
连作与轮作谷子根际土壤真菌群落分布特征*
2023-05-21 02:56王根全郝晓芬郭二虎杨慧卿张艾英程乔林秦玉忠王军
中国生态农业学报(中英文) 2023年5期
王根全 ,郝晓芬 ,郭二虎,杨慧卿,张艾英,程乔林,秦玉忠,王军
(山西农业大学谷子研究所 长治 046011)
谷子(Setaria italica)有抗旱耐瘠特性,是我国古老的栽培作物之一,种植面积和总产量均居世界首位[1-2]。随着全球性干旱日趋严重,旱作农业越来越受到重视[3],加之谷子是全谷物食物[4],营养丰富、药食同源,对调节我国人民膳食结构起着重要作用,为谷子产业发展带来了新的机遇。由于土地资源稀缺,谷农为追求经济效益最大化,谷子连作时有发生,由此引起的连作障碍难题也已成为制约谷子可持续发展的关键问题。
连作是导致作物产量降低、病害加重、品质下降的重要原因。不同作物连作障碍程度不同,有的作物连作障碍较轻,如小麦(Triticum aestivum)、玉米(Zea mays)和水稻(Oryza sative)等,但大部分作物,如谷子、棉花(Gossypium hirsutum)、黄瓜(Cucumis sativus)、草莓(Fragaria×ananassa)、苦参(Sophora flavescens)、黄连(Coptis chinensis)和马铃薯(Solanum tuberosum)等连作障碍严重[5-11]。造成连作障碍的原因主要有4 个方面:第一,土壤微生态失衡,有害微生物富积,有益微生物种群退化[12-13];第二,作物偏好的营养元素缺失,有害元素富集,导致土壤理化性质发生改变[14];第三,作物根际分泌物累积,产生自毒作用[15];第四,连续使用相同的肥料和农药,使得土壤出现酸化、板结等土壤退化现象,病虫害的抗性不断增强[16-17]。其中,土壤微生态失衡是连作障碍发生的根本原因[18]。
土壤微生物是植物-土壤生态系统重要组成,细菌和真菌等微生物在植物生长过程中发挥重要作用。土壤真菌包含有益真菌和病原真菌,有益真菌可促进作物对营养元素的摄取,相比细菌而言,能更好地降解复杂化合物[19-21];病原菌可入侵植物的根、茎、叶等组织器官,并造成危害,从而引起农作物减产或者品质下降[22-23]。谷子不同施肥条件、不同轮作方式对土壤理化性质、微生物多样性的影响已广泛开展研究[24-26]。Debenport等[27]研究表明,谷子与灌木间作,根际土壤微生物种群与谷子生长显著相关;Dang等[28]研究表明谷子与绿豆(Vigna radiata)间作改变了根际土壤微生物多样性;牛倩云等[29]研究表明谷子轮作可促进土壤细菌多样性;孙倩等[30]指出谷子与大豆(Glycine max)轮作模式下,谷子根际微生物环境最佳。
本研究以撂荒地和连作、轮作谷子根际土壤为研究对象,采用Illumina MiSeq 高通量测序技术[31-32],依据ITS 基因测序结果,结合土壤理化性质测定结果,分析了不同种植模式下谷子根际土壤真菌群落结构和多样性变化,旨在了解谷子根际土壤真菌群落、土壤理化性质与谷子连作的关系,为克服谷子连作障碍提供理论依据,促进有机旱作农业的可持续发展。
1 材料与方法
1.1 试验地概况与试验设计
试验在山西农业大学谷子研究所科研基地(以下简称科研基地)进行。科研基地位于36°11′N、113°06′E,海拔930 m。试验从2016 年开始,前茬作物为玉米。试验设4 组处理,每组处理面积为90 m2,不设重复。4 组处理分别为撂荒地(CK,2016 年开始未种植任何作物)作对照、轮作(CR,2016 年种植谷子,谷子-玉米-谷子周年单序轮作) (CR)、谷子连作3 年(TC,2016-2017 年连续2 年种植玉米,2018-2020 年连续3 年种植谷子)、谷子连作5 年(FC,2016-2020 年连续5 年种植谷子),试验田施复合肥(N∶P∶K=10∶5∶5) 1200 kg·hm-2和有机肥7500 kg·hm-2,撂荒地不施肥,其他田间管理措施相同。
1.2 样品采集
2020 年于谷子成熟期,每组处理平分为3 个采样区,采用5 点取样法对各采样区取样。每采样区取5 株长势一致的谷子,将根周5 cm 的根系全部挖出,去除表面附着土,轻抖根系,用软毛刷轻刷根上的根际土,各采样区的5 个样品混匀为1 份,分别置入无菌自封袋(连同样本编号),放入冰盒,带回实验室。在实验室,将各样品分为两部分保存,一部分装入离心管,置于-80 ℃超低温冰箱,用作微生物多样性研究;另一部分置于实验室,风干后置入无菌自封袋,用于土壤理化性质测定。
1.3 土壤真菌总DNA 提取与纯化
采用E.Z.N.A Soil DNA 试剂盒(OMEGA 公司)提取土壤样品DNA,采用Nanodrop 对DNA 进行定量,采用1.2%琼脂糖凝胶电泳检测DNA 质量。采用BECKMAN AMPure XP Beads 再次纯化文库富集产物,采用 2%琼脂糖凝胶电泳,对文库做最终片段选择与纯化。
1.4 真菌ITS1 区PCR 扩增与测序
采用特异性引物ITS5F:5′-GGAAGTAAAAGTCGTAACAAGG-3′,ITS1R:5′-GCTGCGTTCTTCATCGATGC-3′对真菌ITS r RNA 的ITS-V1 区进行PCR 扩增。PCR 产物采用 2%琼脂糖凝胶电泳检测,用Axy Prep DNA 凝胶回收试剂盒回收目的片段。采用Illumina MiSeq 高通量测序平台进行样品测序。
1.5 序列获取与分析
序列获取:采用Illumina 平台对群落DNA 片段进行双端测序,采用DADA2 方法[33],去引物、质量过滤、去噪、拼接和去嵌合体等,得到真菌ITS 序列(ASV,amplicon sequence variants)。对真菌ITS 比对的数据库为UNITE-8 数据库。采用Qiime 2 的classify-sklearn 算法,用预先训练好的Naive Bays 分类器进行物种注释,并通过稀疏曲线反映测序深度是否合适。
序列分析:使用ASV 丰度表制作韦恩图,统计各组独有的及组间共有的ASV 个数。依据序列物种分类学注释的结果,统计各组在门、纲、目、科、属、种6 个分类水平各自含有的分类单元数量。对非单丰度序列的特征表进行统计,以柱状图呈现各组在门、纲、目、科、属5 个分类水平上的组成分布。以Good’s coverage 指数表征覆盖度,以Chao1[34]和Observed species 指数表征丰富度,以Shannon 指数表征多样性,以丰度等级曲线来反映多样性差异。基于 Bray-curtis 距离矩阵,采用主坐标分析(Principal coordinates analysis,PCoA)[35]和通过python 的scikitbio 包进行“permanova”组间差异分析来检验真菌群落的结构。利用在线软件(http://www.genescloud.cn)对土壤真菌属水平物种相对丰度与土壤理化性质的相关性进行冗余分析(RDA)。LEfSe 分析设置LDA阈值为2,筛查标志物种(biomarker)。
2 结果与分析
2.1 不同种植模式下真菌ITS1 区测序结果
为探索不同种植模式下谷子根际土壤真菌群落结构,对土壤样品进行菌群DNA 抽提,通过Illumina Miseq 高通量测序技术对真菌ITS1 区进行测序分析(表1)。从4 组处理12 个样品中共获得1 160 414 条优化序列,每个样品有256 268~357 026 条优化序列,平均290 103 条优化序列;优化序列长度为134~438 bp,平均长度227 bp。4 组处理的稀疏曲线均趋于平坦,说明各样品测序数据量合理,测序深度足够(图1),适合根际土壤真菌丰度和群落结构的进一步分析。
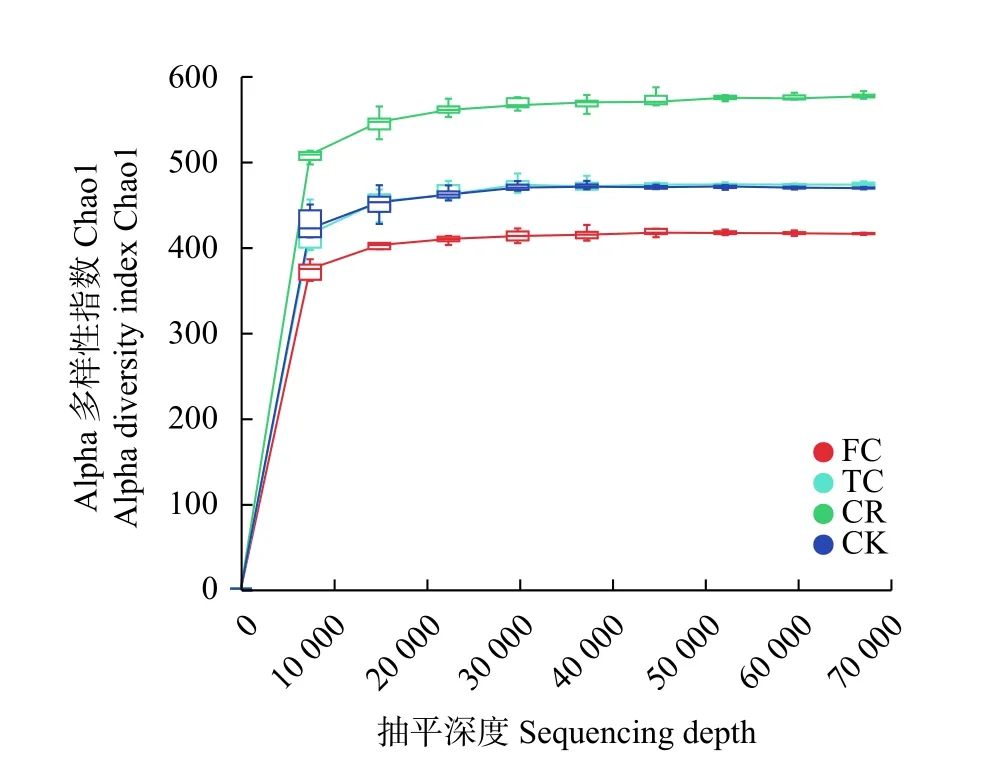
图1 不同种植模式谷子根际土壤真菌群落的稀疏曲线Fig.1 Sparse curves of soil fungal community of foxtail millet rhizosphere soils with different cropping patterns
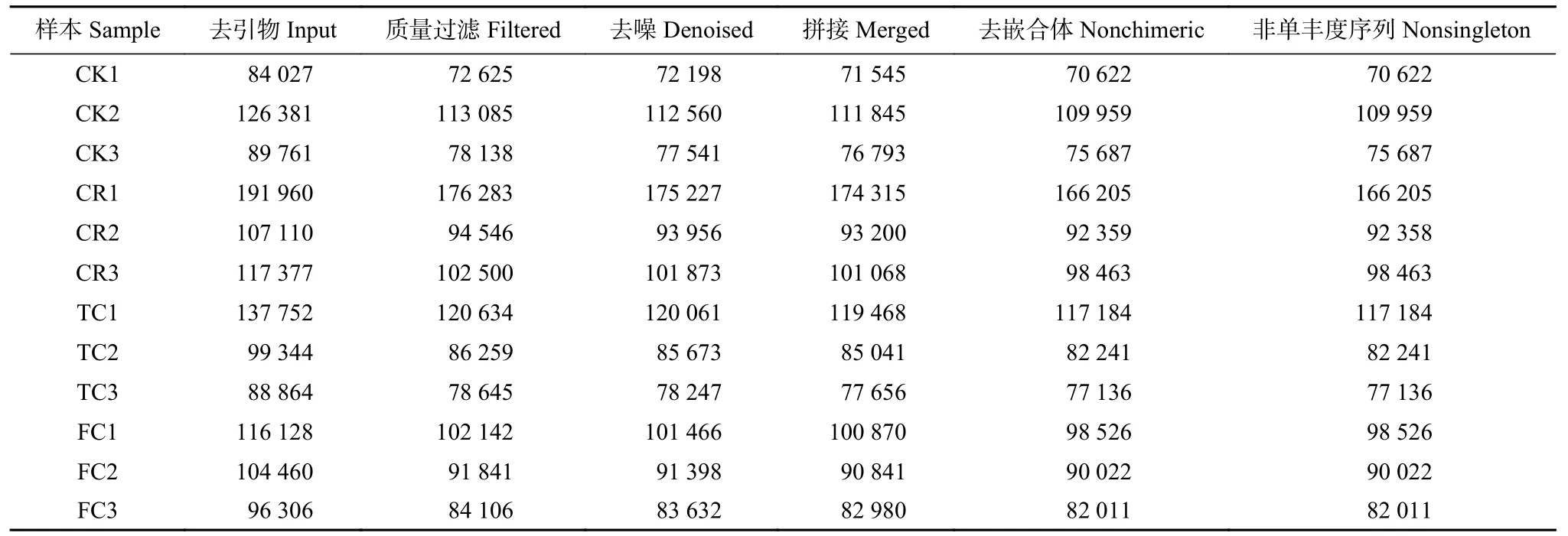
表1 不同种植模式谷子根际土壤的真菌高通量测序结果Table 1 High-throughput sequencing results of fungi in foxtail millet rhizosphere soils with different cropping patterns
2.2 不同种植模式下谷子根际土壤真菌群落结构变化
对测序结果进行聚类,共得到2637 个用于物种分类的ASV,其中CK、CR、TC 和FC 的ASV 分别为1014、1209、929 和831 个,4 组处理共有的ASV为172 个(图2A)。以门、纲、目、科、属和种6 个分类水平对各组处理ASV 进行划分,共检测到10 门、24 纲、46 目、79 科、136 属和149 种,各组处理在门、纲、目、科、属和种分类水平下微生物类群均有差异。与撂荒地(CK)相比,其他种植模式下谷子根际土壤(CR、TC 和FC)真菌群落更丰富,其中,轮作土壤(CR)真菌群落最丰富(图2B)。分类等级树图(图2C)表明,丰度前100 的ASV 主要分布在子囊菌门(Ascomycota)和担子菌门(Basidiomycota),其中以子囊菌门中的ASV 最多,丰度最高的ASV 也在子囊菌门中。
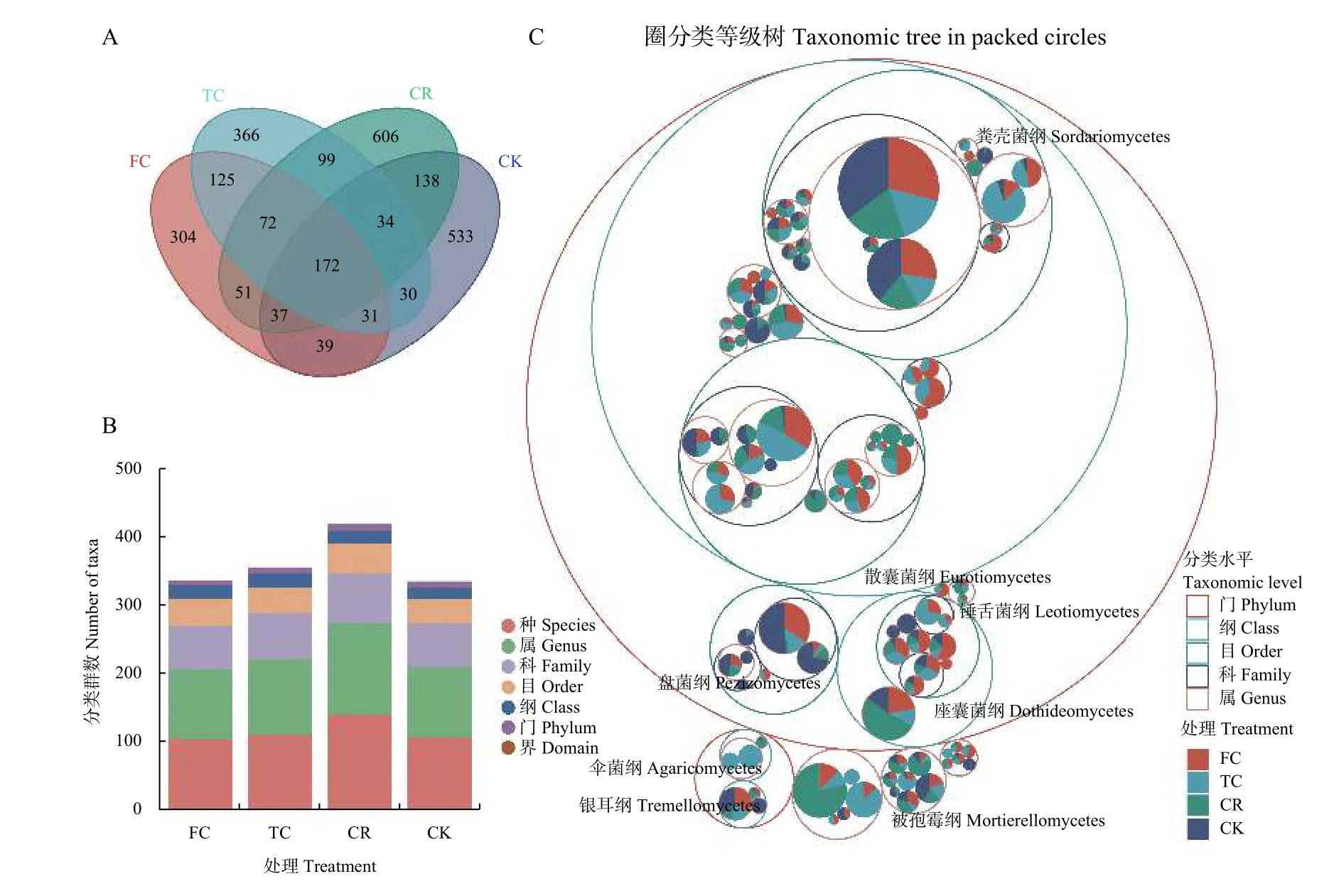
图2 不同种植模式谷子根际土壤真菌ITS 序列(ASV)分类Fig.2 Amplicon sequence variants (ASV) classification of fungi of foxtail millet rhizosphere soil in different planting patterns
在门、纲、目、科、属水平,对4 组处理土壤真菌群落的组成和丰度进行分析。门分类水平上,相对丰度前3 的有子囊菌门(70.48%~83.56%)、担子菌门(3.27%~7.27%)和被孢霉门(Mortierellomycota,3.41%~6.37%),其他门如壶菌门(Chytridiomycota)、球囊菌门(Glomeromycota)、油壶菌门(Olpidiomycota)、捕虫霉门(Zoopagomycota)、毛微门(Mucoromycota)、梳霉门(Kickxrllomycota)和罗兹菌门(Rozellomycota)相对丰度占比较少。轮作土壤(CR)中被孢霉门相对丰度高于连作土壤(TC 和FC),子囊菌门相对丰度低于TC 和FC;另外,在TC 中并未发现梳霉门、罗兹菌门和油壶菌门,FC 中未发现梳霉门(图3A)。
纲分类水平上,相对丰度前10 的有粪壳菌纲(Sordariomycetes)、座囊菌纲(Dothideomycetes)、盘菌纲(Pezizomycetes)、被孢霉纲(Mortierellomycetes)、伞菌纲(Agaricomycetes)、银耳纲(Tremellomycetes)、锤舌菌纲(Leotiomycetes)、散囊菌纲(Eurotiomycetes)、黑粉菌纲(Ustilaginomycetes)和裂壶菌纲(Spizellomycetes),其中相对丰度较高的有粪壳菌纲(53.60%~62.14%)、座囊菌纲(9.94%~13.05%)和盘菌纲(2.1%~15.98%)。与CK 相比,谷子根际土壤中盘菌纲相对丰度降低10%以上,CR 中的被孢霉纲和伞菌纲相对丰度高于TC 和FC,粪壳菌纲相对丰度低于TC 和FC (图3B)。
目分类水平上,大多数真菌属于肉座菌目(Hypocreales)、粪壳菌目(Sordariales)、格孢腔菌目(Pleosporales)、盘菌目(Pezizales)、被孢霉目(Mortierellales)以及煤炱目(Capnodiales)。与CK 相比,谷子根际土壤中的粪壳菌目相对丰度增加8%以上,盘菌目相对丰度降低10%以上。CR 中肉座菌目、格孢腔菌目相对丰度低于TC、FC,被孢霉目、煤炱目相对丰度高于TC 和FC (图3C)。
科分类水平上,相对丰度前10 的有赤壳科(Nectriaceae)、毛壳菌科(Chaetomiaceae)、毛球壳科(Lasiosphaeriaceae)、生赤壳科(Bionectriaceae)、火丝菌科(Pyronemataceae)、被孢霉科(Mortierellaceae)、球腔菌科(Mycosphaerellaceae)、囊菌科(Microascaceae)、荚孢腔菌科(Sporormiaceae)和Myrmecridiaceae。与CK 相比,谷子根际土壤中的赤壳科和火丝菌科相对丰度降低,而毛球壳科相对丰度增加。CR 中毛壳菌科相对丰度低于TC 和FC,被孢霉科、球腔菌科相对丰度高于TC 和FC (图3D)。
在属水平上,对相对丰度前20 的真菌进行了分析。其中与CK 相比,谷子根际土壤中的毛葡孢属(Botryotrichum)增加,TC 中含量最高;CR 中的被孢霉属(Mortierella)和球腔菌属(Mycosphaerella)相对丰度高于TC 和FC,链格孢属(Alternaria)和亚隔孢壳属(Didymella)相对丰度低于TC 和FC;粉红螺旋聚孢霉(Clonostachys)在TC 中最高(9.95%),在FC中较高(3.54%),CK 和CR 中较少,含量分别为0.74%和0.09% (图3E)。

图3 不同种植模式谷子根际土壤真菌在不同分类水平的相对丰度Fig.3 Relative abundance of fungi in foxtail millet rhizosphere soil under different cropping patterns at different classification levels
2.3 不同种植模式下谷子根际土壤真菌群落多样性
2.3.1 α 多样性分析
为了全面评估真菌群落的Alpha 多样性,分别以Chao1、Observed species、Shannon 以及Good’s coverage 等指数(图4A)和丰度等级曲线(图4B)来比较不同组间的多样性差异。4 组处理中的Good’s coverage 数值均在99%以上,证明处理中序列没有被测出的概率很低;Chao1 和Observed species 指数由高到低依次为CR>TC>CK>FC,且差异显著(P<0.05),说明不同种植年限谷子根际土壤真菌群落的丰富度差别较大;Shannon 指数由高到低依次为TC>CR>CK>FC,4 组样品的多样性差别未达显著水平;丰度等级曲线(图4B)表明,CR 具有更高的丰富度和均匀度,TC 的丰富度差异最大且均匀度较低。Alpha 多样性分析表明,谷子连作会改变根际土壤真菌群落丰富度和多样性,其中,CR 的丰富度最高,TC 的多样性最高,均匀度较低。

图4 不同种植模式谷子根际土壤真菌群落的Alpha 多样性分析Fig.4 Alpha diversity analysis of fungi community of foxtail millet rhizosphere soil under different cropping patterns
2.3.2 β 多样性分析
通过主坐标分析(图5A),观察到PCo1 解释了数据集中总方差的18.1%,而PCo2 解释了剩余变化的11.7%。不同处理组的生物学重复各自聚在一起,4组处理均有差异。与CK 和CR 相比,TC 和FC 菌群结构有很高的相似性。说明谷子连作改变了土壤真菌在属水平上的结构分布,撂荒地土壤与其他种植模式下谷子根际土壤之间的差别较大,轮作土壤与连作土壤之间差别也较大,连作土壤真菌结构较为相似。组间差异分析(图5B)表明,CK 组内3 个采样点的差异距离最小,CR 与CK,TC 与CK,FC 与CK 的组间差异距离均大于CK 组内差异距离,进一步验证了组间处理效应大于取样偏差。这些结果表明谷子在连作后根际土壤间真菌群落物种发生了改变,同时群落组成受种植年限的影响较大,受采样点差异的影响较小。
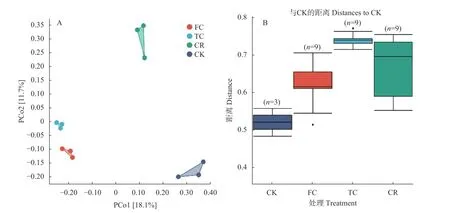
图5 不同种植模式谷子根际土壤真菌群落β 多样性分析Fig.5 β diversity analysis of fungi community of foxtail millet rhizosphere soil under different cropping patterns
2.4 不同种植模式下各指标相关性分析
2.4.1 土壤酶活性与理化性质的相关性分析
土壤理化性质测定结果见之前研究结果[36]。由表2 可知,土壤酶活性与理化性质存在相关性。其中,理化性质之间的相关性为:碱解氮与有机质呈极显著正相关(P<0.01),与有效磷呈显著正相关(P<0.05),有机质与有效磷呈显著正相关(P<0.05);酶活性之间均存在显著正相关性,其中多酚氧化酶与过氧化氢酶、蔗糖酶呈极显著正相关(P<0.01),脲酶和过氧化氢酶也存在极显著正相关(P<0.01);酶活性与理化性质间的相关性为:蔗糖酶活性与pH 呈显著负相关(P<0.05),多酚氧化酶活性与速效钾呈显著正相关(P<0.05),脲酶活性与碱解氮、有机质呈显著正相关(P<0.05)。
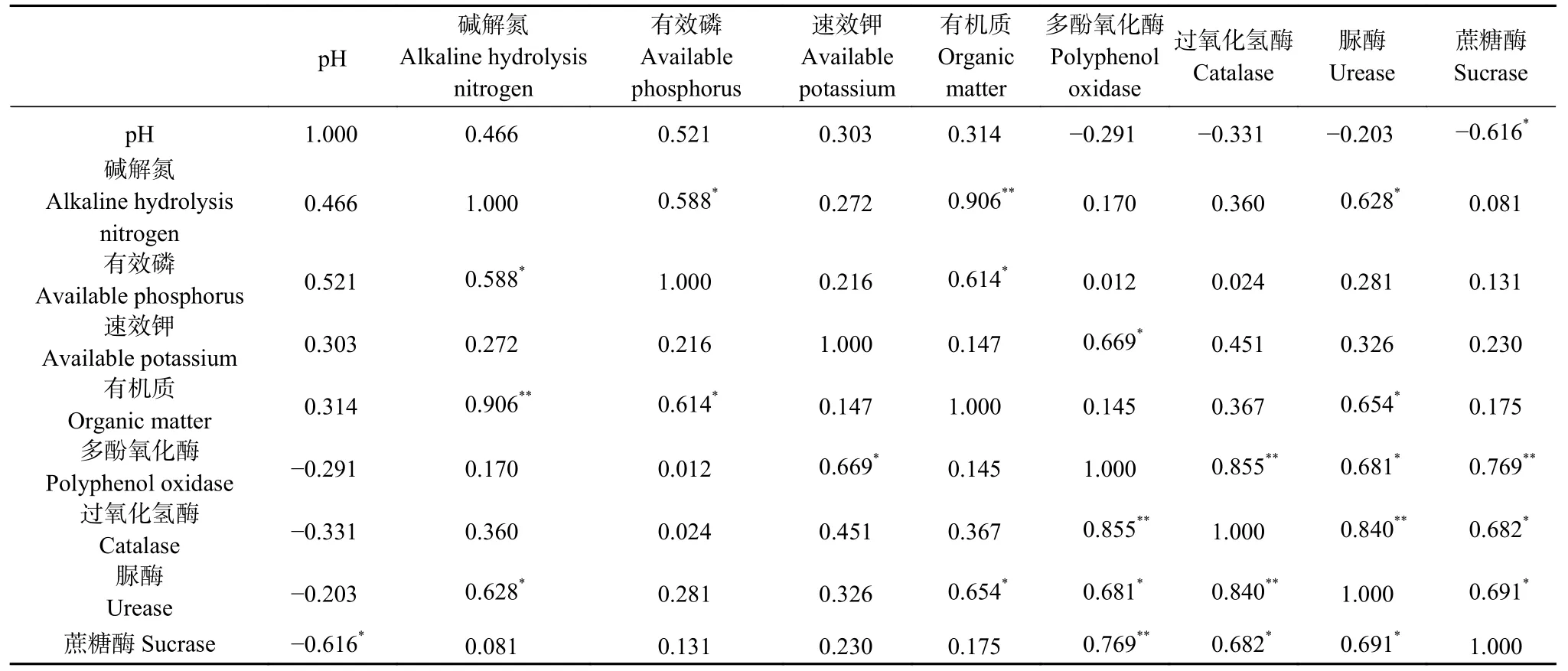
表2 谷子根际土壤酶活性与理化性质的相关性分析Table 2 Correlation analysis between enzyme activity and physicochemical properties of foxtail millet rhizosphere soil
2.4.2 土壤真菌群落多样性指数与理化性质、酶活性的相关性分析
表3 为不同种植模式下,谷子根际土壤真菌群落多样性指数与理化性质的相关性,由表可知,土壤理化性质、酶活性对真菌群落多样性指数的影响不同。真菌群落多样性指数与土壤理化性质存在相关性,但均未达显著水平。Chao1 指数、Observed species指数与多酚氧化酶活性呈极显著正相关(P<0.01),与过氧化氢酶、蔗糖酶活性呈显著正相关(P<0.05),故Chao1 指数、Observed species 指数受土壤酶活性影响较大。
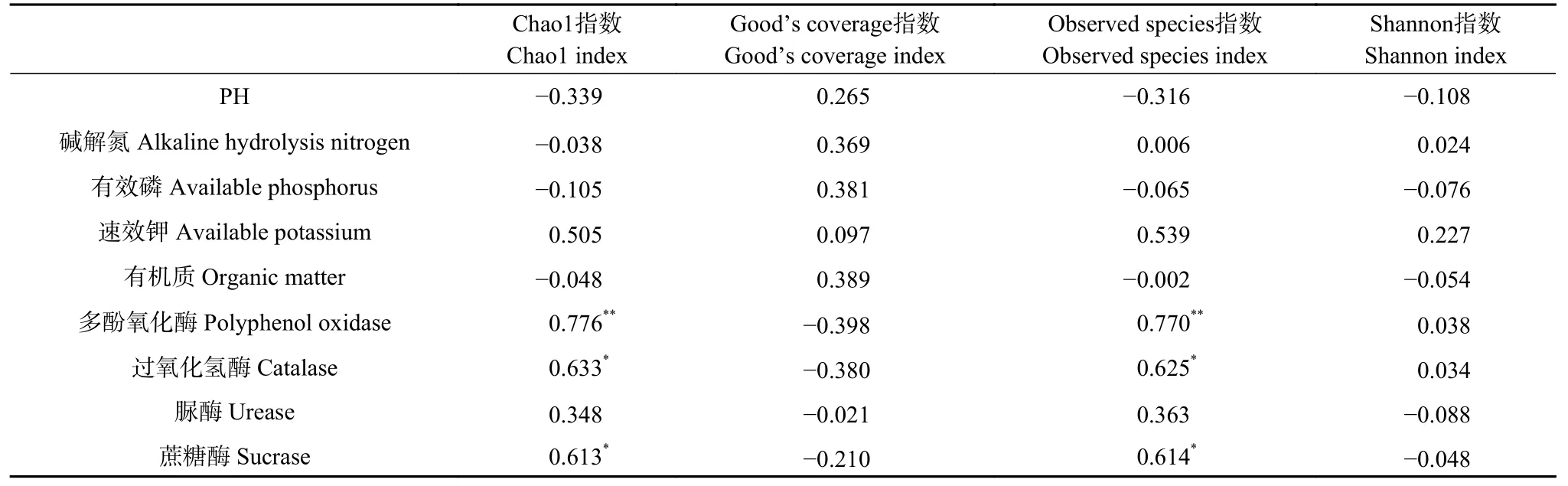
表3 谷子根际土壤真菌群落多样性指数与理化性质、酶活性的相关性分析Table 3 Correlation analysis of fungi community diversity indexes with physicochemical properties and enzyme activities of foxtail millet rhizosphere soil
2.4.3 土壤真菌(属水平)群落结构与理化性质、酶活性的相关性分析
将CK、CR、TC、FC 共4 组处理的土壤理化性质、酶活性与属水平真菌群落进行冗余分析(图6),结果显示,RDA1 和RDA2 的特征值分别为30.1%和20.74%,碱解氮、速效钾、有机质、4 种酶与RDA1 轴呈正相关;pH、有效磷与RDA1 轴呈负相关。CK 受毛壳菌影响较大,毛壳菌与所有理化性质和酶活性的相关性均不大。CR 受球腔菌属和微结节霉属的影响较大,与pH、有效磷、有机质、碱解氮、脲酶、过氧化氢酶呈正相关;TC 和FC 受毛葡孢属、毛喙壳属、亚隔孢壳属等影响较大,与有机质、碱解氮、脲酶、过氧化氢酶、多酚氧化酶、蔗糖酶、速效钾呈正相关。

图6 不同种植模式谷子根际土壤真菌群落冗余分析Fig.6 Redundancy analysis of fungi community of foxtail millet rhizosphere soil under different cropping patterns
2.5 不同种植模式下谷子田间生物标志物不同
LEfSe 分析(图7)设定LDA 阈值为2,确定了CK、CR、TC 和FC 根际土壤的22、19、8 和6 个生物标志物。其中CK 的生物标志物包含被孢霉属;CR 的生物标志物主要在担子菌门,包含被孢霉属和球腔菌属;TC 和FC 的生物标志物主要在子囊菌门,TC 的生物标志物包含毛葡孢属、亚格孢壳属和粉红螺旋聚孢霉属,FC 的生物标志物包含链格孢菌属和亚隔孢壳属。说明4 组处理的生物标志物不同。
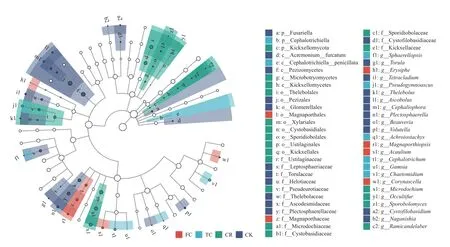
图7 不同种植模式谷子根际土壤真菌群落LEfSe 分析Fig.7 LEfSe analysis of fungi community of foxtail millet rhizosphere soil under different cropping patterns
3 讨论与结论
3.1 讨论
活跃的根际微生物作为植物的第2 套基因组,在植物的生长发育过程中发挥重要作用[37-40],土壤微生物组成及丰富度与谷子的连作障碍关系密切,长期连作会导致病害增加,尤其是黑斑病、叶枯病等真菌型病害[41]。
本研究发现谷子轮作土壤真菌群落多样性高于连作。在本研究中共得到2637 个用于物种分类的ASV,其中CR 土壤中的ASV 最多,稀疏曲线表明在相同测序深度下,CR 土壤真菌的多样性最高,α 多样性分析也表明CR 土壤真菌群落丰富度高于TC 和FC。这与保丽美等[42]在三七(Panax notoginseng)和李锐等[5]在棉花(Goysspiumspp.)上的研究结果类似,但有些研究认为随连作年限增加,土壤真菌种类和数量越多[43-44]。分析其原因,酸性环境有利于真菌生长,而本研究中连作并未导致谷子根际土壤pH 显著下降,4 组处理的土壤pH 均为碱性[36],这是因为2019-2020 年山西长治气候干旱,试验田进行了数次人工灌溉(地下水偏碱性),导致各处理土壤仅表现出轻微酸化,没有出现随连作年限延长,土壤真菌增多的现象。
通过对谷子不同种植模式下的土壤微生物进行高通量测序,发现4 组处理中,在门、纲、目、科、属和种6 个分类水平下微生物类群均有差异,不同种类真菌的相对丰度发生了变化。在门水平上,子囊菌门和担子菌门为谷子根际土壤的优势菌门,其中子囊菌门为谷子根际土壤最丰富的门,主要作用是分解有机质,这与孙倩等[45]对谷子根际土壤真菌研究结果相一致。在纲水平上,粪壳菌纲、座囊菌纲和盘菌纲为谷子根际土壤优势菌纲,其中,粪壳菌纲为谷子根际土壤最丰富的纲,粪壳菌纲是子囊菌门第二大纲,具有更快的进化速率[46]。撂荒地与谷子根际土壤在粪壳菌纲水平上差异不显著,但在目水平,谷子根际土壤粪壳菌目相对丰度是撂荒地的2倍以上。有研究表明,粪壳菌目有降解纤维素及腐烂有机质的作用[47]。撂荒地多年未种植作物,纤维素及有机质较少,故粪壳菌目真菌少。在科水平,轮作土壤中毛壳菌科相对丰度低于连作土壤,被孢霉科、球腔菌科相对丰度高于连作土壤,被孢霉科真菌多数属于腐生菌[48],说明轮作地块有益菌要多于连作。
作物连作会导致土壤理化性质、土壤酶活性等改变[49-50],和土壤微生物组成及丰富度密切相关[17]。本研究中轮作土壤养分高于连作土壤[36],且碱解氮、有效磷、有机质、脲酶活性均显著正相关,真菌群落多样性指数也受土壤理化性质、酶活性的影响,其中Chao1 指数、Observed species 指数与多酚氧化酶、过氧化氢酶、蔗糖酶活性呈显著正相关,说明谷子根际土壤真菌受酶活性影响较大,受土壤理化性质影响较小。
连作障碍的主要原因是微生态失衡,有益真菌减少和病原真菌增加等[11,23]。本研究发现,在属水平,被孢霉菌在CR 中相对丰度最高,与pH、有效磷、有机质、碱解氮、脲酶、过氧化氢酶呈正相关,是CR 的标志物。被孢霉菌属于腐生真菌,是土壤有益微生物,在富含有机质的土壤中丰度较高,为土壤碳和养分转化的关键微生物[50-51];此外,被孢霉可降解纤维素、半纤维素和木质素[52-54],对改变土壤微生物的生境有一定影响。本研究中被孢霉菌在连作后相对丰度降低,表明谷子轮作有利于有益真菌的繁殖,连作会导致有益真菌减少。另外,亚隔孢壳菌、链格孢霉菌在连作土壤中相对丰度较高,推测谷子连作后病原真菌会增加。链格孢霉菌和亚隔孢壳菌均为植物病原菌,可引起黑斑病、叶枯病等多种真菌病害[55-56],其中一些毒素还能在体外或大鼠体内产生遗传毒性[55]。本研究中亚隔孢壳菌是TC 中的生物标志物,亚隔孢壳菌和链格孢霉菌是FC 中的生物标志物,说明谷子连作会导致病原真菌的增加,且连作年限不同,病原真菌的种类也有所不同。
本研究还发现,谷子连作导致病原真菌增加的同时也会产生生防菌。粉红螺旋聚孢霉是一类重要的菌寄生菌,这类菌可以寄生病原真菌,抑制病原菌的侵染,还可通过拮抗作用,对营养物质的竞争和诱导植物抗性等多种方式抑制病害的发生与扩展[57-59],同时具有固氮和拮抗镰刀菌作用[60]。本研究中粉红螺旋聚孢霉在CR 中极少(0.09%),TC 中最高(9.95%),到FC 有所降低(3.54%),推测在谷子连作土壤中病原菌较多,产生的生防菌也多,且在TC 中的病原菌最多,生防菌粉红螺旋聚孢霉菌也产生的最多。
本研究还发现一个有趣的现象,即随连作年限增加,谷子根际土壤环境有好转的趋势。Alpha 多样性分析结果显示连作改变了谷子根际土壤真菌群落丰富度和多样性,CR 真菌群落丰富度最高,随种植年限增加呈下降趋势,且差异显著,表明谷子连作破坏了土壤的稳定结构,降低了土壤真菌的丰度和均匀度;但主坐标分析显示随连作年限增加,谷子根际土壤真菌群落有恢复趋势,TC 和FC 真菌群落在构成上十分相似,与CR、CK 相比,真菌群落结构改变十分明显,说明谷子对于连作十分敏感,但FC 与CR在PC1 上的投影距离比TC 与CR 在PC1 上的投影距离要短,说明随连作年限的增加,这种改变并未进一步加剧;另外,土壤环境因子中碱解氮、有机质、脲酶含量也是在FC 中高于TC,进一步佐证连作5年的谷子根际土壤环境有一定好转。在大豆[61-62]、怀牛膝(Achyranthes bidentata)[63]等作物上也有类似报道。
3.2 结论
本研究利用真菌ITS 高通量测序技术,探究了连作与轮作条件下,谷子根际土壤真菌群落的分布特征。结果表明,谷子轮作根际土壤真菌群落多样性高于谷子连作。谷子根际土壤真菌优势菌门为子囊菌门、担子菌门,优势菌纲为粪壳菌纲、座囊菌纲以及盘菌纲;在目水平谷子根际土壤粪壳菌目相对丰度较高,说明谷子根际土壤中含有较多的纤维素及有机质;在属水平谷子轮作土壤被孢霉菌等腐生菌含量较高,而连作土壤中亚隔孢壳菌、链格孢霉菌等病原菌增加,说明谷子轮作根际土壤真菌环境优于谷子连作根际土壤,连作会导致腐生菌减少,病原菌增加。但是,连作5 年的谷子根际土壤环境好于连作3 年,说明随连作年限增加,谷子根际土壤真菌群落有恢复趋势。病原真菌链格孢霉菌、亚隔孢壳菌以及生防菌粉红螺旋聚孢霉菌在连作土壤中相对丰度高,暗示谷子连作与病害发生有紧密关联,对克服谷子连作障碍提供了有用的参考依据。
猜你喜欢
当代陕西(2021年21期)2022-01-19
中国土壤与肥料(2021年5期)2021-12-02
今日农业(2021年11期)2021-11-27
天然产物研究与开发(2018年3期)2018-05-07
中国化肥信息(2018年12期)2018-03-01
新农业(2017年24期)2018-01-30
中国蔬菜(2016年8期)2017-01-15
现代农业(2015年5期)2015-02-28
中国土地科学(2011年12期)2011-03-20
植物营养与肥料学报(2010年6期)2010-10-26
