
我国近5年类NADC30 PRRSV毒株序列的重组及限制性片段长度多态性分析
2023-09-25 11:33谷尚品侯晓璇王孟月王小娟谭菲菲田克恭
中国兽医学报 2023年8期
王 彪,谷尚品,侯晓璇,王孟月,王小娟,谭菲菲*,田克恭,*
(1.河南农业大学 动物医学院,河南 郑州 450046;2.国家兽用药品工程技术研究中心,河南 洛阳471000)
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)引起的一种高度传染性疾病,给养猪业造成了重大的经济损失[1]。PRRSV属动脉炎病毒科,是一种有包膜的单股正链RNA病毒,基因组长度约为15 kb。根据基因序列和抗原特性之间的差异,可进一步分为欧洲型(PRRSV-1)和美洲型(PRRSV-2)[2-3]。
1996年PRRSV首次在我国华北地区报道[4]。随着PRRSV不断变异和传播,以高热、高流产率、高病死率为特点的高致病性PRRS(highly pathogenic PRRSV,HP-PRRS)于2006年暴发,重创了我国养猪业,造成超过2 000万头猪受到影响,其代表性毒株为JXA1[5]。2014年,周峰等[6]等报道了河南地区类NADC30的流行,其非结构蛋白2(nonstructural protein 2,Nsp2)与美国2008年分离的NADC30毒株均存在131(111+1+19)个不连续氨基酸缺失,因此将此类毒株命名为类NADC30。随后,类NADC30毒株的临床检出数量急剧增加,并且病毒的重组事件频频发生,使PRRSV多样性显著扩大[7]。
Nsp2和ORF5是PRRSV基因组中两个高变区域,其中ORF5基因在免疫应答的调节中起着关键作用。基于PRRSV ORF5序列遗传演化分析和序列之间的同源性,可将PRRSV-2分为9个不同的谱系:Lineage 1~Lineage 9[8],该方法也是目前PRRSV-2比较公认的分类法。另一种序列分类方法是1998年美国学者[9]以PRRSV-2建立的限制性片段长度多态性(restriction fragment length polymorphis,RFLP)分类方法,根据ORF5序列中MluⅠ、HincⅡ和SacⅡ的酶切模式,并分配3个数字代码进行命名。RFLP分类方法在美国养猪行业普遍使用,可用于区分PRRSV-2野毒株与VR2332弱毒疫苗株。
2020年10月,美国中西部养猪场报道了HP-PRRSV RFLP1-4-4 Lineage1C变异株,其对猪致死率高达17.5%[10],且疫苗对其保护效果不佳。鉴于我国2003年就存在PRRSV RFLP1-4-4模式[11],并且PRRSV毒株频繁发生突变和重组。因此本研究对实验室类NADC30 PRRSV 株与参考毒株进行遗传演化分析与RFLP分析,旨在了解我国近年来类NADC30的遗传演变规律,为PRRSV流行株的防控提供借鉴资料。
1 材料与方法
1.1 样本信息本实验室自2016-2020年从我国15个省份(河南、江苏、山东、山西、陕西、天津、重庆、浙江、吉林、福建、辽宁、江西、黑龙江、河北、四川)的猪场采集1 747份组织和血清样品中检测到263份类NADC30样品,选择64份代表性样品进行全基因序列测定。同时从NCBI中选择21条参考毒株序列用于分析,参考序列如表1所示。
1.2 序列比对和遗传演化分析根据经典毒株VR2332序列注释对本研究中的序列进行分割,得到各毒株的Nsp2序列。利用DNAStar软件中MegAlign对64株毒株与参考毒株的Nsp2推导氨基酸序列进行比对,分析各毒株的Nsp2缺失特征。使用MEGA11(邻近法,步长值为1 000)对64株毒株与代表性毒株进行全基因组和ORF5序列遗传演化分析。
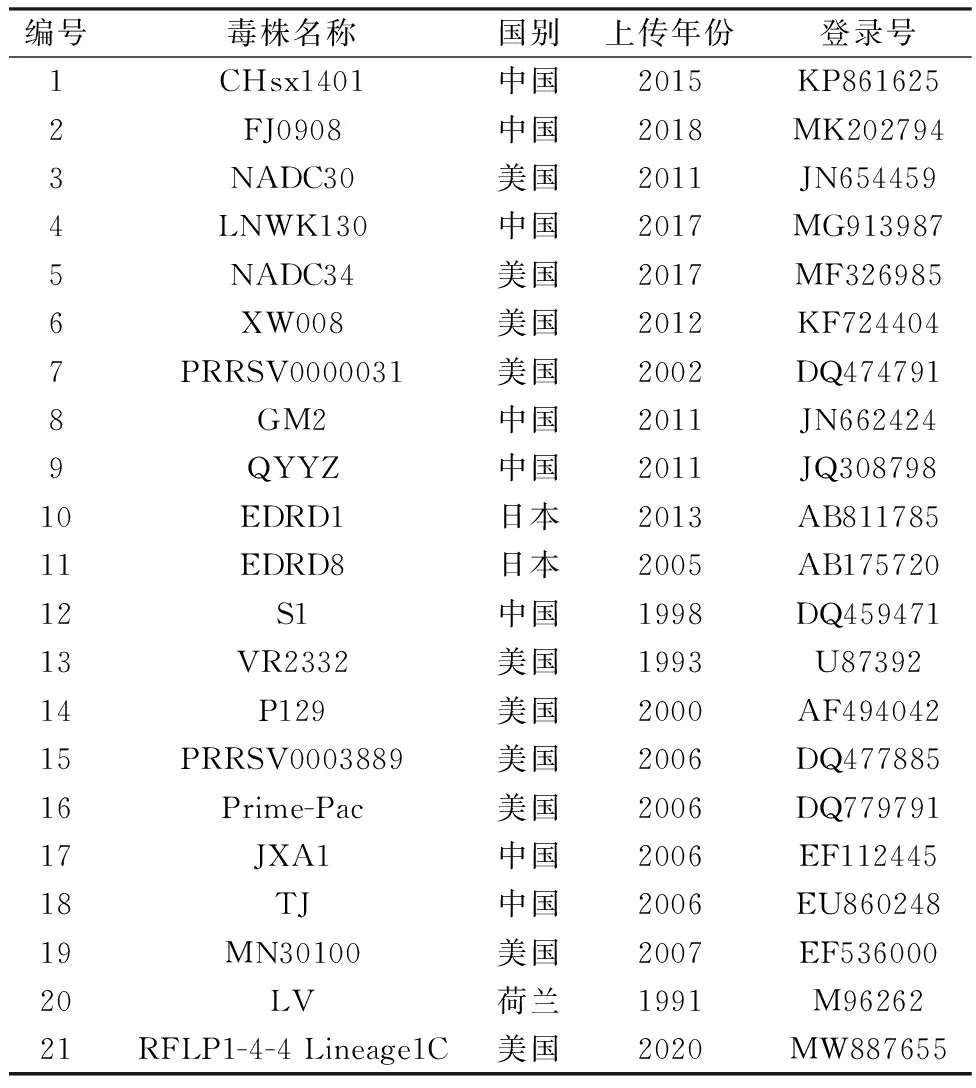
表1 PRRSV参考毒株信息
1.3 重组分析选取各谱系5个代表性毒株作为参考毒株:NADC30、NADC34、QYYZ、VR2332、JXA1。通过SimPlot(v3.5.1)软件对本研究的64条全基因组序列进行全基因组重组事件的初步检测,然后使用RDP4(v4.97)软件对重组毒株进行分析,当7种方法(RDP4、GENECONV、MaxChi、BootScan、SiScan、Chimaera、3Seq)中至少有4种方法检测到重组事件(P-val<10-6)时,认为该序列中可能有重组事件的发生。最后,计算重组毒株的相同重组断点数量占总重组毒株的比例为重组热点。
1.4 ORF5序列的RFLP模式分析以NADC30毒株ORF5序列为参考,截取本研究64条毒株ORF5基因序列。通过SnapGene软件对64条NADC30毒株ORF5序列分别进行MluⅠ、HincⅡ、SacⅡ 3种限制性内切酶的模拟酶切,依据RFLP模式分析进行划分。
2 结果
2.1 Nsp2序列特征64株分离株Nsp2推导氨基酸与参考毒株R2332比对结果显示,其Nsp2在322~432,483,504~522氨基酸位置均存在131(111+1+19)个氨基酸的不连续缺失,与代表性毒株NADC30缺失特征一致,表明64株毒株属于类NADC30毒株。此外,其中有9个毒株含有额外的氨基酸缺失(图1),毒株171365-4、2015HeNP183、161343-1、161433-57在464~468氨基酸位置缺失,毒株170546-2、180411-12、180546-27在18~23氨基酸位置缺失,毒株56full在459~489氨基酸位置缺失, 毒株181120-4在465~467氨基酸位置缺失。

橘色代表Nsp2 131个氨基酸的不连续缺失;蓝色代表毒株的Nsp2额外氨基酸缺失
2.2 遗传演化分析为了建立64株毒株与参考毒株的遗传关系,基于全基因组和ORF5序列进行遗传演化分析。全基因遗传演化分析结果(图2)显示,61株分类在Sublineage 1.8属于类NADC30亚群,其余毒株180049-1、56full、161433-57分类在Lineage 8,属于HP-PRRSV,PRRSV RFLP 1-4-4 L1C变异株分类在Sublineage 1.5,属于类NADC34亚群。而基于ORF5序列遗传演化分析结果(图3)显示,56株毒株与PRRSV RFLP 1-4-4 L1C变异株分类在Sublineage 1.8,其余8株中56full、180049-1、170407-41、161401-3、180991-3分类在Lineage 8,20200714-2分类在Sublineage 1.5,161302-14、171365-4分类在Lineage 5,属于VR2332亚群。由此可见基于ORF5的遗传演化分析分出谱系更多,表明这些毒株可能存在着重组现象。
2.3 全基因组序列重组分析为了解64株分离株是否存在重组事件,以各谱系代表性毒株作为参考毒株,使用RDP4和Simplot软件对64条毒株全基因组序列进行重组分析,64株毒株中有44株存在重组事件(表2,仅展示5种方法的结果),共5种重组模式。最主要的重组模式是以NADC30毒株作为亲本,JXA1毒株提供重组片段。其余4种重组模式分别是:以JXA1为亲本毒株,NADC30毒株提供重组片段的重组模式(161433-57、56full、180049-1);以NADC30为亲本毒株,JXA1和VR2332毒株提供重组片段的重组模式(170438-1、180450-1、2016-32、161302、171365-4、171388-1、180613-4、181425-3、181566-4);以NADC30毒株为亲本,JXA1和QYYZ毒株提供重组片段的重组模式(16448-2、170028-2、170051-4、16756-1、170097-1);以NADC30毒株为亲本,NADC34毒株提供重组片段的重组模式(20200714-2)。根据参考毒株VR2332基因序列确定重组片段的位置,结果显示重组热点主要聚集在非结构蛋白区域5′UTR~1 500 nt(Nsp1)以及5 374~8 177 nt(Nsp4~Nsp9)和结构蛋白区域的12 100~13 279 nt(ORF2~ORF4)(图4A,B,C)。
2.4 ORF5序列的RFLP模式分析结果如图5所示,64株分离株ORF5序列的RFLP模式呈现多样性,数量最多的模式为RFLP1-4-4(31株),其次是1-2-4(9株)、1-4-3(7株)、1-3-4(5株)、1-1-4(4株)、2-5-5(2株)等。结合ORF5序列的遗传演化分析结果,31株RFLP1-4-4模式中有28株与美国PRRSV RFLP1-4-4L1C变异株分类在Sublineage 1.8,2株毒株170407-41、161401-3分布在Lineage 8,1株20200714-2分类在Sublineage 1.5。RFLP1-4-3(7株)类型中56full、180049-1、180991-3分类在Lineage 8,其余4株分类在Sublineage 1.8。RFLP2-5-2模式的毒株161302-14、171365-4分类在Lineage 5,ORF5序列与经典毒株VR2332的RFLP模式相同,且ORF5序列具有99.4%和99.9%的一致性。其余模式(1-2-4、1-3-4、1-1-4)毒株均属于Sublineage 1.8。

▲表示参考毒株;◆表示美国PRRSV RFLP1-4-4 Lineage1C变异株

▲表示参考毒株;◆表示美国PRRSV RFLP1-4-4 Lineage1C变异株
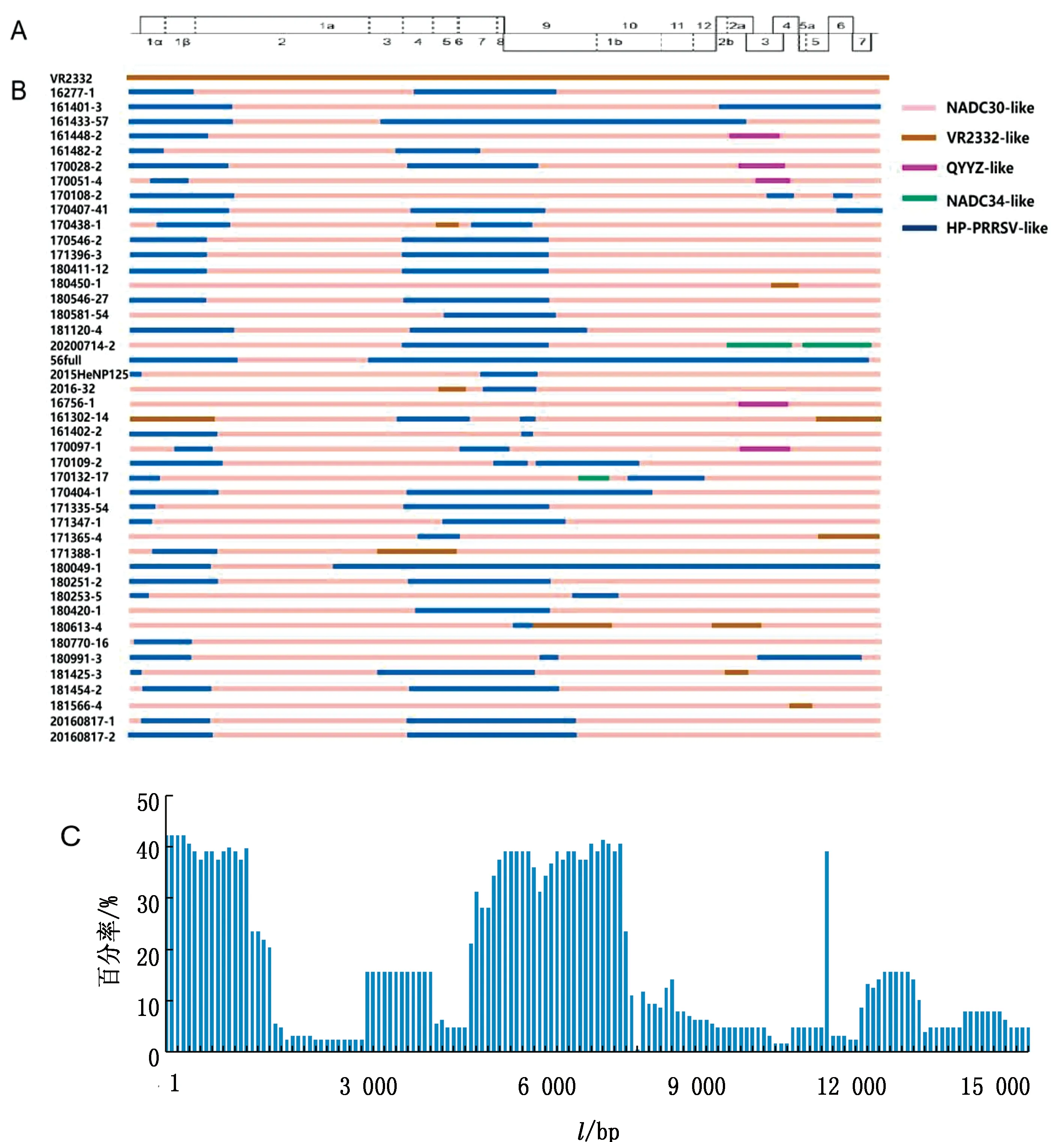
A.参照VR2332株全长基因组结构;B.重组毒株断点位置;C.重组断点位置热点分布图
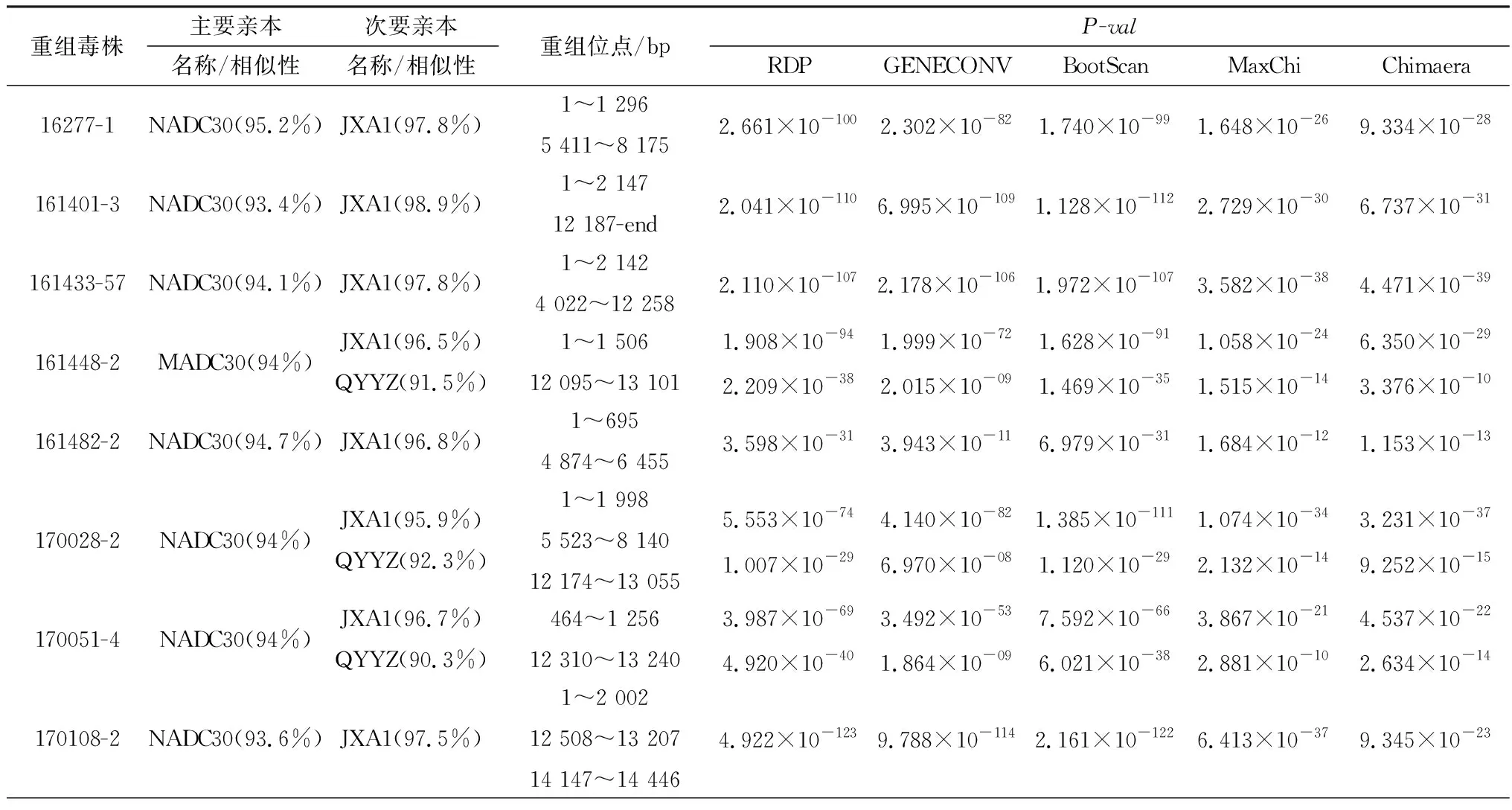
表2 类NADC30毒株重组事件信息

续表2
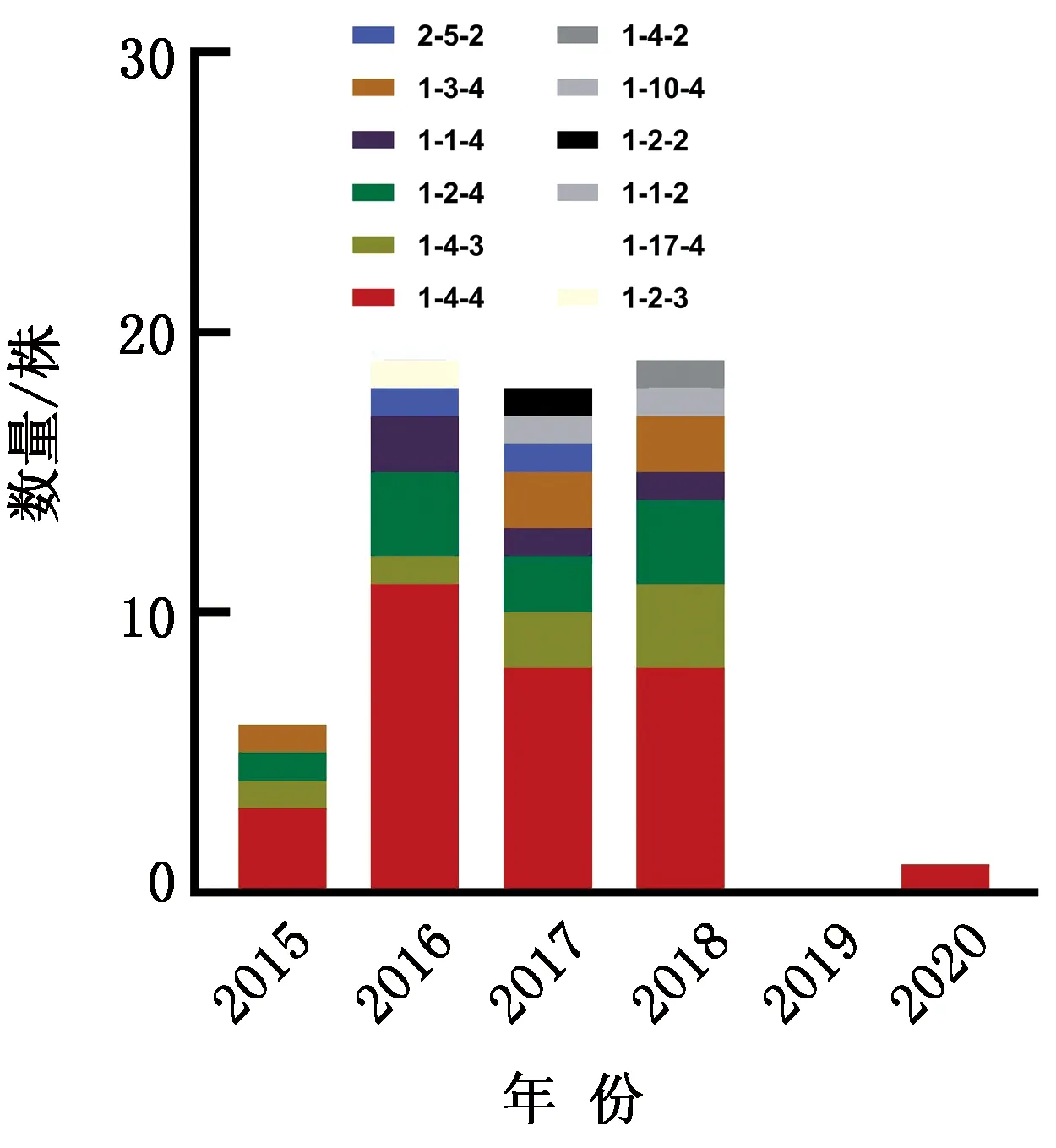
图5 64株类NADC30 PRRSV ORF5 RFLP模式
3 讨论
PRRS被认为是我国养猪业中最具流行性和破坏性的疾病,自2006年HP-PRRS暴发,PRRSV在我国的流行就经历了HP-PRRSV和类NADC30毒株之间交替优势的阶段[12]。近年来,研究表明类NADC30毒株的流行不断扩大,并逐渐成为我国的主要流行毒株[7],引起了业界的广泛关注。
Nsp2是PRRSV全基因组中高度变异的区域之一,包含缺失、重组和插入多种突变模式,常被作为区分不同毒株类型的分子标记[13]。与经典毒株VR2332相比,本研究64条Nsp2氨基酸序列除具有与NADC30毒株缺失特征一致的131个不连续氨基酸缺失以外,毒株171365-4、2015HeNP183、161343-1、161433-57在464~468氨基酸位置含有额外的缺失,这与2016年报道的分离毒株HeN1401、HeN1601研究结果相同[14]。而毒株170546-2、180411-12、180546-27、56full、181120-4在Nsp2氨基酸序列中呈现出了新的缺失,这体现了我国类NADC30毒株在Nsp2区域具有复杂的缺失模式,且在持续的变异中。但不同类型的缺失与毒力等生物学特性之间是否相关还需要进一步的研究。
PRRSV-2是我国主要的流行毒株,可根据其高变异区ORF5序列、Nsp2序列以及全基因组序列将其划分为不同的分支来探究遗传演化规律。目前,我国PRRSV-2主要流行谱系为Lineage1、3、5、8[15]。为了更全面了解类NADC30毒株的遗传演化关系,本研究分别选择Lineage 1~9的代表性毒株进行分析。基于ORF5序列和全基因组序列遗传演化分析结果发现,多个毒株在ORF5序列系统进化树和全基因组系统进化树中的分布存在差异,表明这些毒株的基因组中可能存在着重组现象,同时表明仅依赖ORF5序列分型不能够全面代表病毒的序列变异情况。
重组在PRRSV中普遍存在,我国多谱系毒株的共存会产生多种模式的重组毒株,进一步造成病毒基因组遗传的多样性,从而导致PRRS的防控形势更加严峻。2006-2011年期间,我国PRRSV一直保持着较低的重组率,随着类NADC30毒株的流行,我国重组毒株的检出数量急剧增加[16]。本研究中64株类NADC30毒株有44株存在重组事件,并表现出复杂的重组模式,重组位点几乎分布于整个基因组,其中毒株56full、180049-1、170407-41、161401-3、180991-3、161302-14、171365-44、20200714-2在ORF5区域与其他谱系毒株序列发生重组,该结果也进一步解释了这些毒株在基于ORF5序列和全基因组序列的系统进化树中位置存在差异的原因。另外,毒株161302-14、171365-4、171388-1是NADC30毒株与VR2332弱毒疫苗株之间发生重组,表明PRRSV重组不仅在野毒株与野毒株之间发生,在野毒株与疫苗株之间也时有发生,并且会改变毒株的致病性[17-19]。对44株毒株重组位点进一步分析发现重组热点主要聚集在Nsp1、4~9和ORF2~ORF4区域,有研究表明,Nsp通过不同机制调控宿主抗病毒免疫反应,其Nsp9编码的蛋白与病毒的复制过程密切相关,而ORF2、ORF3、ORF4进行编码蛋白所形成的异源三聚体是促使PRRSV感染的主要决定因素[20]。因此PRRSV在这些区域发生重组是否会导致重组毒株具有更强的增殖能力仍需进一步的研究。
目前,我国PRRSV毒株主要以分支谱系进行分类命名,但鉴于我国PRRSV毒株有美国毒株传入的先例,有时也沿用RFLP分类进行毒株的分析。2020年10月美国暴发的RFLP 1-4-4 L1C变异株具有较高的致病性,且可能是以类NADC34为亲本,类NADC30在ORF5区域提供重组片段的重组毒株[21-22]。本研究中,共发现31株是RFLP1-4-4模式,其中28株与2020年10月美国暴发的RFLP 1-4-4 Lineage1C变异株在遗传演化分析中都属于相同的分支,但分离株样本信息中,未有类似美国PRRSV RFLP 1-4-4 Lineage1C变异株导致大规模发病的报道。其原因可能为:一方面,影响PRRSV毒力的相关因素受到病毒基因组中的多个基因所决定[13],而RFLP 1-4-4 L1C类毒株是基于ORF5序列进行分析,ORF5序列仅占PRRSV全基因组序列长度的4%,因此该方法在展示毒株遗传多样性上存在着一定的局限性,并不能反映出PRRSV全基因组遗传演变特征;另一方面,重组也是导致毒株多样性的原因,不同的重组位置、重组片段长度、重组模式均会产生不同的生物学特征。
综上,本研究基于64条类NADC30毒株全基因组序列进行分析,发现我国类NADC30毒株呈现复杂的Nsp2区域缺失模式和重组模式,丰富了类NADC30毒株的基因组信息。另外,目前尚不清楚美国PRRSV RFLP 1-4-4 Lineage1C变异株是否已传入我国猪群,应加强在肉类进口的检疫以及对国内猪群的流行病学监测,同时关于我国RFLP1-4-4 Lineage1C毒株的流行与变异情况仍需密切关注。
猜你喜欢
科学大观园(2022年2期)2022-01-23
今日农业(2021年11期)2021-08-13
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
动物医学进展(2015年10期)2015-12-07
百科知识(2015年18期)2015-09-10
特产研究(2014年4期)2014-04-10
郑州大学学报(理学版)(2014年3期)2014-03-01
遗传(2014年3期)2014-02-28
世界科学(2014年8期)2014-02-28
