植物lncRNA及其对低温胁迫响应的研究进展
2023-08-15 08:29陈晓于茗兰吴隆坤郑晓明逄洪波
生物技术通报 2023年7期
陈晓 于茗兰 吴隆坤 郑晓明 逄洪波
(1.沈阳师范大学生命科学学院,沈阳 110034;2.沈阳师范大学粮食学院,沈阳 110034;3.中国农业科学院作物科学研究所 农业农村部粮食作物基因资源评价利用重点实验室,北京 100081;4.海南三亚中国农业科学院国家南繁研究院,三亚 571700;5.国际水稻研究所,菲律宾马尼拉 DAPO box 7777)
植物是固着生物,相比能够灵活运动的动物而言,更容易受到各种胁迫的影响[1]。低温属于非生物胁迫的一种,可以对植物造成巨大的伤害。首先,低温能够影响植物地理分布,限制植物生长区域,决定植物的种植范围。中国一些热带水果(如榴莲、椰子、芒果)大都生长在海南等南方地区,北方少有种植。此外,低温还会对植物的生长和发育带来负面影响,有时甚至会造成植物死亡,如木薯叶片发生黄化、对顶端分生组织造成伤害,从而造成发芽延迟,更严重的会使生长受阻甚至死亡[2]。在低温影响下,棉花很容易造成生长发育的停止、代谢功能的紊乱和阻碍授粉等伤害[3]。对于作物来说,低温危害更为严重,低温会导致作物产量和品质下降[4],甚至绝收,直接影响农民的经济收入和国家粮食安全。随着全球人口不断增加和气候变暖,低温冷害等极端天气发生的频率大大增加,给农业生产带来了巨大的损失[5]。
植物进化出了复杂的防御机制以改善低温造成的不利影响,如细胞膜结构的改变[6-8]、含水量的下降[9]、脱落酸含量的增多[10]、活性氧的清除[11]等。近年来,随着长链非编码RNA(long non-coding RNA, lncRNA)的发现,lncRNA参与植物抵抗逆境胁迫成为了研究热点。本文就lncRNA的定义、来源、分类等多方面进行了概述,并且总结了lncRNA在响应植物低温方面的研究进展。
1 lncRNA概述
1.1 lncRNA的定义和生物学特征
非编码RNA(non-coding RNA)根据功能可以分为调节型和组成型2种[12]。lncRNA属于调节型,是存在于细胞核和细胞质中、由体内基因组转录产生的、长度大于200 nt、含有1个少于100个氨基酸的开放阅读框的一类转录本[13-14],称为长链非编码RNA(图1-A)。lncRNA与mRNA相似,都具有5'端的帽子结构和3'端的polyA尾巴,而且在转录后加工、启动子区域的表观遗传标记、二级和三级结构的形成以及发育阶段等特异性方面都具有与mRNA相似的特点;不同之处在于,lncRNA比mRNAs的转录本更短,外显子更少,且在物种之间缺乏序列保守性[15]。大多数lncRNA的转录是由RNA聚合酶Ⅱ完成,少数是由RNA聚合酶III、Ⅳ、Ⅴ的作用完成[16]。相对于进行蛋白质编码的mRNAs,lncRNA的表达水平和序列保守性较低,故被称为“转录垃圾”或者“转录噪音”[17]。随着下一代测序技术和生物信息学方法的发展,lncRNA被大量识别并肯定,越来越多的证据表明,lncRNA在植物根的发育、春化、光形态建成、花粉发育和雄性不育中均起重要作用[18]。Hu等[19]在棉花中鉴定了35 802个lncRNAs,其中,在无纤维品系和有纤维品系中,优先表达的lncRNA分别有645和651个;Salih等[20]指出lncRNA可能参与蛋白质磷酸化、植物激素代谢等生物过程来控制棉花纤维的发育,而且发现在纤维发育第8天的lncRNA的含量远大于纤维发育开始的含量,表明lncRNA在棉花纤维发育中可能起到负调控的作用。Wunderlich等[21]在拟南芥中发现配子体发育所需的热休克基因HSFB2a是由lncRNA控制。lncRNA还可以控制水稻、棉花、玉米等的生殖发育过程,造成雄性不育[22]。此外,lncRNA已经被证实可以响应植物的各种胁迫,如拟南芥中发现的lncRNA-DRIR在干旱条件下被激活,增强了耐受性[23]。
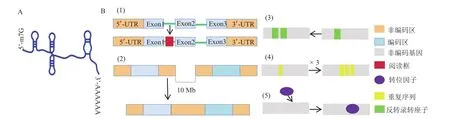
图1 lncRNA的结构及来源Fig.1 Structure and origin of lncRNA
1.2 lncRNA的来源
1ncRNA的表达水平低且序列保守性差,使得其在物种进化过程中受到的阻力小,来源多样。目前,推测lncRNA的形成可能来自于以下这5种情况[24-25](图1-B)[12]:(1)阅读框的插入。已有的编码序列会与突然插进来的阅读框重新整合形成lncRNA;(2)染色体重新组合。远距离的非转录区串联起来产生含有多个外显子的lncRNA;(3)非编码基因的逆转录转座的复制;(4)连续重复事件在非编码RNA内形成相邻的重复序列从而产生新的lncRNA;(5)转位因子的插入。
1.3 lncRNA的分类
随着lncRNA受国内外研究人员的广泛关注,越来越多的lncRNA被发现,因此,建立了相关的lncRNA数据库,如CANTATA数据库(https://yeti.amu.edu.pl/CANTATA/)、NONCODE数据库(https://www.noncode.org)、PlncRNADB数据库(https://bis.zju.edu.cn/PlncRNADB/index.php)、PLNlncRbase数据库(https://bioinformatics.ahau.edu.cn/PLNlncRbase)、PNRD数据库(https://structuralbiology.cau.edu.cn/PNRD/index.php)等,其中CANTATA和PNRD是2个最大的植物lncRNAs数据库[26]。面对数量众多的lncRNAs,可以选择根据不同的分类依据将lncRNA进行归类,从而方便进行研究和分析。
首先,根据lncRNA与邻近蛋白质编码基因位置的不同,lncRNA被分为5种类型(图2)[27],这是最常见的一种分类方式。(1)正义lncRNAs(sense long non-coding RNAs),即从具有相同启动子的蛋白编码基因重叠区域转录而来的lncRNA;(2)反义lncRNAs(antisense long non-coding RNAs),即从蛋白编码基因的反义链转录而来,天然反义转录本(natural antisense transcripts, NATs)就是最典型的例子;(3)内含子lncRNAs(intronic lncRNAs),即存在于转录本的内含子区域的lncRNA;(4)基因间lncRNAs(long intergenic non-coding RNAs),指起源于2个基因之间区域的lncRNA;(5)双向lncRNAs(bidirectional long non-coding RNAs),指1ncRNA位于与互补链相邻的编码基因转录起始位点附近,但转录方向相反[27-31]。
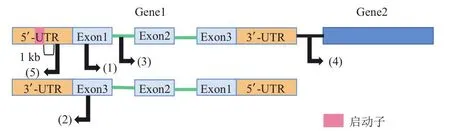
图2 基于基因组位置的lncRNA分类Fig.2 Classification of lncRNAs based on genomic locations
其次,根据作用机理(图3),lncRNA可以作为信号分子、支架分子、引导分子和诱饵分子:(1)信号分子指与转录因子结合或参与信号通路来间接调节邻近基因在时空上的表达的lncRNA。如拟南芥AtR8作为信号分子调控WRKY转录因子的表达来响应低氧胁迫[32]、水稻LDMAR充当信号分子作为小RNA的前体去调控水稻光雄性不育[33]等。(2)支架分子是指可以将不同的蛋白质结合在一起来作为核糖核蛋白复合物支架的lncRNA。如拟南芥APOLO作为支架分子通过影响染色质环的形成来调控极性生长素转运[34]。(3)引导分子,即与核糖核蛋白复合物或者染色质修饰酶结合并将其引导到专门位置的lncRNA,其中lncRNA招募染色质修饰物的工作模型可能有3种:组蛋白修饰剂直接结合lncRNA、lncRNA由一种桥蛋白(如RNA结合蛋白)与染色质修饰物相互作用,形成“超复合体”以及低温胁迫条件下lncRNA和染色质修饰剂形成相分离系统[35]。例如,根瘤MtRBP1蛋白直接与lncRNAEnod40结合,将MtRBP1从植物细胞的核斑点重新定位到细胞质颗粒中发挥重要作用[36]、水稻中的lncRNA-LAIR作为引导分子通过激活LRK表达增加水稻产量[37]。(4)诱饵分子,即通过招募转录因子、染色体修饰、调控分子等RNA结合蛋白来间接调控蛋白编码基因表达的lncRNA。此外,lncRNA还可以作为miRNAs和siRNAs生物合成的前体调节mRNA前体的选择性剪接,或者作为内源性靶标模拟物(eTM)竞争各种miRNAs[38]。lncRNA能够用其相似的结构与miRNA逐一竞争性结合,形成lncRNA/miRNA/mRNA复合物,从而产生功能上的二次调控。竞争性结合通常会发生在lncRNA和mRNA上的miRNA结合靶点相同的情况下。lncRNA与miRNA之间的配对可以在自由或特定条件下发生。这意味着基因表达或生长发育情况下,lncRNA与mRNA通过竞争空间、配对亲和等方式,争夺miRNA的结合位点,影响miRNA的作用目标和活性。过量的lncRNA可能会将miRNA绑定到lncRNA上,而不是目标mRNA上,从而降低了miRNA对mRNA的结合和对mRNA的抑制作用,进而影响miRNA调控下游基因的表达和信号途径。这种lncRNA与miRNA之间的竞争性结合提供了一种新的机制,链式调控激活了miRNA的新模式,这对正常细胞的功能调节及其细胞周期、迅速响应环境等平衡非常重要。如拟南芥中受磷饥饿诱导的miR399所作用的靶基因是PHO2(PHOSPHATE2),但lncRNA-IPS1作为miR399的靶基因模拟物可使PHO2的表达量增加,进而维持磷的稳态平衡[39]。
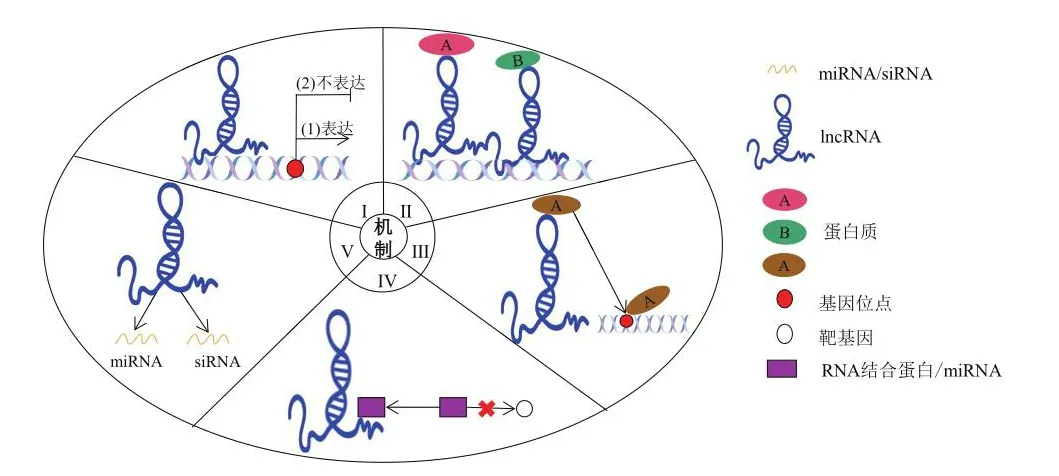
图3 lncRNA的作用机制示意图Fig.3 Schematic diagram of lncRNA action mechanisms
再次,可以根据对DNA序列的影响不同,将lncRNA分为反式作用的lncRNA(trans-lncRNA)和顺式作用的lncRNA(cis-lncRNA)两种[12]:translncRNA是指调控远端基因表达的lncRNA;cislncRNA是指调控基因组邻近基因表达的lncRNA,如在植物发芽中起作用的as-DOG1[40]。
1.4 lncRNA的作用特点
lncRNA与miRNA或siRNA相比,具有更大的长度和更加复杂的空间结构。近些年研究表明,lncRNA在调控细胞基因表达、染色质修饰以及蛋白质稳定性等多个方面发挥着重要作用,其特点和作用机制如下:(1)高度组织特异性。每个组织/细胞类型中的lncRNA表达模式都具有高度的特异性,这种组织特异性与lncRNA结构的多样性和其特征序列的相对稳定性密切相关。(2)与染色质结构和修饰密切相关。lncRNA能够与染色质的修饰酶、转录因子、RNA多聚酶等相互作用,从而影响染色质的结构和发生作用的基因表达。(3)通过多种方式调控基因表达。lncRNA可以调控基因表达的多个环节,包括转录因子的表达、二级结构的稳定性、蛋白质稳定性、转录后加工等。同时,lncRNA还可以与mRNA特异结合,调控RNA的剪接和剪切。(4)亚细胞定位复杂。lncRNA能够定位到细胞核、细胞质或者细胞核和质之间,同一lncRNA在不同环境或者细胞类型之间的亚细胞定位也可能不同。(5)功能多样性高。lncRNA的功能与结构密切相关,可以调控基因表达、RNA的剪接、RNA的稳定性、细胞周期、细胞凋亡等多个细胞生命活动。综上所述,lncRNA具有较高的组织特异性、与染色质结构和修饰密切相关、多种调控方式、亚细胞定位复杂、功能多样性高的特点,其作用机制与基因表达、细胞周期、细胞凋亡等多个细胞生命活动密切相关。
2 lncRNA响应植物低温胁迫
最初,lncRNA的生物学功能来自于哺乳动物和果蝇X染色体失活与性染色体剂量补偿遗传机制[41-42]和果蝇双胸复合的遗传分析[43]的研究。后来lncRNA在植物耐冷过程中的调控作用逐渐被人们所认识。针对表观遗传调控机制,目前已经证实了lncRNA在植物抗寒适应中的调节作用。具体地说,植物体内lncRNA通过与染色质修饰酶(如组蛋白去乙酰化酶和DNA甲基转移酶)结合调控染色质的修饰情况,从而影响靶基因的表达。例如,在植物的耐冷适应过程中,lncRNA可以介导组蛋白去乙酰化酶的定位,促进特定基因启动子区域的去乙酰化修饰,从而促进相关抗寒基因的表达[35];或者lncRNA可通过介导某些转录因子的调节,影响基因表达水平[44]。此外,lncRNA在RNA后转录调节方面也具有重要的调控作用。例如,lncRNA可通过与microRNA(miRNA)结合,从而阻止miRNA与靶基因的结合、抑制miRNA的活性,进而促进靶基因的表达[38];或者lncRNA可通过与蛋白质相互作用,从而调控基因的转录后修饰以及RNA的加工和分解等多种生物学过程[45]。因此,lncRNA通过与染色质修饰酶、转录因子、miRNA和蛋白质等分子相互作用,参与到植物耐冷的调控过程中,具有重要的表观遗传调控机制。下面将从模式植物拟南芥、农作物和园艺作物这3个方面来进行lncRNA响应植物低温胁迫的研究进展总结。
2.1 拟南芥lncRNA
植物中参与冷胁迫的转录调控网络最具特性的是CBFS(C-repeat-binding factor)信号通路[46],目前,大多数上游转录因子通过调节CBF的表达来帮助植物度过低温[47]。Kindgren等[48]在拟南芥基因组的冷敏感区域发现一个lncRNA-SVALKA,与CBF1的反义链有重叠序列。相关分子生物学试验表明,SVALKA转录会影响asCBF1的表达,而asCBF1转录会导致RNAPII碰撞进而抑制CBF1的表达,要在降低适应成本的情况下最大限度地增强植物耐冷性,Svalka-asCBF1级联提供的这种严格控制CBF1表达和时机的机制为其提供了可能。Liu等[49]在拟南芥中筛选到lncRNA-CIL1通过影响活性氧途径或者渗透调节物质来响应冷胁迫,转录分析发现CIL1是通过调节下游低温胁迫反应基因的表达使得植物作出相应反应。Moison等[44]发现在寒冷条件下,拟南芥的转录因子WRKY42会与lncRNA-APOLO相互作用,APOLO通过与RHD6启动子的结合来调节根毛伸长。此外,在拟南芥中发现的TE-lincRNA11195也对寒冷有所响应,进一步分析发现水杨酸刺激反应基因很有可能就是其潜在靶标[50]。以上研究表明,lncRNA在拟南芥中通过各种方式来响应低温胁迫。
植物开花调控网络中的关键抑制基因是春化基因FLC(Flowering Locus C),长时间的低温会抑制FLC的表达,从而促进了植物的早期开花[51]。研究发现,关键基因FLC中检测到3种类型的lncRNAs:COLDAIR、COLDWRAP和COOLAIR[52-53]。Heo等[54]在拟南芥中找到了一个lncRNA-COLDAIR,并发现COLDAIR与PRC2(POLYCOMB REPRESSION COMPLEX 2)的组件相关联,并将PRC2指向FLC,在PRC2与FLC染色质的募集中发挥作用,以建立春化稳定的FLC沉默。Kim等[55]从被抑制的FLC启动子中鉴定出一个长非编码RNA-COLDWRAP,其通过形成一个抑制性的基因内染色质环,将多聚体保留在FLC启动子上,来形成稳定的FLC沉默;与COLDWRAP和COLDAIR不同,COOLAIR是一种FLC反义转录本,具有交替的多聚腺苷和多个剪接变异体,可能参与FLC染色质状态的转换,通过在FLC中富集H3K36me3来抑制FLC的转录水平[56-57]。综上,在春化过程中,位于FLC的lncRNA通过增加PRC2占有率、提高FLC染色质H3K27me3水平等方式来维持FLC稳定的抑制状态。
2.2 农业经济作物lncRNA
俗语说得好,“民以食为天”。只有保证农作物的产量和品质,才能确保国家的粮食安全和国民的生活水平。作为世界范围内重要的粮食作物水稻(Oryza sativaL.),全球有近一半的人口以大米为主食。但因为耕地面积的减少和人口的快速增长,水稻产量以及产量的增加已经成为关系到全球经济和粮食安全的一个重要问题[58]。低温是影响水稻生产的一个主要限制性因素。Yuan等[59]发现低温条件下,部分DPA lncRNAs在水稻耐冷相关的QTL中特异性富集,并参与了冷暴露期间催化活性的负调节,表明lncRNA在水稻的抗逆性中发挥重要作用。Shin等[60]发现一个可能的lncRNA-Chr03G0008,其只在冷处理的幼苗中表达。Gao等[61]在低温胁迫下的水稻种子中鉴定出一种与种子活力相关的lncRNA-SVR,发现是SVR与顺式基因SAUR家族成员相互作用,同时SVR中的基因突变导致种子萌发延迟,说明lncRNAs可能通过控制生长素激活的信号通路来调节水稻种子活力。低温下,Leng等[62]在耐冷品种空育131和敏感品种东农422的孕穗期共鉴定出566个lncRNAs存在差异表达,其中8个lncRNAs与低温响应有关,但具体的耐冷机制有待进一步研究。同样,作为粮食作物的小麦(Triticum aestivum)中也发现存在耐冷相关lncRNA资源。东农冬麦1号作为北方高寒地区首例能安全越冬的栽培品种,在其中分离得到9 971个lncRNA,可以在不同程度的低温下表现出不同的应答,如:-10℃条件下,1 260个lncRNA差异表达,其中591个上调表达,669个下调表达[63]。Díaz等[64]在低温处理的硬粒小麦CBW0101中发现了31个差异表达的lncRNA,24个表达上调,7个表达下调;预测lncRNA Traes_2BS_7A04BF5D5以WCOR413冷驯化基因为靶标,lncRNA Traes_2DL_ABD08139B以开花促进因子为靶标。此外,东农冬麦1号中的lncR9A、lncR117和lncR616可以作为竞争性内源lncRNA调控tae-miR398与mRNA-TaCSD1在低温胁迫下的协同作用,进而影响东农冬麦1号的抗寒性[65]。
低温严重影响热带作物木薯(Manihot esculentaCrantz)的生长,分析响应低温的lncRNA,有利于为木薯抗逆性研究提供分子基础,更好地让木薯适应寒冷环境。Suksamran等[66]鉴定出2 229个lncRNAs(Me-lncRNAs)均匀分布木薯染色体上,250个潜在的新的lncRNA存在显著的差异性表达,其中,低温条件下86个Me-lncRNAs表达增加,96个Me-lncRNAs表达降低。在木薯主栽品种‘60444’中鉴定到一个受低温胁迫调控的lncRNA(cold-responsive lncRNA5,CRR5),其与一些重要的蛋白激酶(如谷胱甘肽S-转移酶,AUX/IAA蛋白)协同表达响应低温胁迫[67]。Li等[68]在木薯中鉴定出一种新的对冷胁迫反应的正向调节因子—lncRNA1(CRIR1),通过招募RNA伴侣MeCSP5来提高mRNA的翻译效率响应低温胁迫。
棉花是世界上重要的经济纤维和油料作物,也是一种喜温植物。遭受低温冷害后的棉花会代谢紊乱、乙烯含量增加,从而加速叶片衰老,甚至会诱发黄萎病,造成棉花减产[69]。在陆地棉(Gossypium hirsutumLinn.)中,lncRNA基因XH123参与棉花苗期冷胁迫的适应性调控,XH123沉默会引起冷调控基因PIF3、叶绿体基因和氧化还原反应基因的差异表达,造成超氧化物歧化酶、脯氨酸的显著升高和叶绿体结构的降解,进而引起棉花的生理和细胞结构差异来抵御冷胁迫带来的危害[70]。同样,苜蓿(Medicago sativa)作为全球性栽培的牧草,抗寒性决定其冬季存活和来年可再生产的能力[71]。苜蓿中已经鉴定出部分lncRNA在叶和根中对低温处理有响应,且存在组织特异性。通过整合lncRNA MtCIR1及其潜在靶基因MtCBF,发现一个可能在适应低温胁迫中发挥关键作用的lncRNA- mtcbfs调控网络[72]。
番茄(Lycopersicon esculentumMill.)属喜温作物,对低温比较敏感。遭遇低温的番茄,果实上会出现瑕疵、不能成色甚至可能会引起由病原菌造成的疾病,严重地影响番茄品质[73]。Wang等[74]在番茄中鉴定1 411个lncRNA属于基因间lncRNA和反义lncRNA;其中239个lncRNA在冷害果实中的表达模式存在显著差异,138个lncRNA表达上调,101个lncRNA表达下调;同时发现41个lncRNA是33个miRNA的前体,186个lncRNA是45个miRNA的靶标,推测lncRNA可能通过竞争与共享miRNA结合来调控mRNA的表达,来进一步在低温中发挥作用。在正常温度和冷藏条件下,Zuo等[75]发现了380个差异表达的长非编码RNA,这些lncRNA能够以顺式或反式的方式调控参与冷诱导损伤的关键酶的表达;Baruah等[76]在辣椒低温处理下发现了2 101个lncRNA差异表达,其中部分可以作为miRNAs的前体响应胁迫。
Wang等[77]发现低温条件下葡萄(Vitis viniferaL.)中的lncRNA存在差异性表达,233个已知的lncRNAs和284个新发现的lncRNA显著上调,144个已知lncRNAs和182个新发现lncRNA显著下调;且冷诱导葡萄lncRNAs的靶基因在顺式调控关系中比在反式中更多,说明顺式调控关系中的靶基因可能与冷应激反应更相关。此外,在葡萄中发现了212个lncRNAs作为miRNAs的靶标,涉及150个miRNAs。香蕉(Musa nanaL.)也很容易受到低温的影响,尤其在冬季和早春。Liu等[45]在冷胁迫的中国野生香蕉中鉴定出12 462个lncRNAs发现,lncRNAs能够通过调节类黄酮的生物合成、调节丝氨酸/苏氨酸或双特异性蛋白激酶等来响应低温胁迫。研究认为在野生香蕉中,TCA循环和硫传递系统途径等生物途径可能会响应寒冷,并受到野生香蕉中lncRNAs的调控。
3 展望
低温冷害严重影响植物的生长发育。近年来,随着生物信息学的广泛应用和测序技术的改进,越来越多的研究表明lncRNA是调控植物耐冷性的重要分子(表1)。因此,利用lncRNA来提高植物的耐冷性已成为当前植物抗逆研究领域热点之一。首先,研究人员可以分析鉴定植物在低温胁迫下的lncRNA,并进一步确定与低温适应相关的lncRNA,以此为基础开展基因功能和调控机制的研究。其次,也可以利用 CRISPR/Cas9等基因编辑技术对lncRNA相关基因进行靶向编辑,如破坏lncRNA的表达,通过转化基因的方式提高植物抗寒性。此外,研究人员也可以利用RNAi技术对植物低温适应相关的基因进行表达下调,从而提高植物的抗寒性。除此之外,利用表达调控元件可以实现lncRNA的特异性表达,并且可以通过这种方式将lncRNA所带来的抗逆能力迅速广泛地应用于各类农作物。
尽管lncRNA对植物低温胁迫的研究已经取得了一些进展,但相较于动物而言,植物lncRNA的研究还处于一个相对早期的阶段,还存在很多问题亟待解决。首先,植物lncRNA数量庞大、长度较长、生物学功能复杂、鉴定难度较高,需要增加样本数据、建立更加精准的筛选算法,开发更加高效和精确的计算算法和大规模测序技术进行验证,以确定lncRNA的可靠性和准确性。其次,由于植物lncRNA种类繁多、表达量低、作用机制复杂,其在植物的各种生长发育和应激响应的调控机制中的准确作用尚未完全明确。当前植物lncRNA的功能研究主要集中在通过基因表达谱和功能富集分析以及互作网络分析等方法预测其生物学功能。在未来的研究中,可以结合各种生物实验技术来进一步验证预测的功能,精确阐明植物lncRNA的生物功能和调控机制。同时,加强多方位的合作研究,如在生物大数据、计算模拟、生物统计和机器学习等方面开展合作,提高研究效率和精度。再次,植物lncRNA命名混乱,目前还没有建立起一个统一、全面、官方的lncRNA命名规则,研究人员应该加强规范化管理,在国际上建立一个全球化的lncRNA命名标准,并且联合相关部门监管,以建立可持续的、可靠的lncRNA命名规则体系,以便不同地区的研究者能有效地分享他们的研究结果。总之,植物lncRNA的研究,需要不断开展基础和创新性的研究,加强合作与实验验证,并制定完善的命名规则,优化方法和体系,以不断提升lncRNA在植物抗逆和作物耐冷育种研究方面的应用价值。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
真空与低温(2022年6期)2023-01-06
畜牧兽医学报(2022年3期)2022-03-30
中国畜牧兽医(2022年1期)2022-02-15
现代塑料加工应用(2021年5期)2021-02-28
现代泌尿外科杂志(2019年10期)2019-10-31
生物学通报(2019年2期)2019-06-15
劳动保护(2018年8期)2018-09-12
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10