
小豆WRKY基因家族鉴定及应答锈菌侵染的表达分析
2022-12-22 14:03滑艳敏杨阳孙伟娜殷丽华柯希望穆娟微左豫虎
黑龙江八一农垦大学学报 2022年6期
滑艳敏,杨阳,2,孙伟娜,殷丽华,柯希望,穆娟微,左豫虎
(1.黑龙江八一农垦大学农学院/黑龙江省作物有害生物互作生物学及生态防控重点实验室,国家杂粮工程技术研究中心,大庆 163319;2.四川省广元市昭化区农业农村局;3.黑龙江省农垦科学院植物保护研究所)
小豆(Vigna argularis)是我国重要的杂粮作物,在全国各地均有种植,黑龙江省是我国小豆的主要产区[1]。据统计2018年黑龙江省小豆产量达11.90万t,占全国小豆总产量的42.82%[2]。然而,由豇豆单胞锈菌(Uromyces vignae)引起的小豆锈病[3],在我国各小豆种植区均有发生,对小豆的产量和品质产生了严重影响。挖掘小豆抗锈病基因,培育及合理利用抗性品种是小豆锈病可持续控制的关键。
WRKY转录因子是植物中重要的转录因子家族,在植物应答生物、非生物胁迫中起到了重要的调控作用[4]。目前在高粱(Sorghum bicolor)[5]、向日葵(Helianthus annuus)[6]、甜瓜(Cucumis melo)[7]等众多植物基因组中均鉴定到了WRKY家族基因。高度保守的WRKY结构域是WRKY基因家族的典型结构特征,WRKY结构域长度约60个氨基酸残基,包含位于N端的七肽域和位于C端的锌指结构,在WRKY转录因子中,锌指结构有C2H2(CX4-5CX22-23HXH)和C2HC(CX7CX23HXC)两种类型[8]。根据WRKY结构域的数量和锌指结构类型,WRKY家族可分为GroupⅠ、Ⅱ、Ⅲ等3个亚家族,其中GroupⅠ包含2个典型的WRKY结构域,分别位于N-端和C-端;GroupⅡ和GroupⅢ通常仅有1个WRKY结构域,二者的区别在于GroupⅡ的锌指结构为C2H2型,而GroupⅢ为C2HC型。此外,根据进化关系,GroupⅡ可进一步分为Ⅱ-a、Ⅱ-b、Ⅱ-c、Ⅱ-d、Ⅱ-e等5个亚组[9]。
大量研究表明,WRKY可通过激活或抑制植物防卫反应基因的表达,进而调节植物对生物或非生物逆境的适应性[10]。如在水稻中,过量表达WRKY家族GroupⅡ-a亚家族成员OsWRKY62.1可显著抑制防卫反应基因的表达,从而减弱水稻对白叶枯病菌(Xanthomonas oryzae pv.oryzae,Xoo)侵染的免疫反应,OsWRKY62在水稻抗白叶枯病中发挥负调控作用[11]。然而,Vo K等[12]研究发现,在水稻中过表达WRKY基因OsWRKY67可增强水稻的抗病能力,在水稻应答白叶枯病菌和稻瘟病菌(Magnaporthe oryzae)侵染过程中发挥正调控作用。最近的研究发现,在小麦中过量表达大麦WRKY转录因子HvWRKY6和HvWRKY70均不同程度提高了小麦对条锈菌(Puccinia striiformis)和白粉菌(Blumeria gramnis)的抗性[13]。上述研究表明,WRKY转录因子在植物抵抗生物胁迫过程中发挥重要的调控作用。
然而,有关小豆基因组中WRKY转录因子家族基因的数量、结构特点及其在小豆应答锈菌侵染过程中的作用仍不十分明确。因此,为深入探索小豆WRKY转录因子在抗锈病中的功能,以小豆全基因组[14]为参考,利用生物信息学方法,分析了小豆基因组中WRKY转录因子的数量及其序列特征,并比较分析了小豆WRKY与其他物种WRKY基因家族的进化关系,以期从分子进化的角度深入认识小豆WRKY基因家族的特征及功能。在此基础上,利用课题组前期获得的抗病品种“QH1”接种后24 h和48 h的转录组数据,初步明确了小豆WRKY家族成员在小豆抗锈病过程中的表达模式,为深入解析小豆抗锈病的分子机制提供理论依据。
1 材料与方法
1.1 小豆WRKY基因家族鉴定及染色体定位
小豆全基因组数据下载自NCBI数据库(http://www.ncbi.nlm.nih.gov/)[15]。参考苹果[16]和甜菜[17]WRKY基因家族鉴定的方法,鉴定小豆全部WRKY基因。以基因组注释信息为依据,利用TBtools[18]对小豆WRKY基因染色体定位进行可视化。
1.2 小豆WRKY基因家族序列特征及系统发育分析
利用DNAMAN8.0分析小豆WRKY基因的保守域及序列特征,参考Rushton等[8]和Eulgem等[9]的分类依据,对小豆WRKY基因家族成员进行分类。并以拟南芥WRKY家族GroupⅡ亚家族成员的蛋白序列(序列下载自https://www.arabidopsis.org/index.jsp)为参考,利用MEGA 7软件,采用Neighbor-Joining(NJ)(Bootstrap=1 000)法构建系统发育树,明确小豆WRKY GroupⅡ亚家族成员的分类。
1.3 小豆WRKY基因家族成员在小豆抗锈病过程中的表达分析
为初步了解WRKY转录因子在小豆抵抗锈菌侵染中的作用,从NCBI下载课题组前期获得的高抗锈病品种“QH1”接种后24和48 h的转录组数据,数据下载自NCBI SRA数据库(数据编号:SRP130843,链接:https://www.ncbi.nlm.nih.gov/sra/?term=SRP130843),用于分析小豆WRKY基因家族成员在小豆应答锈菌侵染过程中的表达模式。
2 结果与分析
2.1 小豆WRKY基因家族成员鉴定及染色体定位分析
小豆全基因组中共有90个WRKY转录因子,蛋白序列特征分析表明,90个WRKY转录因子有47个呈酸性,42个呈碱性,1个呈中性。分子量在16.0~65.3 kDa之间,氨基酸序列长度在139~600 aa之间,理论等电点在4.91~9.99之间。染色体定位结果显示,90个成员中有18个定位未能组装到染色体的Scanffold上,其余成员在小豆11条染色体(Chromosome,Chr)上均有分布,其中Chr3分布数量最多为12个,最少的为Chr8仅有1个(图1)。
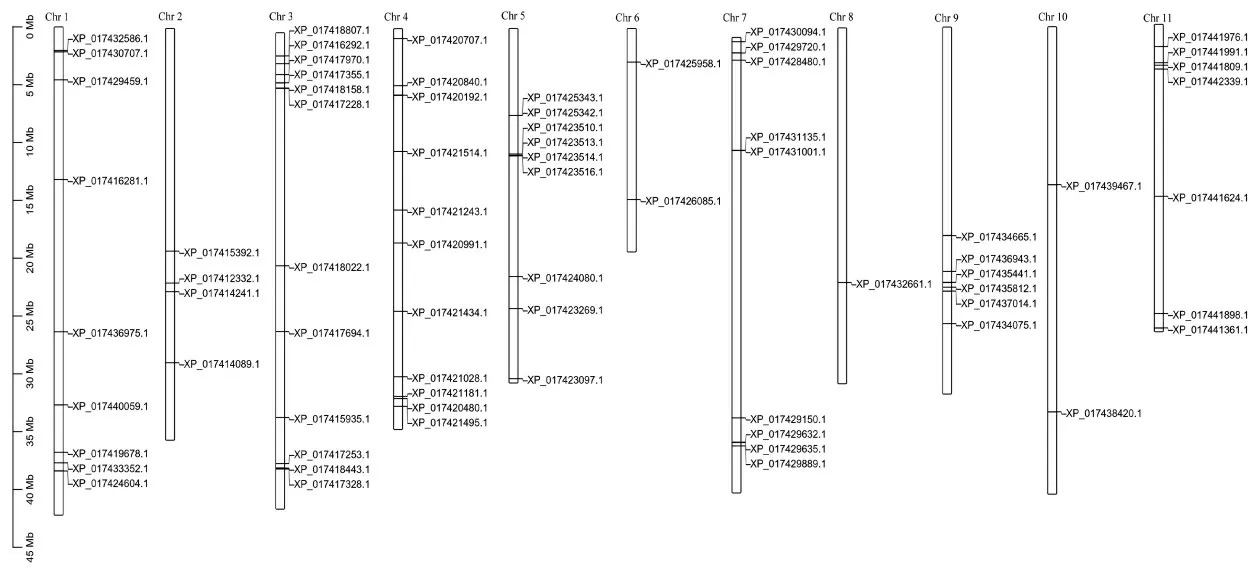
图1 小豆WRKY基因家族染色体定位Fig.1 Chromosome distribution of WRKY gene in Vigna angularis
2.2 小豆WRKY序列特征分析及基因家族分类
利用DNAMAN软件,对小豆WRKY基因家族成员的氨基酸序列进行多序列比对,依据WRKY结构域的数量及锌指结构类型,小豆90个WRKY家族成员可分为GroupⅠ、Ⅱ、Ⅲ等3个亚家族。其中GroupⅠ有17个成员,GroupⅡ有61个成员,GroupⅢ有12个成员。对不同亚家族成员蛋白序列的特征分析发现,与保守的WRKY七肽域氨基酸序列[WRKYGQK]相比,小豆WRKY家族成员在七肽域的多个位点都存在不同类型的氨基酸取代或丢失(表1)。如GroupⅠ成员XP_017434665.1的C端七肽域发生了整体丢失(图2,红色方框),XP_017441624.1序列N端的WRKY七肽域第六位的谷氨酰胺(Q)被谷氨酸(WRKYGEK)取代。除七肽域存在氨基酸取代外,该亚家族成员的C端锌指结构存在一个氨基酸的插入,由CX4CX22HXH变为CX4CX23HXH(图2);与GroupⅠ相似,GroupⅡ部分成员七肽域第二位点的精氨酸(R)被赖氨酸(WKKYGQK)取代,或第六位的谷氨酰胺(Q)被赖氨酸(WRKYGKK)取代。GroupⅡ个别成员C2H2型锌指结构(CX4-5CX22-23HXH)序列中的半胱氨酸(C)和组氨酸(H)中间存在缺失(图3,红色方框),或在两个组氨酸(H)中间存在1个单插入(图3,红色箭头)等变异类型;而GroupⅢ亚家族成员的七肽域则没有发生氨基酸取代,都为典型的WRKYGQK,但个别成员的C2HC型锌指结构(CX7CX23HXC)序列存在变异,如XP_017408549.1的锌指结构在两个半胱氨酸(C)中间存在单个氨基酸的缺失(图4,红色方框)。

表1 小豆WRKY家族成员七肽域和锌指结构的氨基酸变异类型Table 1 Amino acid substitution types in heptapeptide domain and zinc finger structure of WRKY family members in Vigna angularis

图2 小豆WRKY基因家族GroupⅠ亚家族成员氨基酸序列特征Fig.2 Characteristics of the amino acid sequences of Vigna angularis WRKY gene family GroupⅠsubfamily members
采用MEGA7软件,利用邻接法(Neighbor-Joining)构建小豆90个WRKY转录因子的系统发育树(图5)显示,GroupⅢ和GroupⅠ亚家族成员分别聚为一簇,而GroupⅡ亚家族成员则较为分散,研究表明,根据氨基酸数量的差异GroupⅡ又分可为多个亚类,为进一步确定小豆WRKY基因家族GroupⅡ的分类,将已知类型的拟南芥WRKY GroupⅡ成员的氨基酸序列与小豆GroupⅡ成员共同构建系统发育树(图6),结果表明,小豆GroupⅡ的61个成员明显被区分为Ⅱ-a(6个)、Ⅱ-b(16个)、Ⅱ-c(21个)、Ⅱ-d(7个)和Ⅱ-e(11个)5个亚类(图6),其中GroupⅡ-a和GroupⅡ-b,GroupⅡ-d和GroupⅡ-e亲缘系更近,GroupⅡ-c的不同成员在进化树中分布在3个不同的枝上。

图3 小豆WRKY基因家族GroupⅡ亚家族成员氨基酸序列特征Fig.3 Characteristics of the amino acid sequences of Vigna angularis WRKY gene family GroupⅡsubfamily members
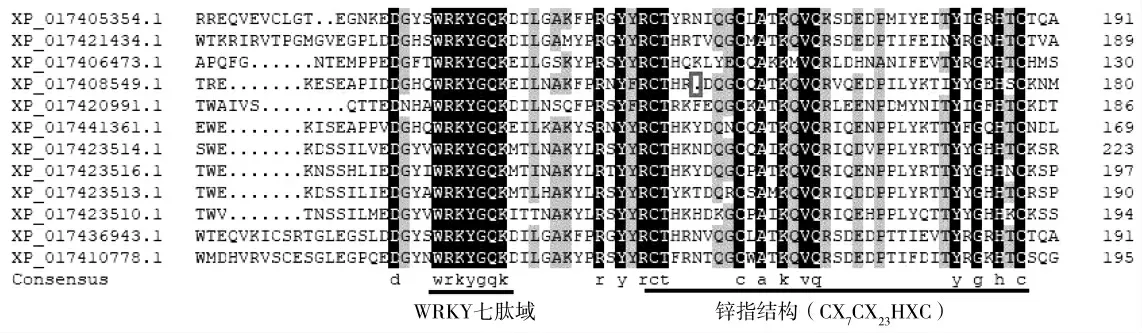
图4 小豆WRKY基因家族GroupⅢ亚家族成员氨基酸序列特征Fig.4 Characteristics of the amino acid sequences of Vigna angularis WRKY gene family GroupⅢsubfamily members

图5 小豆WRKY基因家族成员系统发育分析Fig.5 Phylogenetic analysis of the WRKY gene family members of Vigna angularis
2.3 小豆WRKY成员应答锈菌侵染的表达模式
为初步了解小豆WRKY基因家族在小豆抗锈病中的作用,利用1.3中的转录组数据(SRP130843)对90个小豆WRKY家族成员的表达分析发现,90个基因中有82个基因在不同程度表达,依据各基因RPKM(Reads Per Kilobase per Million mapped read)值对82个WRKY成员在抗病品种中应答锈菌侵染的表达模式进行聚类后发现,82表达的WRKY成员中,接菌后24 h上调表达的有16个(图7-①);接菌后24 h和48 h均上调表达的有18个(图7-②);接菌后24和48 h下调表达的有4个(图7-③);接种后48 h上调表达的成员29个(图7-④);其余15个成员的表达不受锈菌侵染的影响,但接种无菌水后48 h的表达水平高于接种无菌水后24 h(图7-⑤),暗示这些成员可能在小豆生长过程中发挥重要作用。
3 结论与讨论
研究基于小豆基因组数据,系统分析了小豆WRKY基因家族成员的数量、结构特征及其应答锈菌侵染的表达模式,明确了小豆基因组中共包含90个WRKY成员,依据结构特征及进化关系,小豆WRKY基因家族分化为GroupⅠ、Ⅱ和Ⅲ等3个亚家族,蛋白序列特征分析表明,小豆WRKY家族成员在保守的WRKY结构域上存在多种变异,且七肽域上的氨基酸取代常发生于第6个氨基酸位点,而锌指结构的变异则以个别位点的氨基酸插入或缺失为主。基于转录组数据的表达模式分析表明,90个WRKY成员中有82个成员不同程度表达,其中63个成员的表达受病原菌侵染的影响,表明WRKY家族基因在小豆应对锈菌侵染过程中发挥了重要的调控作用。
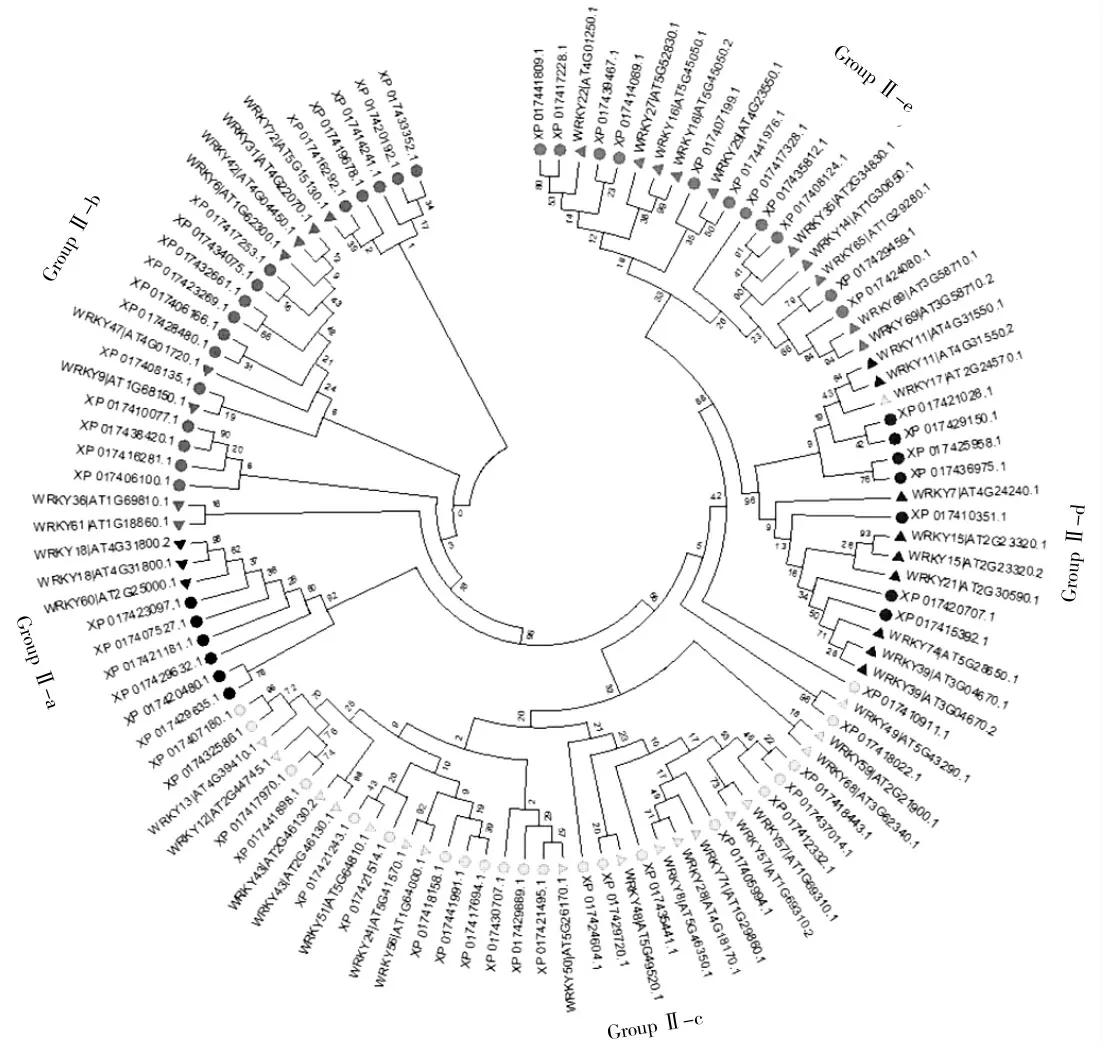
图6 小豆WRKY家族GroupⅡ成员与拟南芥WRKY家族GroupⅡ成员的系统发育分析Fig.6 Phylogenetic analysis of Group II members of Vigna angularis WRKY family and GroupⅡmembers of WRKY family in Arabidopsis thaliana
WRKY转录因子作为植物中最大的基因家族之一[10],已在多种植物的基因组中被鉴定。从成员数量看,在小豆基因组中共包含90个WRKY家族成员,与菜豆(Phaseolus vulgaris)的WRKY家族成员数量一致[19],高于鹰嘴豆(Cicer arietinum)的70个[20],但低于大豆(Glycine max)的188个[21]。家族成员众多可能是WRKY转录因子在植物中调控发育过程及逆境适应性的基础。研究表明,WRKY基因家族存在明显的家族分化现象,依据WRKY结构域的数量和序列特征,WRKY基因家族常被分为GroupⅠ、Ⅱ、Ⅲ等3个亚家族,且GroupⅡ亚家族进一步分化出了Ⅱa、Ⅱb、Ⅱc、Ⅱd和Ⅱe等5个亚组,如拟南芥[9]、大豆[21]等的GroupⅡ亚家族都存在类似的分化。研究同样发现,小豆90个WRKY成员同样分属于GroupⅠ、Ⅱ、Ⅲ等3个亚家族,且GroupⅡ也存在类似的5个亚组的分化(图6),暗示小豆WRKY家族成员与拟南芥、大豆等植物的WRKY家族存在类似的演化模式。
WRKY转录因子的七肽域和锌指结构是该家族基因发挥功能的关键DNA结合结构域,大量研究表明,该DNA结合结构域的差异是WRKY基因家族在植物中功能多样性的结构基础[10]。研究发现,小豆WRKY家族成员七肽域常在第6位氨基酸处发生取代,且最常见的取代类型为谷氨酰胺(Q)突变为谷氨酸(E)或赖氨酸(K),即WRKYGQK突变为WRKYGEK或者WRKYGKK。与研究结果相似,玉米WRKY家族部分成员七肽域[WRKYGQK]第6位的谷氨酰胺[Q]被赖氨酸[K]取代[22]。除第6位氨基酸发 生取代外,小豆WRKY七肽域第2位的精氨酸(R)常被赖氨酸(K)取代。目前的研究发现,七肽域第2位发生变异的情形在少数WRKY成员中存在,除R被K取 代 外,也 存 在WIKY、WVKY、WKRY和WSKY等变异类型[14]。此外,小豆WRKY家族成员还出现了N-端七肽域缺失的情形,该类突变在其他物种中目前报道较少。综上所述,小豆WRKY家族成员在关键结构域上变异的多样性,预示其在小豆生长发育及逆境应答中通过调控不同基因的表达以发挥作用。
WRKY作为在植物中调控免疫的重要转录因子家族,同时具备正向和负向调节植物免疫的功能[4]。如海岛棉GbWRKY003、GbWRKY020、GbWRKY075等在黄萎病菌侵染后下调表达,而GbWRKY020的表达则受黄萎病菌的显著诱导,通过正向或负向调控的作用参与棉花对黄萎病的抗性[23]。同样在辣椒中,CaWRKY58受青枯病菌诱导后表达量下调[24],但CaWRKY6和CaWRKY40则在抗青枯病菌中发挥正调控作用[25]。研究对小豆WRKY成员的表达分析同样显示,90个成员中的80个在小豆应答锈菌侵染过程中不同水平表达,其中受锈菌侵染上调表达的成员34个,下调表达的有33个,表明小豆WRKY家族成员在小豆应答锈菌侵染过程中同样具备正向和负向调控抗病的可能,但WRKY家族基因是如何调控小豆对锈病抗性的仍需要进一步的试验验证。
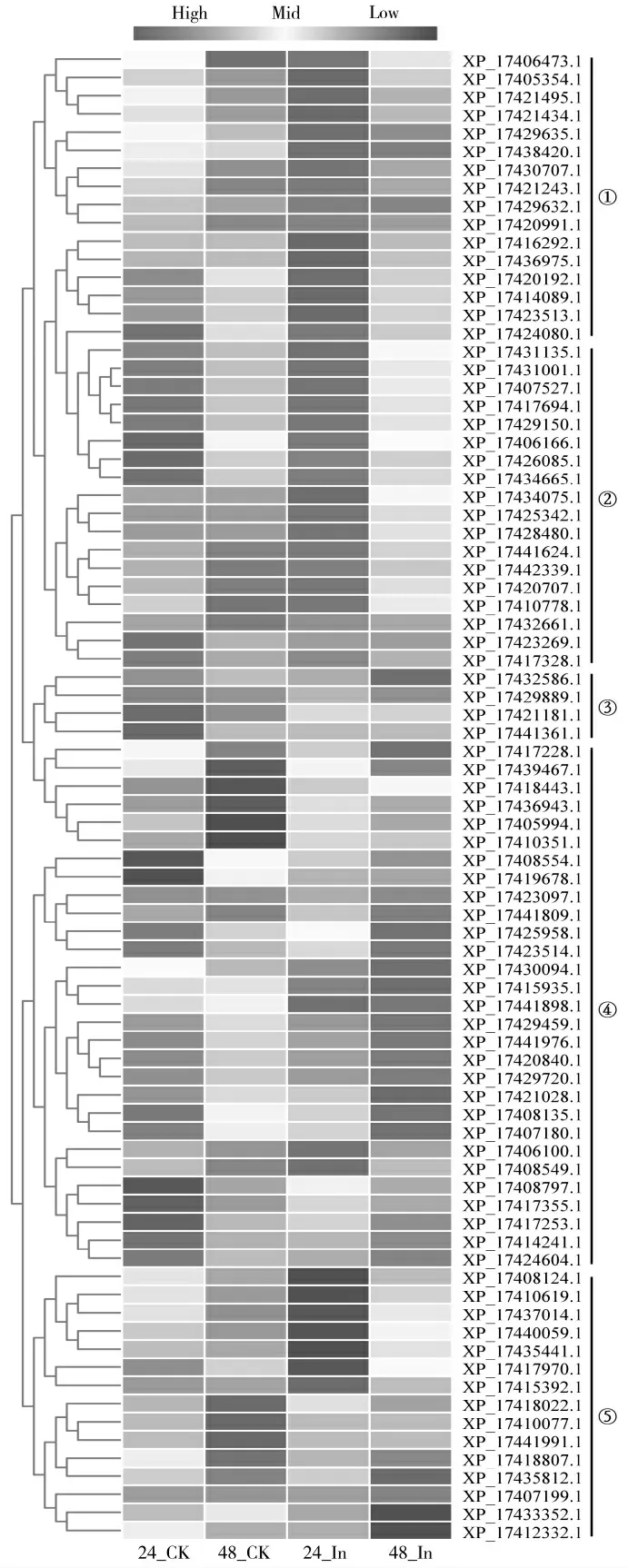
图7 小豆WRKY基因家族成员应答锈菌侵染的表达分析Fig.7 Expression analysis of Vigna angularis WRKY gene family members responding to rust infection
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
当代水产(2022年1期)2022-04-26
河北农业大学学报(2022年2期)2022-04-26
中国农业科学(2021年23期)2022-01-14
昆虫学报(2021年3期)2021-05-11
实用肿瘤学杂志(2020年4期)2020-12-08
中国瓜菜(2020年8期)2020-09-26
莫愁·家教与成才(2016年7期)2016-07-13
医学综述(2011年12期)2011-12-09
当代小说(2009年9期)2009-12-15
