序列及基因组等信息。如图7所示,MS-FJ01基因主要分为细胞过程、环境信息加工、遗传信息加工、人类疾病、代谢及机体系统6大分类。其中,在新陈代谢和遗传信息处理中占的基因比例较高。在这些类别中,基因富集最多的类别是代谢(基因数占比34.8%),主要包括碳水化合物代谢、能量代谢、辅助因子和维生素的代谢以及核苷酸代谢等。遗传信息加工富集的基因次之(基因数占比29.1%),其中翻译(基因数 78)富集的基因最多。六大类中富集最少的是机体系统,仅占全部富集基因数的1.6%。
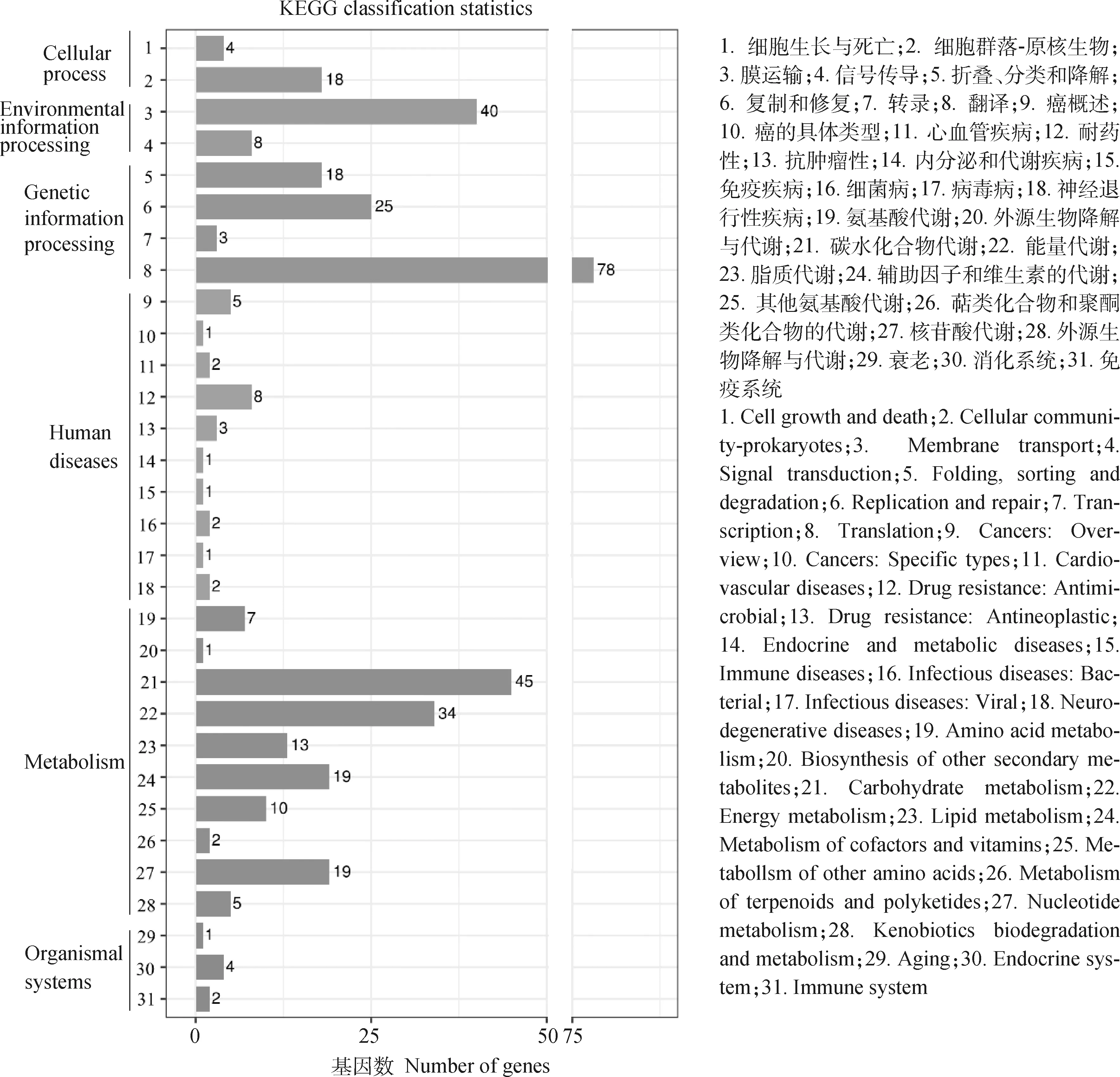
图7 MS-FJ01基因组KEGG注释统计Fig.7 KEGG annotation statistics of MS-FJ01 genome
2.5 特定功能注释
将只整合某一类功能相关的数据库和注释方法划分到特定功能注释模块,该模块包含耐药基因(CARD)、碳水化合物相关酶(CAZy)、病原与宿主互作(PHI)、致病菌毒力因子(VFDB)、膜转运蛋白分类(TCDB)、限制性修饰系统(RMS)、Ⅲ型分泌系统效应蛋白(T3SS)。
2.5.1 MS-FJ01基因组CARD数据库注释 CARD(The Comprehensive Antibiotic Resistance Database)是目前使用最为广泛的细菌耐药基因数据库。因此使用CARD数据库做耐药基因注释,本研究在CARD库注释到了40个细菌耐药基因。其中对四环素具有抗性的基因最多(基因数7),此外,基因具有多抗生素耐药性。详见表1。

2.5.2 MS-FJ01基因组CAZy数据库注释 CAZy全称为Carbohydrate-Active enZYmes Database,碳水化合物酶相关的专业数据库,内容包括能催化碳水化合物降解、修饰、以及生物合成的相关酶系家族。其包含6个主要分类:辅助氧化还原酶(auxiliary activities,AAs)、糖苷水解酶(glycoside hydrolases, GHs)、糖基转移酶(glycosyl transferases,GTs)、多糖裂解酶(polysaccharide lyases, PLs)和糖类酯解酶(carbohydrate esterases, CEs)。本研究中,在CAZy库共注释到4个基因,其中,3个为糖苷水解酶,1个为糖类酯解酶。
2.5.3 MS-FJ01基因组PHI数据库注释 PHI全称为Pathogen Host Interactions Database,病原与宿主互作数据库, PHI是目前现医疗、农业真菌和卵菌候选靶向位点的重要在线资源。在 PHI 数据库中注释到了136个与宿主互作的基因,97个基因在突变后会导致病原菌致病性的减弱或丧失,15个基因在突变后会使得病原菌的致病性增强。
2.5.4 MS-FJ01基因组VFDB数据库注释 VFDB数据库全称为Virulence Factors of Pathogenic Bacteria,用于专门研究致病细菌、衣原体和支原体致病因子的数据库。在本研究MS-FJ01共有47个基因与细菌毒力相关,其中,序列相似度在40%以上的基因只有3个,分别为胆碱激酶、脂酸蛋白连接酶和ABC(ATP结合盒)转运体CylA,其他基因序列相似度均低于40%。另外,作者还挖掘到DnaK、PDHA、PDHB、Eno、GapA、PK、FBA、NOX等多个与细菌黏附相关的基因。
2.5.5 MS-FJ01基因组TCDB数据库注释 TCDB是对膜转运蛋白(Membrane Transport Protein)进行分类的一个数据库,它制定了一套转运蛋白分类系统(Transporter Classification), 简称TC System,TC系统除了对膜转运蛋白进行分类,同时还提供了其功能和进化信息。本研究共注释到106个与膜转运蛋白相关的基因,其中,与初级活性转运体相关的基因最多(基因数 64)。
2.5.6 MS-FJ01基因组RMS数据库注释 RMS主要由限制性内切酶(restriction enzymes)及甲基化转移酶(methyltransferase)两部分所构成。限制修饰系统按照组成功能上的差异主要分为基本的四种类型,分别是Type I systems、Type II systems、Type III systems、Type IV systems。该系统对自身基因组某些Motif的修饰可能会对其基因表达及生长代谢调控有非常重要的作用。REBASE包含大量限制修饰系统的分类、识别修饰、识别切割位点等信息。在本研究,中Ⅲ型甲基化转移酶(基因数 8)和Ⅲ型限制性内切酶(基因数 7)注释的基因数最多。
2.6 比较基因组学分析
2.6.1 进化树分析 MS的烯醇化酶(Eno)是一种重要的黏附相关因子,对宿主细胞的黏附、定植和侵袭有着重要作用。因此作者使用MS的和16S rRNA基因的DNA序列进行系统发育树分析。参考菌株与MS-FJ01的16S rRNA序列构建的进化树拓扑结构显示,MS-FJ01与2020年分离的中国宁夏菌株5-9亲缘关系最近,而与2018年分离的河南菌株HN01、韩国菌株G3、A4和模式MS菌株WVU1853等亲缘关系则较远,在基于的进化树拓扑结构中显示出的结果也表明MS-FJ01与MS 5-9菌株亲缘关系最近(图8)。

A. MS-FJ01的16S rRNA系统进化树;B. MS-FJ01的Eno系统进化树A. 16S rRNA phylogenetic tree of MS-FJ01;B. Eno phylogenetic tree of MS-FJ01图8 MS-FJ01的系统进化树Fig.8 Phylogenetic tree of MS-FJ01
2.6.2 MS-FJ01与MS菌株基因组基本信息对比 本研究将MS-FJ01相关基因信息与GenBank中已公布的15个MS菌株的完整基因组序列进行比较分析,该菌株基因信息与已公布的MS参考菌株基本相似,只有编码基因的数目少于其他菌株。
2.6.3 基因组共线性分析 MS-FJ01与中国流行株HN01和5-9基因组进行比对,确认基因组序列之间的共线性关系及结构变异情况,由图9可知MS-FJ01与5-9基因组共线性程度明显高于HN01基因组;相比于5-9基因组,HN01存在较多基因倒置、缺失和分布不同。

A.MS-FJ01和HN01基因组共线性分析;B. MS-FJ01和5-9基因组共线性分析A.Genomic collinearity analysis of MS-FJ01 and HN01;B. Genomic collinearity analysis of MS-FJ01 and 5-9图9 MS-FJ01共线性分析Fig.9 MS-FJ01 collinearity analysis
3 讨 论
本研究通过三代联合二代测序技术,对MS-FJ01进行了全基因组精细测序,并对该基因组的基本特征进行了分析。MS目前已有15个完整基因组被公布,但相同物种的不同菌株在基因和蛋白结构上仍存在差异。本研究将MS-FJ01基因组预测基因的蛋白序列比对到NR数据库发现所有蛋白序列匹配的同源蛋白相似度最高分别是支原体属(53.9%)和鸡滑液囊支原体 (34.54%)。之后又将预测基因的蛋白序列比对到SwissProt数据库进行验证,发现相似度最高的物种仍然是鸡滑液囊支原体(29.6%),剩余的相似性大部分都属于支原体属,包括肺炎支原体、生殖支原体等。
作者在MS-FJ01基因组的编码基因中预测到长度为1 010 bp的烟酸磷酸核糖基转移酶和长度为674 bp的3-磷酸甘油酰基转移酶,Yongjun等在韩国分离株的基因组中发现这两种编码序列仅存在于NAD非依赖性MS的基因组中。Yagihashi等在日本发现的菌株也具有NAD非依赖性。MS-FJ01是否具有NAD非依赖性还有待后续试验进行验证,如果MS-FJ01等中国菌株具有NAD非依赖性,那么NAD非依赖性则可能是亚洲MS菌株的共同特征,这一发现不仅可以降低MS的培养成本,还可以引起人们对亚洲MS菌株的研究兴趣在CARD数据库,作者预测到、等多个氟喹诺酮类耐药基因,MIC结果也验证了MS-FJ01对其耐药,这可能是由于临床上常使用大环内酯类、氟喹诺酮类抗生素对MS感染进行治疗,从而导致MS对氟喹诺酮类抗生素耐药性严重。此外,作者注释到MsbA具有多药耐药性,MsbA是一种多药耐药转运蛋白同源物,属于含有三磷酸腺苷结合盒的转运蛋白超家族,该盒也被称为核苷酸结合域(NBD)。MsbA是革兰阴性菌中必不可少的ATP结合盒转运蛋白,可将脂质A和脂多糖从细胞质小叶运输到内膜的周质小叶,而脂质A是细菌细胞外膜的主要成分,也是唯一对细胞活力至关重要的细菌ABC转运蛋白,可以作为新抗生素的可行靶点进行深入研究。
在CAZy库,作者注释到4个基因,其中,3个为糖苷水解酶,研究表明,糖苷水解酶通过水解糖苷键,释放糖类产物,用于生物体代谢途径。糖基转移酶参与细胞内多种生命活动,将体内活性物质的单糖部分转移到蛋白质、脂质、糖和核酸中形成糖基化。这些酶在生物体的次级代谢活动中起着不可替代的作用。
支原体的侵袭性由表面的黏附素、辅助蛋白、荚膜多糖、侵袭性酶和生物被膜介导;而黏附是支原体感染过程中的初始步骤,也是确保它们在宿主体内定植和传播的重要步骤。
在MS-FJ01基因组中挖掘出、、、、、、、等与黏附素相关的基因,这些都与其他支原体相同,如肺炎支原体、牛支原体、猪肺炎支原体、鸡毒支原体等。在VFDB数据库,注释到相似度最高的33号基因(基因相似性41.67%)编码磷酸胆碱,磷酸胆碱可以作为HepI或HepIII的末端部分掺入并修饰脂多糖,通过与上皮细胞上的血小板激活因子 (PAF) 受体结合而增加了对抗菌肽(人导管素LL-37)的抗性与黏附性;由于磷酸胆碱表达是相位可变的,因此可以促进逃避抗原特异性宿主免疫防御,并在不同宿主微环境中定植。
4 结 论
从福州鸡场疑似感染鸡滑液囊支原体的病鸡中分离到一株福建鸡滑液囊支原体菌株MS-FJ01,通过三代联合二代测序技术对MS-FJ01进行全基因组测序,经过拼接后得到细菌基因组完成图,测序结果显示MS-FJ01全长为795 381 bp,GC含量为28.39%;预测编码基因数为704个,GC含量为28.64%;共鉴定出34个tRNA和7个rRNA;通过数据库对组装基因进行COG、GO和KEGG富集及分析,其中,在COG数据库注释到476个基因,GO数据库注释到382个基因,KEGG数据库注释到425个基因。在抗生素抗性基因中发现的多药耐药基因MsbA可能成为新抗生素的潜在靶点;在毒力基因中发掘到多个与黏附相关的基因,其在MS入侵宿主中可能有着至关重要的作用;对这些基因的注释将有助于全面认识MS致病相关基因及其与宿主互作过程中的致病性。
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年7期)2022-04-22
山地农业生物学报(2022年1期)2022-01-26
科学导报(2021年29期)2021-06-03
科海故事博览·下旬刊(2019年6期)2019-04-16
植物保护(2018年1期)2018-05-14
人人健康(2017年19期)2017-10-20
大自然探索(2017年8期)2017-08-28
湖北农业科学(2014年22期)2015-01-20
中国民族民间医药·下半月(2014年2期)2014-09-26
