
不同温度对油樟种子生理及萌发的影响
2022-11-15 03:32赵鑫王慧娇何秀燕严悦刘孟婕廖世宏魏琴
中国野生植物资源 2022年10期
赵鑫,王慧娇,何秀燕,严悦,刘孟婕,廖世宏,魏琴*
(1.宜宾学院,四川 宜宾 644000;2.四川省油樟工程技术研究中心,四川 宜宾 644000;3.宜宾梦想农旅发展有限公司,四川 宜宾市 644000)
油樟[Cinnamomum longipaniculatum(Gamble)N.Chaoex H.W.Li]是樟科樟属常绿乔木,根、茎、叶、果均可提取精油,在香料香精、医药卫生、油脂化工、园林绿化等多方面具有重要的应用价值。油樟以扦插、压条[1]等方式繁殖,但扦插苗根系生长弱影响植株生长[2],因此目前油樟以实生繁殖为主。而油樟种子萌发率低,仅30%左右[3],种子沙藏保存时间有限,且易引起种子活力弱、出苗差等问题[4]。因此,筛选适宜的油樟种子贮藏温度及有效提高油樟种子萌发成为促进油樟产业发展亟待解决的问题。
种子萌发不仅与多种内在因素相关[5],还受各种环境因素的影响,如湿度、空气、温度等密切相关[6],其中温度是影响种子萌发的主要因素之一[7]。低温能有效延长种子贮藏时间[8],不同植物对于低温催芽的最适温度范围差异很大[9]。低温干燥有助于种子长期保存[10]。含水量低的种子发芽效果更好[11]。但温度过低会使种子丧失发芽能力[12]。此外,超氧阴离子自由基的积累能够引起种子劣变[13]。因此种子内部各种生理活性物质,如超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)、过氧化氢酶(Catalase,CAT)活性、丙二醛(Malondialdehyde,MDA)含量、相对电导率、可溶性蛋白含量等[13-16]都对种子萌发及活力有很大的影响。
油樟种子为顽拗性种子[17],主要集中分布在热带区域,而目前热带植物种子贮藏和研究工作还很少[18]。因此本研究以宜宾油樟种子为材料,采用不同温度进行处理,测定种子的各项生理指标以及相关萌发率,为探究油樟种质保存与开发奠定基础。
1 材料与方法
1.1 材料
本试验油樟种子2020年9月采收后沙藏保存于宜宾梦想农旅发展有限公司油樟基地。挑选大小均一且饱满的种子进行不同温度处理。设置4个温度梯度:-20℃、2℃、15℃、55℃,以室温15℃为对照(CK)。-20℃、2℃处理5天,55℃以温汤浸种处理,15℃普通沙藏处理。每个处理均设三个生物学重复,每个重复200粒种子。种子处理后,种入基质,统计第25天的发芽率。
1.2 方法
SOD活性采用氮蓝四唑法测定[19]。POD活性采用愈创木酚法测定[20]。CAT活性采用紫外分光光度法测定[21]。MDA活性采用硫代巴比妥酸法测定[22]。相对电导率采用电导率仪法测定[23]。可溶性蛋白含量采用考马斯亮蓝G-250法测定[24]。种子含水量采用烘干称重法测定[25]。种胚形态变化观察采用石蜡切片法观察[26]。用Excel 2010及Origin-Pro 9.0进行数据处理及做图,采用SPSS 20.0软件进行方差分析(P<0.01)。
2 结果与分析
2.1 不同温度对种子萌发率的影响
不同温度处理25天后,油樟种子发芽率差异显著(表1)。2℃处理种子发芽率最高,为28%,显著高于其他处理。15℃与55℃处理无显著性差异,-20℃无种子发芽。

表1 不同处理后种子发芽率Tab.1 The germination rate of different treatments
2.2 油樟种子形态与解剖结构变化
2.2.1 油樟种子外形及胚乳内含物
油樟种子呈球形或扁园形,黑褐色,种皮有许多线条成网状花纹,微凹,经过沙藏后呈棕色,种皮坚硬且厚,中央有种脊环绕,并向外突出(图1A)。油樟种子由种皮、胚乳、胚3个部分组成,肉质胚乳,胚较小,长约0.2~0.4 cm(图1B)。利用碘液染色,胚乳细胞基本无颜色变化,说明油樟胚乳细胞中不含淀粉(图1D);考马斯亮蓝染色胚乳细胞后,胚乳靠外层细胞内含物变蓝,表明部分胚乳细胞含有蛋白体(图1E),在靠近种皮附件环状分布;苏丹Ⅲ染色后胚乳细胞中富含红色的脂滴(图1F),且胚乳细胞越靠近种皮,脂滴含量越高。结果表明油樟种子中含有脂滴和蛋白体,基本不含淀粉(图1C)。
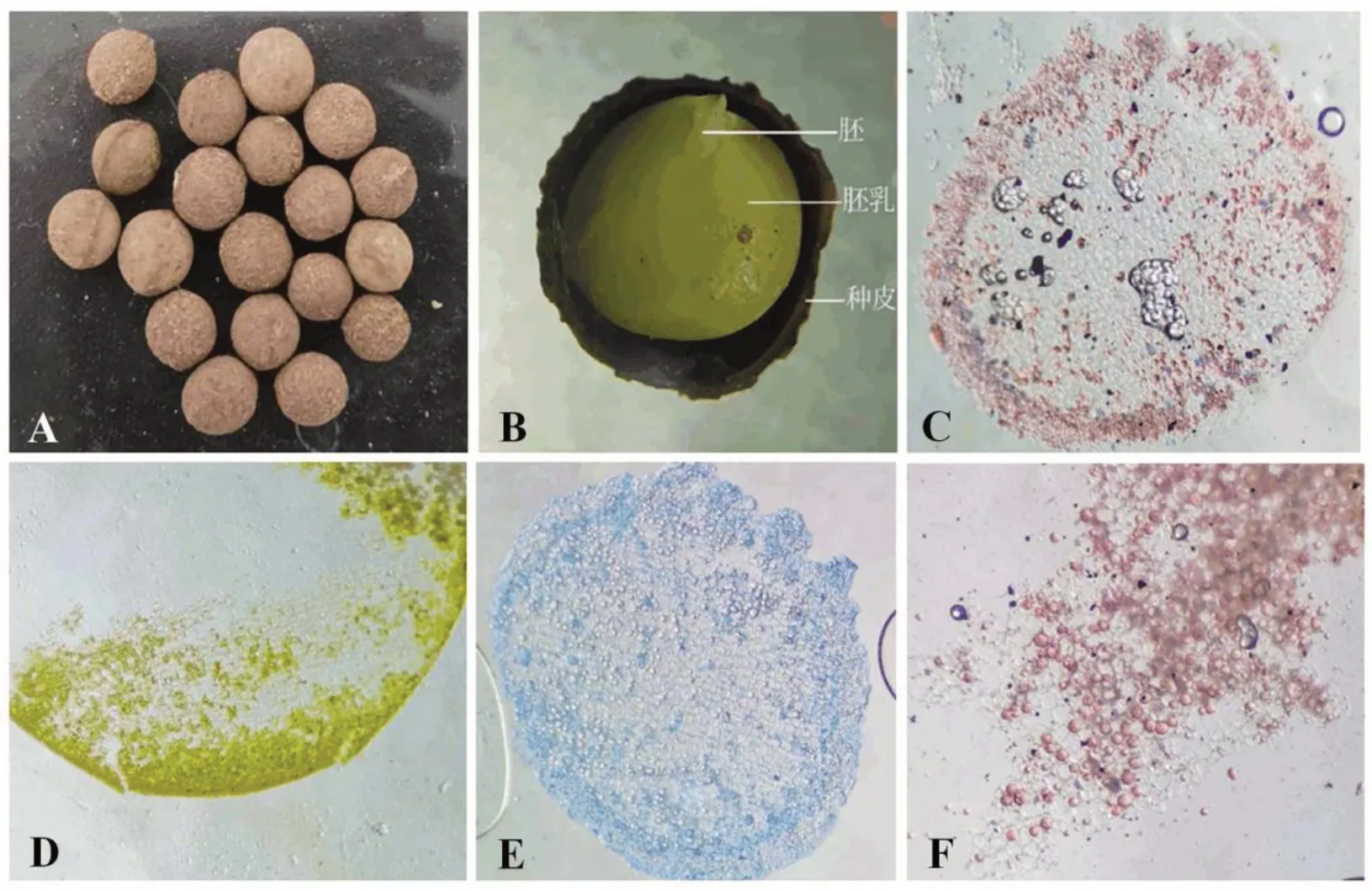
图1 油樟种子形态及胚乳结构Fig.1 The seed morphology and endosperm structure
2.2.2 不同温度对油樟胚及胚乳的影响
油樟为双子叶植物,种子由胚、胚乳及种皮组成,胚较小。不同温度处理后观察油樟胚的变化,2℃与55℃处理胚变化明显,-20℃处理后胚无明显变化(图2)。

图2 油樟种子胚的形态Fig.2 The embryo morphology of C.longepaniculatum seed
石蜡切片观察CK和2℃处理油樟种子胚乳结构。胚乳由薄壁细胞构成,番红固绿染色后细胞壁等为绿色,脂类物质呈红色(图3)。通过对比可知2℃处理后胚乳细胞排列较为松散,细胞壁与CK相比有明显的差异,胚乳细胞中油脂含量减少(图3 C、D)。

图3 不同温度处理后油樟种子胚乳的变化Fig.3 Changes of endosperm under different treatment
2.2.3 不同温度处理对种子含水量、可溶性蛋白及相对电导率的影响
不同温度处理后油樟种子含水量差异显著(图4A),2℃处理油樟种子含水量为14.87%,显著低于其他处理。-20℃处理含水量为18.25%,与对照17.35%无显著性差异。55℃处理含水量可达23.86%。2℃处理含水量显著低于其他处理,结果表明2℃能够有效维持种子活力。
不同温度处理后种子可溶性蛋白含量差异显著(图4B)。2℃和55℃处理种子可溶性蛋白含量均显著高于对照,其中55℃处理含量最高,达358.64 mg·g-1,2℃处理为261.61 mg·g-1。-20℃处理可溶性蛋白含量最低,为187.53 mg·g-1,相比CK降低了13.93%。在种子贮藏过程中大分子蛋白质降解为小分子的可溶性物质,为种子生长发育提供所需营养物质,较高含量的可溶性蛋白可以使植物的抗逆能力增强。本研究结果表明:2℃和55℃处理油樟种子可溶性蛋白含量高,对种子活力的保持或抗逆性的提升最佳。
不同温度处理后种子中相对电导率差异显著(图4C)。与其他处理相比,2℃处理相对电导率最低,为43.68%。-20℃与55℃处理电导率分别为56.61%、49.78%,显著高于对照。结果表明:2℃处理相对电导率最低,细胞膜完整性破坏程度最轻,膜功能受损程度最小,细胞内物质渗漏最少。

图4 不同温度对种子含水量、可溶性蛋白及相对电导率的影响Fig.4 Effects of different temperature on water content soluble protein and relative conductivity of seeds
2.2.4 不同温度处理对种子抗氧化酶活性与丙二醛的影响
种子SOD活性在不同温度处理后显著变化(图5A)。其中-20℃处理显著低于对照,为245.20 U·g-1。而2℃与55℃处理种子SOD活性显著上升,分别为420.50 U·g-1、436.09 U·g-1。与对照相比,2℃和55℃处理后均能显著提高种子SOD活性。种子POD活性在不同温度处理后显著变化(图5B)。2℃处理POD活性最高,可达108.30 U·g-1。55℃、-20℃处理与对照POD活性分别为63.90 U·g-1、32.00 U·g-1、46.40 U·g-1,显著低于2℃处理。本研究结果表明:2℃处理显著提升种子POD活性。不同温度处理后种子CAT活性差异显著(图5C)。2℃处理CAT活性为88.20 U·g-1,显著高于其他处理。-20℃处理CAT活 性 为59.30 U·g-1。对 照CAT活 性 最 低,为13.30 U·g-1。不同温度处理后种子MDA含量差异显著(图5D)。2℃处理MDA含量最低,为13.21 μmol·g-1,显著低于其他处理。而55℃处理后MDA含量可高达35.39 μmol·g-1,显著高于其他处理。-20℃与CK无显著性差异。-20℃和55℃处理后的种子,MDA含量大于室温贮藏,种子内膜脂质过氧化程度更严重。本研究结果表明:2℃处理MDA含量最低,该处理对种子伤害最小。

图5 不同温度对种子SOD、POD、CAT及MDA的影响Fig.5 Effects of different temperature on SOD,POD,CAT and MDA of seeds
3 讨论与分析
3.1 油樟种子内含物及萌发
油樟种子为顽拗性种子[17],油樟种子种皮较厚,对种子具有机械保护的同时影响种子的正常萌发。胚乳中富含油脂与蛋白质。在适宜温度处理下,胚乳细胞发生形态与内含物的变化,是由于脂类物质降解并重新利用引起的[27]。
当种子含水量减少时,呼吸减缓,能量消耗可维持在机体最低水平,有助于维护种子活力。所以种子含水量越低,活力越高[28]。2℃处理油樟种子含水量最低,能更有效维持种子活力。而55℃温水处理后,种子含水量增加并开始萌发,说明温汤浸种可能促进油樟种子萌发。生长25天后2℃处理的种子萌发率显著高于55℃处理,表明油樟种子萌发与温汤浸种无明显关系。
种子活力稳定的基础来源于种子内贮藏物质的合成与积累,种子活力会随着贮藏物质的消耗而逐步下降[8],且较高含量的可溶性蛋白质可以使植物的抗逆性得到增强[29-30]。本研究中2℃和55℃贮藏的种子可溶性蛋白含量明显高于其他处理种子的含量,2℃低温处理种子代谢弱,消耗贮藏物质少,有利于贮藏物质的积累,因此有利于种子活力的保持;而55℃处理后的油樟种子可溶性蛋白含量增多明显,这表明种子开始萌发后将大分子蛋白质降解为小分子的可溶性物质,为种子生长发育提供所需营养物质[3]。
相对电导率可反映植物细胞膜在逆境条件的受损伤程度[31]。油樟种子在2℃处理相对电导率最低,细胞膜完整性破坏程度最轻,膜功能受损程度最小,细胞内物质渗漏最少。
3.2 抗氧化酶活性与种子活力
植物体主要的抗氧化酶为SOD、POD、CAT,抗氧化酶系统各种酶之间相互配合,相互协同,在清除自由基的过程中起作用。SOD是广泛存在于植物体内能消除O2-对植物细胞的损害,有效维持细胞内活性氧代谢平衡[32]。
SOD可清除机体过多活性氧,使植物对逆境胁迫的耐受能力提高。SOD活性高低与种子劣变及种子的活力密切相关。种子内SOD活性越高,种子抗逆性越强,活力也越高。油樟种子经过2℃和55℃温度处理后,SOD活性显著升高,能有效清除种子内部活性氧,使油樟种子保持较高的生物活性,抗逆性得到增加。POD能有效阻止植物体内活性氧系统中的O2-和H2O2积累,防止细胞受自由基的毒害,能有效保护植物体内酶活性,同时还可抑制膜脂过氧化,延缓细胞衰老进程[33]。同时,POD活性高低与质膜损伤造成的种子劣变及种子的活力也密切相关。POD活性越高,种子抗逆性越强,种子活力越高。油樟种子经过2℃处理后POD活性最高,所以该处理能有效阻止O2-和H2O2的积累,提高油樟种子抗逆性。CAT能防止过氧化氢在机体内积累,高活性的CAT能够使种子得到较好的保护,维持其生物活性。CAT活性越高,种子抗逆性越强,种子活力越高。油樟种子经2℃处理CAT活性最高,因此低温能有效阻止H2O2在油樟种子内部的积累,有效保护油樟种子,增加种子抗逆性。种子的生理生化响应主要与较高的SOD和POD活性有关[14]。适当低温处理,油樟种子SOD活性升高,延缓油樟种子衰老,保持种子生物活性,促进种子萌芽。2℃低温处理保持较高的抗氧化酶活性来清除贮藏过程中积累的自由基,以防御氧化损伤,维持种子活力。
MDA含量与相对电导率能反映种子的劣变程度[34]。MDA是膜脂过氧化的分解产物,可反映细胞膜的受伤害程度[35]。相对电导率是反映植物细胞膜透性的一项基本指标[36],相对电导率越高,细胞膜受损程度越深。2℃低温贮藏下种子MDA含量与相对电导率与室温贮藏的油樟种子相比均显著降低,能较好地保护细胞膜结构不受破坏,种子活力维持较好。
4 结论
本试验用不同温度处理油樟种子,通过对种子含水量、可溶性蛋白、相对电导率、MDA、CAT、POD、SOD以及种胚发育结构的研究,结果表明:2℃能够显著提高种子SOD、POD、CAT活性,MDA含量、相对电导率和含水量最低,而且可溶性蛋白含量能保持较高水平。综上所述,2℃处理能够有效保持种子活性,提高种子萌发过程中的各项生理指标,利于油樟种子贮藏与萌发。
猜你喜欢
今日农业(2022年6期)2022-07-05
干旱地区农业研究(2022年3期)2022-05-24
矿冶工程(2021年4期)2021-09-15
三农资讯半月报(2021年2期)2021-03-22
辽宁农业科学(2021年1期)2021-03-17
世界有色金属(2020年15期)2020-10-10
科学导报(2020年8期)2020-03-12
小学阅读指南·高年级版(2016年9期)2016-10-31
现代园艺(2016年2期)2016-03-15
湖北农业科学(2014年13期)2014-08-28
