
细胞器之间相互作用在非酒精性脂肪性肝病发生发展中的作用
2023-02-08 13:13刘天会
临床肝胆病杂志 2023年1期
刘天会
首都医科大学附属北京友谊医院 肝病中心, 北京 100050
细胞器可以通过膜接触位点与其他细胞器相互作用,完成物质与信息的交换,形成互作网络[1]。越来越多的研究[2]证实,细胞器互作网络的紊乱与多种疾病的发生发展密切相关。线粒体功能损伤是非酒精性脂肪性肝病(NAFLD)发病机制中重要的一环,在非酒精性肝脂肪变进展为非酒精性脂肪性肝炎(NASH)的过程中发挥重要作用[3-6]。肝细胞中异常增多的脂滴是NAFLD的特征性病理表现,近期,脂滴和其他细胞器之间的相互作用成为细胞生物学研究的热点[7]。本文主要聚焦于NAFLD中线粒体、脂滴与其他细胞器之间的相互作用。
1 细胞器之间相互作用概述
1.1 细胞器之间相互接触在细胞中广泛存在 真核细胞中存在多种由单层或双层膜包围的亚细胞结构——细胞器,如内质网、线粒体、脂滴、高尔基体、溶酶体等。细胞器看似相对独立,各自具有特定的生物学功能。实际上,这些细胞器常常相互接触,通过精细分工、相互协作,维持细胞的正常功能[1]。
应用多光谱荧光图像采集方法,研究者[8]观察了成纤维细胞中细胞器的大小、位置和相互接触等信息,对六种细胞器(内质网、高尔基体、溶酶体、过氧化物酶体、线粒体和脂滴)之间相互接触的频率及动态变化进行了系统描述。结果显示:几乎所有细胞器都可以与其他种类的细胞器形成接触,且这种接触呈现高度动态变化的特点。在细胞生长的不同阶段,不同细胞器之间的接触频率会随时间发生改变。当细胞接受不同外来刺激时,细胞器之间的接触也会发生特定的改变。细胞器之间相互作用依赖于完整的由微管组成的细胞骨架,如果应用微管聚合抑制剂破坏正常的细胞骨架,除了高尔基体与溶酶体的接触增加,其他细胞器之间的相互接触均显著减少[8]。
1.2 细胞器之间相互作用通过膜接触位点实现 细胞器之间的相互作用是通过细胞器之间形成的微小膜连接实现的,这种膜连接被称为膜接触位点 (membrane contact sites,MCS)[9]。MCS通常是由细胞器膜上的特定蛋白互相结合形成的,这些蛋白除了参与形成MCS,通常还具有其他的重要功能。例如,羟甾醇结合蛋白参与形成内质网与高尔基体之间的MCS,同时还具有脂质转移的功能。
通过形成MCS,细胞器之间的距离被拉近,这一距离可以在一定范围内发生变化。例如,在哺乳动物细胞中,内质网和质膜之间的距离在19~22 nm发生动态变化[10];而在酿酒酵母中,这一距离变化范围为17~57 nm[11]。当胞质中钙离子浓度升高时,内质网和质膜之间的距离缩短,提示细胞器之间距离的变化可能具有特殊的意义[10]。另外,根据细胞功能的需要,MCS可以发生动态改变,也可以稳定存在。有的MCS持续时间在10 s以下,有的甚至低于1 s。在胰岛素分泌细胞中,胞质钙离子浓度升高促进内质网蛋白TMEM24磷酸化,导致内质网与质膜之间的MCS解离;而TMEM24蛋白去磷酸化后MCS恢复;上述MCS的动态变化对于胰岛素脉冲式分泌至关重要[12]。在肌肉细胞中,内质网和质膜之间的MCS参与肌肉的兴奋收缩耦联过程,这种MCS可以持续存在[13]。
1.3 细胞器通过相互作用完成重要的生理功能 细胞器之间通过相互作用,可以进行离子、脂质及蛋白质等的交换及转移,还可以参与细胞器生物合成及分裂等重要过程[1,9]。MCS中常常富含脂质转运蛋白家族成员(lipid transfer proteins,LTP),LTP可同时存在于两个细胞器的膜上,形成连接细胞器的桥梁或通道,完成脂质在细胞器之间的转移[14]。内质网膜上的三磷酸肌醇受体与线粒体膜上的电压依赖性阴离子选择性通道蛋白及葡萄糖调节蛋白75之间结合形成MCS,调控内质网和线粒体之间的钙离子传输,维持线粒体钙离子稳态[9]。
内质网上的肌动蛋白成核因子2和线粒体上的与甲酸结合的肌动蛋白成核蛋白1C在内质网-线粒体接触位点结合,促进肌动蛋白-肌球蛋白复合物形成,将动力相关蛋白1募集至线粒体外膜,促进线粒体收缩并完成裂变[15-16]。Wong等[17]的研究表明,线粒体裂变发生在线粒体、内质网和溶酶体三种细胞器之间的连接处,溶酶体上的Ras相关GTP结合蛋白7可促进溶酶体与线粒体接触,参与线粒体分裂。另有研究[18]显示内质网与线粒体相互作用与线粒体的融合也密切相关。
2 线粒体和细胞器之间相互作用与NAFLD
线粒体是细胞中糖类、脂肪和氨基酸最终氧化释放能量的场所,其功能障碍是NAFLD发病的关键环节。越来越多的研究[19-20]发现,线粒体可以与多种细胞器相互作用(图1),参与钙离子稳态失衡、内质网应激、胰岛素抵抗等NAFLD相关的病理过程。
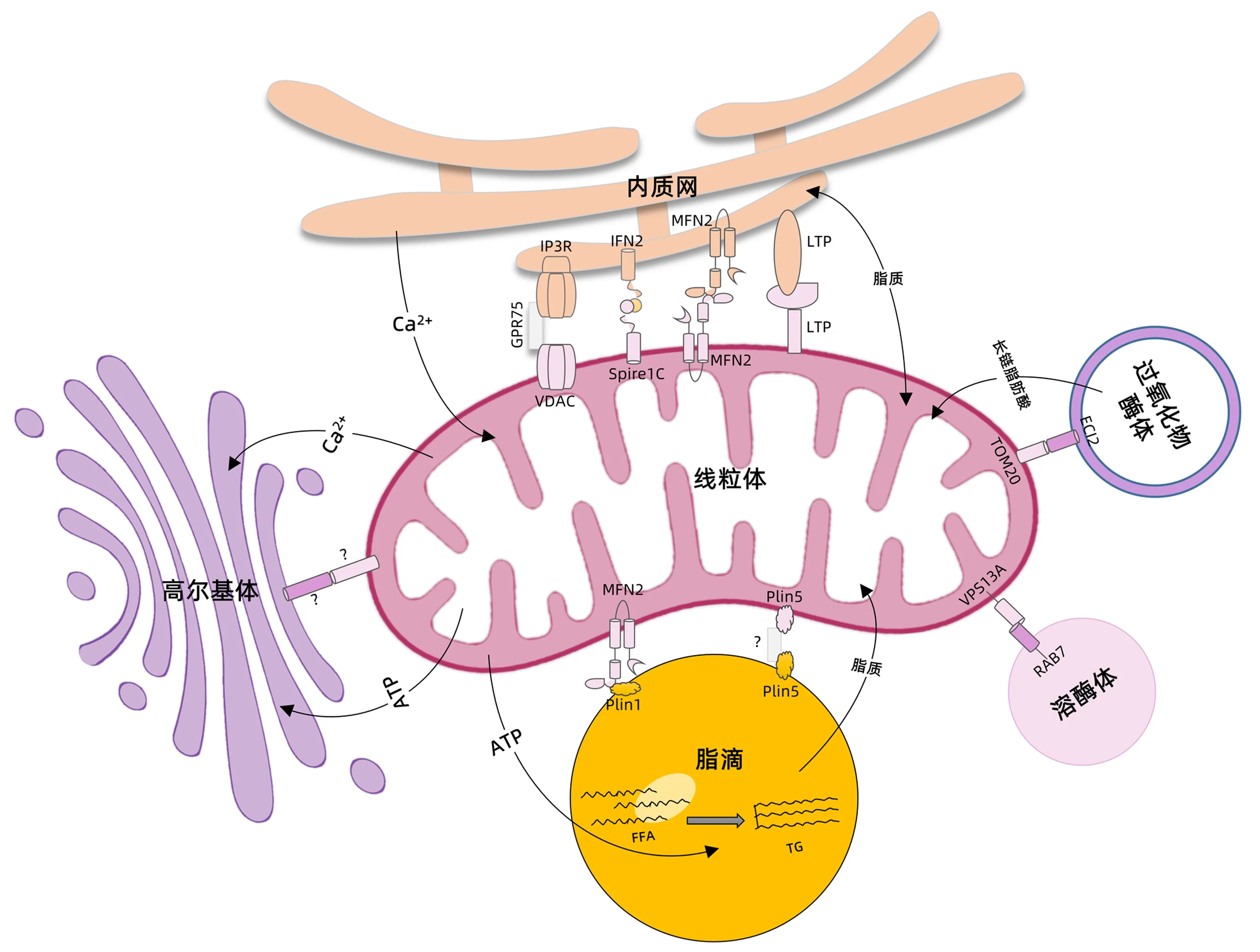
注:FFA,游离脂肪酸;IP3R,三磷酸肌醇受体;VDAC,电压依赖性阴离子选择性通道蛋白;GRP75,葡萄糖调节蛋白75;IFN2,肌动蛋白成核因子2;Spire1C,与甲酸结合的肌动蛋白成核蛋白1C;Mfn2,线粒体融合蛋白2;Plin1,脂滴包被蛋白1;Plin5,脂滴包被蛋白5;ECI2,烯酰辅酶A异构酶2;TOM20,线粒体外膜转位酶20;RAB7,Ras相关GTP结合蛋白7。图中使用的线粒体及高尔基体模式图来自BioRender网站。
2.1 线粒体和内质网之间相互作用与NAFLD 内质网是细胞中面积最大的膜性细胞器,与大多数细胞器均存在密切接触。其中,内质网与线粒体相互作用在NAFLD中的研究最为深入。内质网与线粒体形成多个接触位点,这些接触位点连在一起形成了线粒体-内质网相关膜(mitochondria-associated membranes,MAM),完整的MAM结构是内质网与线粒体之间钙离子、活性氧及脂质转移的基础[21-22]。
在NAFLD患者的肝脏中,MAM的完整性被破坏,表现为膜之间空间距离的改变及MAM中蛋白组成的变化。MAM中含有多种参与调控钙离子转移及糖脂代谢的蛋白,其完整性的破坏可导致钙离子稳态失衡,引起接下来的内质网应激和胰岛素抵抗,进而导致凋亡及炎症等病理改变,促进NAFLD的进展[21-22]。进一步的研究发现,Mfn2通过影响磷脂代谢调控MAM的完整性。在NASH患者肝组织中,Mfn2表达显著降低。与此相一致,在高脂饮食及胆碱-蛋氨酸缺乏饮食诱导的NASH小鼠肝组织中,Mfn2表达也显著降低。特异性敲除肝组织Mfn2基因,小鼠出现肝组织慢性炎症及脂质代谢异常,肝细胞凋亡和纤维化相关基因表达增加。通过尾静脉注射Mfn2重组腺病毒,使基因敲除小鼠重新表达Mfn2,小鼠脂质代谢异常得到恢复,纤维化及促炎基因的表达也显著降低[23]。一些对NAFLD有效的治疗药物,如罗格列酮及二甲双胍等有助于恢复MAM的完整性[22]。上述研究提示,调控内质网与线粒体之间的相互作用有可能成为治疗NAFLD的新策略。
2.2 线粒体和脂滴之间相互作用与NAFLD 棕色脂肪细胞中存在两群线粒体,一群与脂滴相互独立存在,被称为胞浆线粒体;另一群线粒体与脂滴密切接触,被称为脂周线粒体(peridroplet mitochondria,PDM)。进一步的研究[24-27]显示,PDM与脂滴以紧密锚定的方式稳定结合,用除垢剂或者胰蛋白酶处理后,两者之间的连接仍未受到破坏。与胞浆线粒体相比,PDM有较强的丙酮酸氧化和ATP合成能力,而脂肪酸β-氧化能力较弱。这些功能特点为甘油三酯(TG)合成提供了ATP,促进脂肪酸以TG的形式储存在脂滴中,同时减少了脂肪酸氧化[24]。
肝脏脂肪变性时线粒体功能的改变与棕色脂肪中PDM的功能特点有很强的相似性[28]。电镜下可以观察到高脂饮食的小鼠肝脏中线粒体与脂滴相互接触[29],提示PDM也存在于脂肪变的肝组织中。Plin5可以促进线粒体和脂滴相互接触,形成PDM[24,30],将脂肪酸以TG的形式储存于脂滴中,缓冲多余的FFA,保护肝细胞免于脂毒性[31]。Plin5可以通过促进脂滴与线粒体接触,上调线粒体功能相关基因,降低细胞活性氧水平,保护肝细胞免受氧化应激[32]。如果敲除Plin5,肝脏脂变减轻,但脂肪分解产生的FFA增加,引起脂毒性损伤[33]。另一方面,当胞浆中脂肪酸含量增加时,线粒体与脂滴接触位点的Plin5增加,介导脂肪酸直接从脂滴进入线粒体,促进脂肪酸的氧化利用,减少胞浆中高浓度脂肪酸产生的脂毒性[34-35]。上述研究提示,PDM在NAFLD的发病过程中可能发挥保护作用。
2.3 线粒体和其他细胞器之间相互作用与NAFLD 线粒体与其他细胞器之间也存在相互作用,参与调控钙离子传输、线粒体分裂及脂代谢等重要过程,提示其在NAFLD发生过程中可能也发挥相应的作用。线粒体与高尔基体之间的相互作用参与了钙离子及ATP向高尔基体传递的过程[20]。上文中提到,溶酶体可以通过与线粒体相互作用,参与线粒体的分裂过程[17]。最近的证据[36-37]还表明,线粒体通过膜上的外膜转位酶和过氧化物酶体上的ECI2与过氧化物酶体相互作用,调控类固醇生物合成。另外,过氧化物酶体还负责将长链脂肪酸运送至线粒体中完全氧化分解,提示线粒体与过氧化物酶体相互作用在脂代谢调控中发挥重要作用[20]。
3 脂滴和细胞器之间相互作用与NAFLD
脂滴是一种具有中性脂质核心的多功能膜性细胞器,可以与多种细胞器相互作用[7,38-39],发挥重要的代谢调控功能[40-41]。肝细胞中脂滴积聚引起的大泡脂变是NAFLD的主要病理学特征,脂滴与细胞器之间的相互作用成为NAFLD新的研究热点(图2)[42-43]。
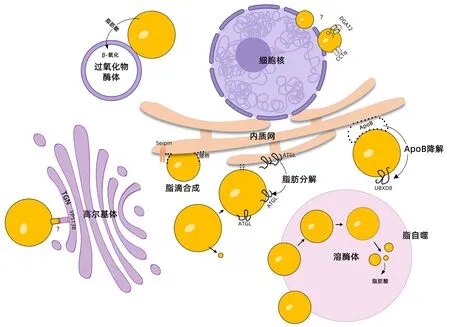
注:UBXD8,泛素调节X结构域蛋白8;TGN,高尔基体反面囊膜;VPS13B,液泡蛋白分选相关蛋白13B;DGAT2,二酰甘油酰基转移酶2;CCTα,磷酸胆碱胞苷酰转移酶α;ATGL,脂肪甘油三酯脂肪酶。图中使用的细胞核及高尔基体模式图来自BioRender网站。
3.1 脂滴和内质网之间相互作用与NAFLD 脂滴与内质网之间的相互作用存在两种方式:第一种称为膜桥,由连续的内质网膜与脂滴膜共同组成,常存在于脂滴生成的过程中;Seipin蛋白是促进膜桥稳定结合的关键蛋白,干预Seipin表达可导致内质网钙离子稳态失衡及脂质异常累积。第二种是经典的MCS,由分别定位于内质网膜及脂滴膜上的蛋白形成复合体连接而成,常发生于成熟的脂滴离开内质网后;如果干预复合体的形成,脂滴的扩增及TG的储存受到影响[7,44-45]。
肝脏中的脂肪来源于脂肪组织分解产生的FFA、脂肪酸从头合成及饮食摄入,而肝脏中合成的脂肪主要以极低密度脂蛋白(VLDL)的形式分泌,脂肪来源的增加及脂肪分泌障碍是引起NAFLD患者肝脏中脂质增多的主要原因[46]。脂滴与内质网相互作用在脂肪分解及VLDL合成分泌的过程中均发挥重要作用。成熟的脂滴会通过出芽的方式形成并释放出微小的脂滴,此时脂滴中磷脂比例降低,表面张力增加;这种增加的表面张力会诱导脂滴与内质网之间形成膜桥,并将ATGL从内质网募集到脂滴上,促进脂肪分解[44]。在肝细胞中,载脂蛋白B(ApoB)是组装分泌VLDL的关键分子;脂滴与内质网的相互作用通过调控ApoB降解影响VLDL的组装。ApoB在内质网腔侧结合脂质后在脂滴上积聚,接下来,ApoB从内质网的腔侧转移至胞质侧,被泛素化后与脂滴胞质侧的UBXD8结合,运送至蛋白酶体降解。在UBXD8基因缺陷小鼠中,血清VLDL水平降低,肝脏脂肪变性增加[44]。在高脂饮食喂养的NAFLD小鼠肝组织中,脂滴与内质网之间的相互接触增多,提示其可能影响脂肪的储存及输出过程[47]。
3.2 脂滴和高尔基体之间相互作用与NAFLD 脂滴与高尔基体相互作用可能参与了高尔基体分泌功能的调控。TGN是分选蛋白和脂质到细胞特定部位的关键结构,VPS13B可以介导TGN与脂滴之间形成膜接触位点,是维持TGN完整性和功能的关键蛋白,VPS13B功能异常会导致蛋白质分泌功能紊乱和脂质代谢失衡[7]。
Krahmer等[47]通过免疫荧光双染方法观察了高尔基体与脂滴之间的相互作用,在正常小鼠肝组织中,高尔基体特异性蛋白仅存在于少数较大的脂滴上;在高脂饮食喂养12周的NAFLD小鼠肝组织中,高尔基体特异性蛋白与脂滴包被蛋白Plin2存在广泛的共定位;进一步的体外研究也证实,在油酸处理的细胞中,高尔基体与脂滴的接触增加;上述结果提示,脂质负荷的增加促进高尔基体与脂滴的相互作用。在严重脂肪变性的小鼠肝组织中,高尔基体在细胞内的分布发生改变,大量高尔基体的膜结构碎片包裹在脂滴周围;分离脂变肝组织中的原代肝细胞,结果显示细胞的蛋白分泌能力显著降低,提示严重脂肪变性时高尔基体与脂滴相互接触增加,但TGN的完整性和功能可能受损[47]。
3.3 脂滴和其他细胞器之间相互作用与NAFLD 与细胞核相互接触的脂滴被称为核脂滴,核脂滴上含有TG和磷脂酰胆碱合成所需的DGAT2和CCTα,提示其可能参与脂代谢的调控。在产生内质网应激的肝细胞中,内核膜中核脂滴的数量显著增多,提示NAFLD发病过程中核脂滴的形成可能存在异常[7,43]。溶酶体与脂滴相互作用形成自噬小体,分解脂滴中的中性脂肪,提供FFA进入能量代谢,这一过程被称为脂自噬。在肝脏中,脂自噬是脂质分解的主要途径,而NAFLD患者肝组织中常常存在脂自噬的异常[48]。氧化应激和脂质过氧化与NAFLD的发生密切相关,过氧化物酶体中富含抗氧化酶和负责产生活性氧的酶,在脂肪酸β-氧化及氧化还原稳态维持中发挥重要作用。在油酸处理的细胞中,过氧化物酶体与脂滴形成稳定的膜接触位点,过氧化物酶体的内膜可延伸至脂滴中[7];对提取纯化的脂滴进行蛋白组分析,发现过氧化物酶体中的β-氧化酶在脂滴中富集[47]。上述结果提示,过氧化物酶体可以通过与脂滴相互作用影响氧化还原稳态。
4 结语与展望
综上所述,细胞器之间相互作用在细胞中广泛存在,通过离子、脂质及蛋白质等的交换,完成细胞的各种功能;细胞器互作网络的紊乱在钙离子稳态失衡、脂毒性、内质网应激、胰岛素抵抗、脂肪分解及输出等NAFLD相关病理过程中发挥重要作用。目前,细胞器之间相互作用在NAFLD发生发展中的作用还有许多未知的领域有待探索。以细胞器之间相互作用为切入点,进一步阐明NAFLD的发病机制,将为NAFLD药物研发提供新的方向。
利益冲突声明:作者声明不存在利益冲突。
猜你喜欢
中国预防兽医学报(2022年6期)2022-12-29
中国医学创新(2022年18期)2022-07-12
家畜生态学报(2022年2期)2022-02-28
科学24小时(2021年4期)2021-03-22
中国当代医药(2018年24期)2018-11-28
——可作为磷酸盐库再利用!
蔬菜(2018年12期)2018-01-16
中风与神经疾病杂志(2017年5期)2017-01-13
听力学及言语疾病杂志(2015年5期)2015-12-24
现代检验医学杂志(2014年3期)2014-02-02
——细胞脂类代谢的细胞器
Biophysics Reports(2010年2期)2010-09-13
