
胍基乙酸和甜菜碱对羔羊生长性能、瘤胃发酵和血液代谢的影响
2023-03-09 08:42任国栋郝小燕张暄梓刘森张宏祥田光元张建新
中国农业科学 2023年4期
任国栋,郝小燕,张暄梓,刘森,张宏祥,田光元,张建新✉
1山西农业大学动物科学学院,山西太谷 030801;2山西祥和岭上农牧开发股份有限公司,山西右玉 037200
0 引言
【研究意义】羊产业是我国畜牧业中重要的支柱产业,集约化舍饲养殖正逐渐成为养羊行业重要的生产模式。通过营养调控,提高肉羊饲料效率和生产性能,对肉羊产业“节本、提质、增效”发展具有重要意义。胍基乙酸(guanidineacetic acid,GAA)和甜菜碱(betaine,BT)作为两种功能性营养添加剂,现阶段研究主要集中在其单独使用效果,二者联合应用对动物生产性能影响尚不明确。【前人研究进展】胍基乙酸又称胍乙酸,天然存在于脊椎动物体内,主要在肝脏或胰腺中由精氨酸和甘氨酸生物合成,是肌酸的直接前体物质[1-2]。肌酸通过磷酸化产生磷酸肌酸,磷酸肌酸可为细胞活动提供能量,在机体能量代谢过程中发挥着重要作用[3-4]。WYSS等[5]报道肌酸主要存在于动物性副产品中,如鱼粉或骨粉,而植物性饲料缺乏肌酸,且动物源性副产品已被禁止用作反刍动物饲料[6]。此外,动物机体自身合成的肌酸仅占机体需要的50%—75%[7],难以满足需要。因此,需外源性补充肌酸。现如今,胍基乙酸作为稳定替代肌酸来源的营养补充剂[1],具有性质稳定、生物效价高等优点。研究表明,饲粮中添加胍基乙酸可以提高绵羊生长性能[8-9],促进安格斯公牛瘤胃发酵[10],改善育肥猪肉品质[11-12]。但有研究表明,外源性补充GAA可能会消耗机体 40%—75%的不稳定甲基供体,其甲基需要量甚至比其他甲基化反应总和还要多[13-15]。且 GAA在生成肌酸的同时会伴随同型半胱氨酸(homocysteine,Hcy)的生成,后者过度积累可引发“高同型半胱氨酸血症”,对机体健康造成危害。甜菜碱(betaine,BT),又名三甲基甘氨酸,作为良好的甲基供体,广泛存在于自然界植物中。饲粮中添加甜菜碱可以提高奶牛泌乳性能[16],促进公牛生长[17],且饲粮中添加过瘤胃甜菜碱可提高羔羊生长性能和抗氧化能力[18]。研究表明,甜菜碱甲基与同型半胱氨酸在甜菜碱同型半胱氨酸甲基转移酶的作用下转化为蛋氨酸和二甲基甘氨酸[19-20]。【本研究切入点】胍基乙酸作为合成肌酸的前体物质,可促进肌酸合成。但胍基乙酸在合成肌酸的同时会消耗机体大量的甲基供体,并伴随同型半胱氨酸的产生。而甜菜碱作为良好的三甲基供体,不但能为机体提供甲基,还参与同型半胱氨酸水解途径。鉴于胍基乙酸和甜菜碱在合成肌酸途径上的相互关系,本研究假设饲粮中添加胍基乙酸和甜菜碱能够促进肌酸合成,进而提高动物的生产性能。【拟解决的关键问题】在研究饲粮中添加胍基乙酸和甜菜碱对羔羊生长性能、瘤胃发酵及血清生化指标的影响,为胍基乙酸和甜菜碱在反刍动物生产中的应用提供理论依据。
1 材料与方法
1.1 试验设计
本试验于2020年7—10月在山西右玉祥和岭上养殖场进行。试验选用48只体重相近(22.03±1.30)kg、3月龄左右的杜泊×小尾寒羊杂交公羔,采用2×2因子完全随机试验设计,试验因素为 GAA(0或 900 mg·kg-1)和 BT(0 或 5 g·d-1)。随机分配到 4 组,每组 12只羊。分别为:(1)对照组,饲喂基础日粮;(2)GAA(纯度≥98%)组,饲喂基础日粮+900 mg·kg-1GAA(干物质基础);(3)BT(纯度≥78%)组,饲喂基础日粮+5 g·d-1BT(干物质基础);(4)GAA+BT组,饲喂基础日粮+900 mg·kg-1GAA 和 5 g·d-1BT(干物质基础)。试验期共75 d,其中预试期15 d,正试期 60 d。在正试期内,晨饲前用分析天平称取每日每只羊GAA和BT添加量,放入已装有60 g粉状饲粮的不锈钢碗里混合后,一次性饲喂给每只羊,以保证完全摄入。所有试验羊单栏饲养,每天各饲喂2次(08:00和17:00),自由采食和饮水,试验饲粮参照NRC(2007)绵羊营养需要量设计配方,基础饲粮组成及营养水平见表1。饲粮采用全混合颗粒饲料。

表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet (air dry basis, %)
1.2 样品采集
于正试期第 1天晨饲前对所有羊进行空腹称重,作为初始体重(IBW),之后试验第 30天进行第2次称重,记录数据。正试期结束当日晨饲前先采血后进行空腹称重,作为终末体重(FBW)。血清样品2 000×g离心15 min后分离血清,-80℃冷冻保存。饲喂期间记录每只羊的日采食量、剩料量。正试期结束后计算平均日增重(ADG)、平均日采食量(ADFI)和饲料效率(FE),其中饲料效率(FE)=平均日增重(ADG)/平均日采食量(ADFI)。
所有试验羊正试期结束当日正常饲喂,在试验羊采食后3 h通过口腔采集每只试验羊的瘤胃液。为了避免唾液污染,弃掉最初收集的瘤胃液,随后的80 mL瘤胃液立即测定其pH,同时将其分装在离心管中,一部分保存在液氮中用于分析瘤胃菌群定量和酶活,另一部分保存于-20℃测定瘤胃发酵指标。
1.3 指标测定
血清中乳酸、肌酸、肌酸激酶、L-精氨酸-甘氨酸脒基转移酶(AGAT)、S-腺苷蛋氨酸-胍基乙酸-N-甲基转移酶(GAMT)和同型半胱氨酸(Hcy)采用酶联免疫吸附(ELISA)法测定,ELISA试剂盒购自上海优选生物科技有限公司。
采集饲喂后3 h的瘤胃液用于测定挥发性脂肪酸(VFA)和氨态氮(NH3-N)浓度。参照严淑红等[22]方法利用气象色谱仪(Aglient 7980B,美国)测定VFA;参照金亚倩等[23]方法测定NH3-N浓度;参照AGARWAL等[24]方法测定瘤胃液中纤维素酶和淀粉酶;参照MILLER等[25]方法测定瘤胃液中蛋白酶。
采用珠磨-CTAB法[26]提取瘤胃微生物DNA。用核酸蛋白分光光度计(Thermo Scientific, 美国)测定提取的瘤胃微生物 DNA浓度和质量。由北京华大基因科技股份有限公司合成目的瘤胃微生物引物(表2)。目的微生物标品由山西农业大学动物营养实验室提供,采用10倍连续稀释法建立目的微生物标注曲线[27]。参照LI[10]等方法,使用TaKaRa 公司SYBR®Primic Ex TaqTM(TliRNaseHPlus)试剂盒及 ABI stepone plus 实时荧光定量聚合酶链式反应(PCR)仪器进行操作,对目的基因进行Real-time PCR。PCR反应体系如下(20.0 μL):2.0 μL DNA模板,10.0μL SYBR®PrimicExTaqTM(TliRNaseH Plus)(2x),6.0 μL dH2O(灭菌蒸馏水),0.8μL PCR Forward Primer(10 μmol·L-1),0.8 μL PCR Reverse Primer(10 μmol·L-1),0.4 μL ROX Reference Dye(50x)Ⅱ。

表2 基因引物序列Table 2 Primer sequences of genes
1.4 数据分析
试验所得数据通过Excel 2019进行初步整理,应用SAS9.2软件中一般线性模型(GLM)有交互作用的双因素过程进行统计分析,当交互作用显著时采用Tukey’s法进行多重比较。P<0.01表示差异极显著,P<0.05表示差异显著,0.5<P<0.10表示有趋势。
2 结果
2.1 饲粮中添加GAA和BT对羔羊生长性能的影响
生长性能数据已发表[33],详见表 3。饲粮中添加GAA或BT对羔羊终末体重无显著影响(P>0.05)。饲粮中添加 GAA有降低羔羊平均日采食量的趋势(P=0.072),且一定程度提高了 ADG,但差异不显著(P>0.05),添加 BT 则显著提高了 ADG(P<0.05)。添加GAA或BT显著提高FE(P<0.05)。在互作效应方面,GAA和BT对FE有显著互作效应(P<0.05),对ADG有极显著互作效应(P<0.01)。

表3 胍基乙酸和甜菜碱对羔羊生长性能的影响Table 3 Effects of GAA and BT on growth performance of lambs
2.2 饲粮中添加GAA和BT对羔羊瘤胃发酵的影响
由表4可知,饲粮中添加GAA或BT显著降低了瘤胃液 pH(P<0.05),对瘤胃液中乙酸、丁酸、异丁酸、乙丙比和 NH3-N均无显著影响(P>0.05)。添加BT显著提高了羔羊采食后3h的丙酸和总挥发性脂肪酸浓度(P<0.05),GAA则极显著提高了丙酸和总挥发性脂肪酸浓度(P<0.01)。添加BT极显著降低了戊酸和异戊酸浓度(P<0.01),而GAA显著提高了戊酸的浓度(P<0.05)。饲粮中添加GAA和BT对羔羊瘤胃发酵指标无显著互作效应(P>0.05)。

表4 胍基乙酸和甜菜碱对羔羊瘤胃发酵的影响Table 4 Effects of GAA and BT on ruminal fermentation of lambs
2.3 饲粮中添加GAA和BT对羔羊瘤胃微生物酶活性的影响
由表5可知,饲粮中添加GAA或BT显著提高了蛋白酶活性(P<0.05)。添加BT极显著提高了木聚糖酶、果胶酶和α-淀粉酶活性(P<0.01),显著提高了羧甲基纤维素酶活性(P<0.05),且有提高纤维二糖酶活性的趋势(P=0.075)。饲粮中添加GAA则有提高果胶酶和 α-淀粉酶活性的趋势(P=0.091,P=0.056)。饲粮中添加GAA和BT对果胶酶活性具有极显著互作效应(P<0.01)。
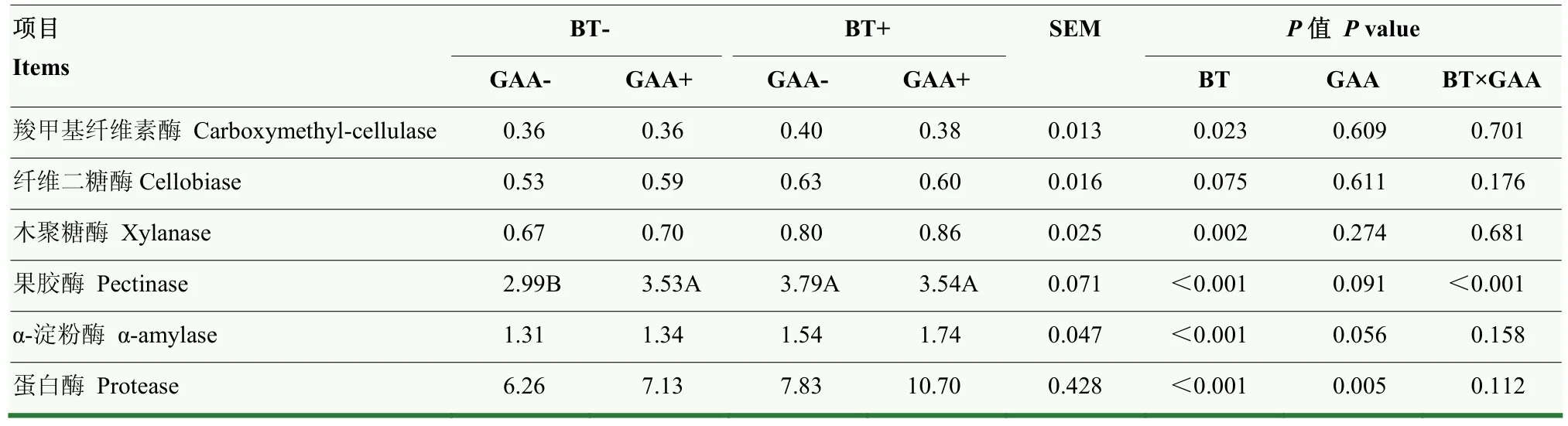
表5 胍基乙酸和甜菜碱对羔羊瘤胃微生物酶活性的影响Table 5 Effects of GAA and BT on rumen microbial enzyme activity of lambs (U·mL-1)
2.4 饲粮中添加GAA和BT对羔羊瘤胃功能微生物的影响
由表6可知,饲粮中添加GAA或BT极显著提高了羔羊瘤胃液中黄色瘤胃球菌和溶纤维丁酸弧菌的数量(P<0.01),显著提高了产琥珀酸丝状杆菌的数量(P<0.05),显著降低了瘤胃原虫的数量(P<0.05),对总细菌、总厌氧真菌、白色瘤胃球菌和嗜淀粉瘤胃杆菌则无显著影响(P>0.05)。添加BT则显著提高了栖瘤胃普雷沃氏菌的数量(P<0.05),显著降低了产甲烷菌的数量(P<0.05),且有提高嗜淀粉瘤胃杆菌数量的趋势(P=0.098)。添加GAA则显著提高了栖瘤胃普雷沃氏菌的数量(P<0.05),有降低瘤胃产甲烷菌数量的趋势(P=0.085)。添加GAA和BT对羔羊瘤胃微生物菌群无显著互作效应(P>0.05)。

表6 胍基乙酸和甜菜碱对羔羊瘤胃菌群的影响Table 6 Effects of GAA and BT on ruminal microflora of lambs (U·mL-1)
2.5 饲粮中添加GAA和BT对羔羊血清生化指标的影响
由表7可知,饲粮中添加GAA和BT对羔羊血清生化指标无显著互作效应(P>0.05)。添加GAA或BT显著提高血清中肌酸含量(P<0.05)。饲粮中添加 GAA有提高血清中同型半胱氨酸的趋势(P=0.053),有降低AGAT的趋势(P=0.056)。添加BT极显著降低血清中同型半胱氨酸浓度,且有提高肌酸激酶浓度的趋势(P=0.063)。
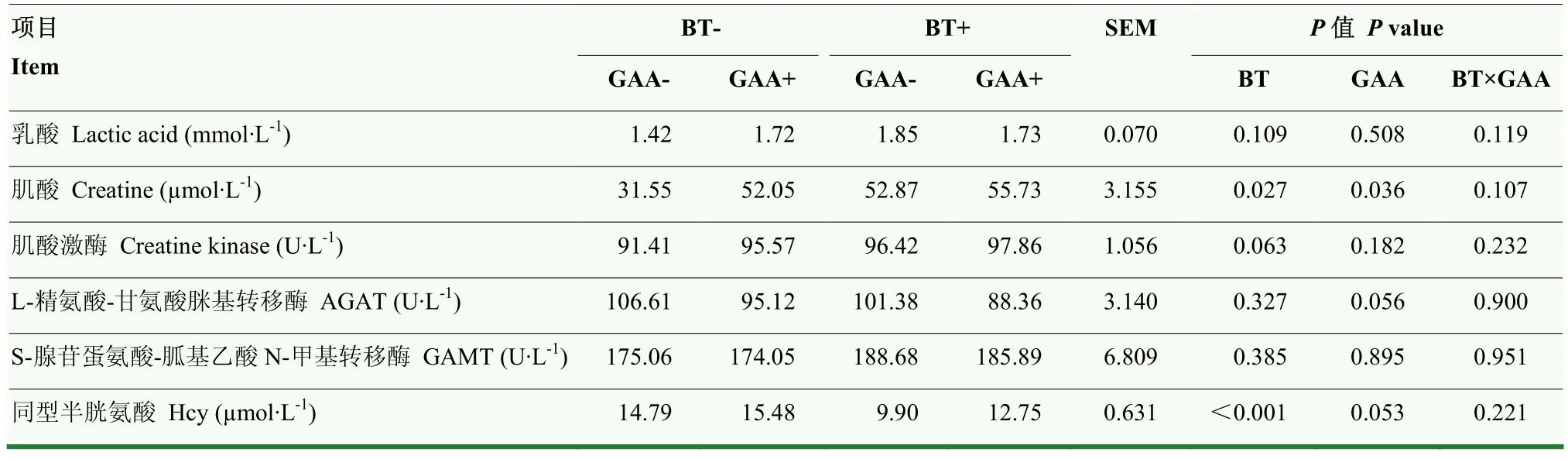
表7 胍基乙酸和甜菜碱对羔羊血清生化指标的影响Table 7 Effects of GAA and BT on serum biochemical parameters of lambs (U·mL-1)
3 讨论
3.1 饲粮中添加胍基乙酸和甜菜碱对羔羊生长性能有不同程度的影响
SPEER等[34]通过瘤胃与皱胃灌注 GAA研究发现,有50%左右的GAA可通过瘤胃进入肠道。这说明GAA在瘤胃中被部分降解。研究发现,在饲粮中添加未包被的GAA可提高绵羊和公牛的生产性能[8-10]。ARDALAN等[35]发现瘤胃后灌注GAA提高了瘘管牛血浆中的肌酸浓度,而包被胍基乙酸对反刍动物生产性能的效果还有待于进一步探究。甜菜碱具有较高的水溶性,进入瘤胃后大约有20%—45%被利用,而且在以粗饲料为主的日粮中代谢更快[36-37]。饲粮中添加BT可提高公牛的生长性能[17],增加奶牛的产奶量[16],饲粮中添加过瘤胃BT可减少羔羊腹部脂肪,但对生产性能无显著影响[18,38-39]。但也有研究表明饲粮中分别添加BT和过瘤胃BT均提高了羔羊的日增重,改善肉品质,且过瘤胃BT效果优于未包被BT[40]。造成研究结果差异的原因可能是因为BT添加水平、试验动物等因素。
采食量的高低影响反刍动物能量的摄入。前人研究表明饲粮中添加 GAA对羔羊平均日采食量无显著影响[8-9]。但也有研究发现,饲粮中添加GAA提高了安格斯公牛和育肥猪的平均日采食量[10-11]。本研究结果表明,饲粮中添加 GAA有降低羔羊平均日采食量的趋势,这与前人研究结果不一致。影响动物采食量的因素有很多,主要包括动物生长发育水平、日粮结构、环境和饲养管理等方面。研究发现,较高的NDF水平下,瘤胃的充盈程度会直接抑制采食[41]。辛均平等[42]研究发现添加 GAA可以促进体外瘤胃产气量及挥发性脂肪酸的增加。在本试验中,饲粮中添加GAA可能促进了瘤胃产气,使瘤胃充盈程度相对提高,这可能是导致平均日采食量有降低趋势的原因。LI等[10]研究发现,在精粗比5∶5全混合日粮中添加0.9g·kg-1GAA显著提高了营养物质消化率,改善了瘤胃发酵,提高了公牛的生长性能。此外,也有研究表明 GAA对 ADG无显著影响[43-45]。在本试验条件下,饲粮中添加GAA提高了FE,但对ADG影响有限。不同试验结果的差异可能是由于不同的 GAA添加比例及日粮配方组成导致。
本试验结果表明饲粮中添加BT对羔羊的平均日采食量无显著影响。CHENG等[16]发现饲粮中添加0.6 g·kg-1BT对奶牛平均日采食量无显著影响。同样地,LI等[18]研究发现饲粮中补充6 g·d-1过瘤胃BT对湖羊平均日采食量虽有提高,但无显著影响,与本试验结果一致。日粮在瘤胃中经微生物发酵作用产生的挥发性脂肪酸主要经瘤胃壁吸收后进入机体组织,进行氧化供能或用于合成脂肪或葡萄糖,而能量水平的高低又决定动物的生长速度。本试验中,饲粮中添加 BT显著提高了羔羊的日增重和饲料效率,这可能与饲粮中添加BT促进瘤胃发酵有关。
3.2 饲粮中添加胍基乙酸和甜菜碱对羔羊瘤胃发酵有影响
日粮经反刍动物采食后进入瘤胃,瘤胃中微生物会产生各种消化酶,将日粮中的碳水化合物分解并生成乙酸、丙酸、丁酸等挥发性脂肪酸[46],为反刍动物提供70%—80%的能量来源,并且VFA产量与能量转化效率呈正相关,与pH呈负相关[47]。瘤胃发酵产生的大量VFA会在短时间内导致瘤胃pH下降,随着机体对VFA的吸收及唾液中的大量缓冲盐流入瘤胃,瘤胃内容物的酸度得到很好的缓冲,使瘤胃 pH保持稳定[48]。LI等[10]研究发现,饲粮中添加GAA显著提高了丙酸及总挥发性脂肪酸的浓度,降低了瘤胃pH,这与本试验研究结果一致。日粮在瘤胃中降解发酵的程度与瘤胃微生物菌群数量密切相关,总挥发性脂肪酸的增加可能与瘤胃主要功能微生物菌群数量及微生物酶活性的提高有关。NH3-N是瘤胃微生物降解饲粮蛋白质的产物,也是合成微生物蛋白的主要前体物质,当其释放速率与瘤胃微生物可利用能量释放速率不同步时,就会造成瘤胃中NH3-N累积[49]。研究发现,在精粗比 5∶5的饲粮中添加 GAA显著降低了瘤胃NH3-N浓度[10],而LIU等[50]研究发现添加GAA显著提高了瘤胃 NH3-N浓度。在本试验中,饲粮中添加GAA显著增加了相关蛋白降解菌群的数量及蛋白酶活性,但对瘤胃NH3-N浓度无显著影响,这与刘笑梅等[8]研究结果一致,其原因可能是GAA的添加促进了微生物利用NH3-N合成微生物蛋白。
甜菜碱作为一种功能性添加剂,可以通过调节渗透压维持细胞正常形态与功能,还可以帮助胃肠道抵抗氧化应激改善其功能[51-52]。研究发现饲粮中添加甜菜碱可以提高干物质、粗蛋白等营养物质在公牛消化道中的消化率,并显著提高乙酸、丙酸和总挥发性脂肪酸的含量[17]。RATRIYANTO等[53]发现饲粮中添加BT显著提高了粗蛋白和大部分氨基酸在仔猪全消化道中的消化率。本试验结果表明,饲粮中添加BT显著提高了羔羊瘤胃中丙酸和总挥发性脂肪酸的浓度,这可能与瘤胃功能性微生物数量及酶活性的提高有关。CHENG等[16]研究表明,饲粮中添加4 g·kg-1BT显著降低了奶牛瘤胃NH3-N浓度,而WANG等[17]研究发现添加0.6 g·kg-1BT对安格斯公牛瘤胃NH3-N浓度无显著影响,这与本试验结果一致。导致不同结果的原因可能是不同的BT添加比例及饲粮配方组成。
3.3 饲粮中添加胍基乙酸和甜菜碱可提高羔羊瘤胃功能微生物数量及酶活性
研究表明添加 GAA显著提高了公牛瘤胃中白色瘤胃球菌、黄色瘤胃球菌、产琥珀丝状杆菌、溶纤维丁酸弧菌和嗜淀粉瘤胃杆菌的菌群数量,提高了瘤胃微生物纤维素酶、蛋白酶和α-淀粉酶的活性[50]。其中白色瘤胃球菌、黄色瘤胃球菌、产琥珀丝状杆菌和溶纤维丁酸弧菌是具有很强纤维降解能力的纤维分解菌,它们能够产生大量的纤维素酶。溶纤维丁酸弧菌和栖瘤胃普雷沃氏菌是主要的蛋白降解菌,能够分泌降解蛋白的酶。嗜淀粉瘤胃杆菌主要发酵淀粉,是瘤胃中主要淀粉降解菌之一。本试验中,饲粮中添加GAA显著提高了黄色瘤胃球菌、产琥珀丝状杆菌、溶纤维丁酸弧菌和栖瘤胃普雷沃氏菌菌群数量,提高了果胶酶和蛋白酶的活性,且有提高淀粉酶活性的趋势,这与前人研究结果一致。GAA作为肌酸的前体物质,可间接通过促进肌酸合成从而在细胞能量代谢过程中发挥重要作用[54],外源补充的GAA可能为瘤胃微生物的生长繁殖提供了额外的能量来源。此外,瘤胃中还栖息着一类重要的微生物瘤胃原虫,其种类和数量随反刍动物饲喂条件、生理等因素的变化而发生变动。因此,瘤胃原虫的这种变化通常被作为反映瘤胃环境状况的一个指标。韩昊奇等[55]研究发现随着精料比例的增加,瘤胃 pH显著降低,而瘤胃内原虫数量在精料比例最高时降至最低。这说明瘤胃原虫数量与反刍动物的日粮结构及瘤胃 pH密切相关。本试验结果表明,饲粮中添加 GAA显著降低了瘤胃原虫的数量,这与LI等[10]研究结果一致,瘤胃原虫数量的减少可能与瘤胃pH显著降低有关。但也有研究表明饲粮中添加0.6 g·kg-1GAA显著提高了瘤胃原虫的数量[43]。不同的研究表明 GAA对瘤胃原虫数量的影响有差异,其影响还需进一步深入研究。在瘤胃中产甲烷菌是一类重要的微生物,它能将CO2、H2、甲酸及其他化合物转化成甲烷或甲烷和CO2,从中获得能量。瘤胃产甲烷菌和原虫之间属于共生关系[56],且研究发现在瘤胃去除原虫后瘤胃甲烷生成量减少9%—40%[57]。瘤胃发酵趋于丙酸型发酵可竞争性抑制瘤胃甲烷产生,而低的瘤胃 pH也可抑制产甲烷菌[58-59]。本试验结果表明,饲粮中添加 GAA显著降低了瘤胃产甲烷菌的数量,这与瘤胃原虫数量减少及降低的瘤胃pH相一致。
甜菜碱作为良好的甲基供体,其甲基也可作为瘤胃微生物的直接底物[60]。研究发现饲粮中添加BT可以提高瘤胃原虫、白色瘤胃球菌、黄色瘤胃球菌、产琥珀酸丝杆菌和嗜淀粉瘤胃杆菌的数量,并提高蛋白酶和纤维素酶的活性[16-17]。在本试验中,添加 BT显著提高了黄色瘤胃球菌、产琥珀酸丝状杆菌、溶纤维丁酸弧菌和栖瘤胃普雷沃氏菌的数量,有提高嗜淀粉瘤胃杆菌数量的趋势,且提高了瘤胃中纤维素酶、淀粉酶和蛋白酶的活性,这与前人研究结果一致。瘤胃原虫和瘤胃产甲烷菌数量的降低可能与补充BT促进了挥发性脂肪酸的生成直接降低了瘤胃pH有关。
3.4 饲粮中添加胍基乙酸和甜菜碱对羔羊血清生化指标的影响
外源性肌酸前体物 GAA在肝脏中由合成肌酸并释放进入血液[2]。本试验结果表明,饲粮中添加GAA显著提高了血清中肌酸的浓度。LI等[10]研究发现饲粮中添加 GAA显著提高了公牛血清中的肌酸浓度。刘洋等[12]发现在给育肥猪补充GAA15d后,显著提高了育肥猪肌肉中的肌酸含量。这与本试验研究结果一致。血清中肌酸浓度的提高可能与 GAA促进肌酸转化有关。AGAT的作用主要是在肝脏或胰腺中促进精氨酸和甘氨酸合成 GAA,而 GAA的浓度可以反馈调节AGAT的生成[5]。在本试验中,饲料中添加GAA有降低血清中AGAT浓度的趋势,这可能与额外补充GAA有关。ARDALAN等[35]发现通过皱胃输入GAA显著提高了奶牛血清中肌酸含量,但同型半胱氨酸的含量也随之提高。本试验研究表明,饲粮中添加 GAA有提高血清中同型半胱氨酸的趋势。作为 GAA合成肌酸时的伴随产物,同型半胱氨酸的提高可能与 GAA促进肌酸合成有关。本研究表明,补充BT显著提高了血清中肌酸含量,并显著降低了同型半胱氨酸的含量,这可能是由于BT的添加促进了肌酸的合成,并与生成的同型半胱氨酸一同转化为蛋氨酸和二甲基甘氨酸。
在本试验中,饲粮中添加GAA和BT对ADG和饲料效率有显著的交互作用,但是GAA和BT的组合效果却对ADG的提高有一定的限制。高浓度的谷物和可发酵碳水化合物可以为反刍动物提供足够的能量,但在长期饲养过程中也会导致瘤胃和后肠道pH下降和VFA积累,导致微生物代谢紊乱,阻碍生长[61-62]。我们推测在日粮中添加GAA和BT可以协同促进瘤胃发酵,过量的挥发性脂肪酸在短时间内使瘤胃 pH急剧下降,使羔羊产生应激,对机体健康生长造成负面影响。此外,与单独添加GAA或BT相比,联合添加GAA和BT可提高血液中肌酸的浓度,这表明BT的添加能够促进GAA向肌酸的转化。GAA和BT都可以促进挥发性脂肪酸的产生,降低瘤胃pH,因此未来可以探究在精粗比5∶5饲粮配比下GAA和BT能否会有更好的促进作用。
4 结论
饲粮中添加胍基乙酸或甜菜碱可以提高血清中肌酸含量,增加瘤胃总挥发性脂肪酸产量和功能微生物菌群数量,提高瘤胃内微生物酶活性,进而提高饲料效率,且甜菜碱可以降低血清中同型半胱氨酸的浓度。但与单独添加胍基乙酸或甜菜碱相比,联合添加胍基乙酸和甜菜碱对羔羊日增重的影响有限,这可能与该研究中饲粮精料比例过高和过低的瘤胃 pH使羔羊在长期饲喂过程中产生了一定的应激有关。
猜你喜欢
中学生数理化·高一版(2022年4期)2022-05-09
中国畜牧业(2022年4期)2022-04-02
中国学校体育(2018年9期)2018-05-14
运动(2016年17期)2017-01-03
中国体育教练员(2016年4期)2016-11-29
浙江大学学报(工学版)(2016年9期)2016-06-05
当代化工研究(2016年5期)2016-03-20
中国洗涤用品工业(2015年8期)2015-02-28
无机化学学报(2014年9期)2014-02-28
江西理工大学学报(2013年1期)2013-03-20
