
高温干旱背景下太湖藻情变化特征及机制*
2023-11-06 08:19康丽娟朱广伟朱梦圆国超旋张运林秦伯强
湖泊科学 2023年6期
康丽娟,朱广伟**,邹 伟,朱梦圆,国超旋,许 海,肖 曼,李 娜,张运林,秦伯强
(1:中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,南京 210008)(2:中国科学院大学,北京 100049)
在过去几十年,由水体富营养化引起的蓝藻水华问题在全球范围内呈现出不断扩张趋势,已成为淡水水体面临的重要生态问题之一[1-2]。湖泊蓝藻水华的发生不仅会造成水体浊度增加、缺氧,还会产生藻毒素,影响水生动植物的生存,严重威胁湖泊生态环境的健康与安全[3]。由于城镇化、人口的快速增长及污染排放,我国长江中下游流域的许多浅水湖泊都处于富营养化状态,深受蓝藻水华问题的困扰[4]。太湖是该流域典型的富营养化湖泊之一,从1980s末开始出现蓝藻水华问题并一直持续至今,经过长期的大规模治理,蓝藻水华问题依然严峻[5]。
氮和磷是藻类生长和增殖的关键限制元素[6-7],淡水湖泊中藻类常受到磷限制或氮和磷的共同限制[8-9]。在太湖的氮磷添加实验中,发现在夏季添加氮元素可以积极促进藻的生长,而磷则是限制藻在冬春季生长速率和生物量的主要因素[10]。一般来说,流域的外源输入是湖体营养盐的主要来源,在太湖,每年环太湖河道进入湖体的外源总磷负荷可达到2206.5 t[11],在洪水年份该负荷会更高。对于浅水富营养化湖泊来说,沉积物内源的释放也是湖体的一个重要磷来源[12]。较高的风速会通过增强颗粒物的再悬浮过程,促进水柱磷浓度的增高[13];在蓝藻水华暴发期间,藻细胞的衰亡也会促进磷从沉积物向水体的迁移释放[14],并维持藻类的继续扩张[15]。
营养盐与温度的相互作用是驱动浮游植物生长和群落组成最重要的两个因素[16]。营养盐的可利用性及浓度会影响细胞繁殖速率,同时改变藻类对温度的响应[17]。而温度会通过促进生物体酶和细胞的新陈代谢,改变藻类生物量与养分浓度的关系[17]。因此,高温环境通常会显著加强蓝藻的优势地位,增加湖泊中蓝藻水华的发生强度与范围[3,18-19]。变暖、强降雨、风速下降、干旱等极端天气通过改变湖泊的水文过程和营养盐动态,影响湖泊蓝藻水华的生长和扩张过程。如变暖显著延长了藻类的生长期,促使蓝藻水华的提前发生[20];强降雨在短期内将大量营养盐输送进入湖体,在雨期后易被藻类捕获并刺激水华的发生[3,21];研究表明在太湖夏季台风后,常常伴随着较大面积的蓝藻水华出现[21-22]。
在全球变化背景下,温度升高、风速下降、强降雨、干旱等极端天气的发生频率和范围的不断增加[23],增加了湖泊蓝藻水华发生的不确定性和控制的难度[18,24]。已有的研究表明,太湖的蓝藻水华在近5年处于较为强烈的波动状态,对气象水文因素的变化敏感[25]。2022年长江流域经历了自1961年有气象记录以来最严重的干旱事件,以及持续的高温[26-27],湖泊水位下降。干旱环境下,湖泊的入湖水量和外源负荷下降,水力滞留时间延长,影响湖体营养盐供给从而改变藻类水华态势。为探究干旱对湖泊蓝藻水华的影响,本研究利用太湖从2005以来的长期观测的营养盐和叶绿素a浓度数据,结合遥感影像反演的太湖蓝藻水华面积数据,旨在:(1)揭示高温干旱环境背景下太湖蓝藻的生长与分布特征;(2)阐明气象因素与营养盐共同作用对太湖藻情的影响机制;(3)提供蓝藻水华对气象环境的响应所带来的管理启示。
1 材料与方法
1.1 研究区概况
太湖地处我国长江下游平原,是我国第三大淡水湖泊(水域面积2338 km2),平均水深1.9 m。太湖流域河网复杂,水系众多,与太湖相连的环湖河道多达两百余条,其中入湖河道是太湖的主要水量和污染物负荷来源。由于流域内人口的增长、经济的快速发展和水资源的不合理开发,太湖自1980s开始富营养化并在北部的梅梁湾区域出现蓝藻水华,此后蓝藻水华逐渐扩张并在每年的春夏季频繁出现[28]。自2007年无锡饮用水危机事件暴发以后,太湖流域实施了一系列的外源营养盐削减和内源保护措施,包括污水处理厂的建设、污水管网改造、湖滨岸带的建设和蓝藻的打捞等[18],投资巨大,但蓝藻水华的改善甚微。2017年,太湖的蓝藻水华强度和水体总磷浓度双双反弹,给太湖蓝藻水华防控及周边城市饮用水安全保障带来了挑战[29]。
1.2 观测数据
太湖湖泊生态系统研究站(简称太湖站)自2005年以来在太湖布设的32个监测点位(图1),包括14个月度监测点位和18个季度监测点位(季度监测时间为每年的2、5、8、11月),进行氮磷浓度、叶绿素a(Chl.a)和藻类生物量等的观测分析。根据湖泊位置,32个监测点位分成梅梁湾、贡湖湾、胥口湾、东太湖、竺山湾、湖心区、西南湖区7个区域。在每个采样点,利用采水器分别采集表、中、底3层水样充分混合后获得水柱混合样品,并立即冷藏带回实验室。原水进行总氮、总磷浓度测定;收集经过0.45 μm孔径的玻璃纤维滤膜(Whatman GF/F)的定量水样和滤膜,滤膜用于提取Chl.a,滤后水用于溶解态总氮(DTN)、溶解态总磷(DTP)和无机态氮磷的测定。其中,Chl.a采用改良的热乙醇萃取—分光光度法测定。总氮、总磷浓度根据GB 11894-1989的方法测定。浮游植物的样品是通过加入1%的酸性鲁哥溶液进行固定(一般是采集1 L 水样加入10 mL的鲁哥溶液),带回实验室指定漏斗内静止沉淀48 h,浓缩至30 mL后在显微镜下对样品浮游植物的种类进行鉴定和计数。湖面表层水温(WT)、溶解氧(DO)、pH等参数利用便携式多参数水质分析仪(YSI, 6600V2)进行原位测定。
从2016年起,选取环太湖主要的16条环湖河道,进行河道氮磷浓度的逐月监测。采样的时间一般为每月中旬,和湖体监测时间相对应。河道采样点在靠近湖口的位置,能比较真实地反映河道入湖的营养盐浓度情况,河道样品的水质指标测定方法与湖体水质的测定方法一致。16条河道依次为望虞河、胥江、瓜泾、太浦河、大钱港、长兜港、长兴港、合溪港、大浦河、陈东港、沙塘港、殷村港、漕桥河和太滆运河、雅浦港、直湖港(图1)。其中望虞河、胥江、瓜泾、太浦河位于太湖东面,以出湖为主,其余河道多以入湖为主。
1.3 水文气象数据
气象数据来源于设置在太湖站的逐日水温、水位(吴淞高程)人工观测,于每日8、14、20时3次现场记录水温与水位的观测值,其均值为日均值,其中水温测点为水下0.5 m。降雨量与其他相关气象指标由设在太湖站的气象观测场自动气象监测系统获得。
1.4 水华面积
本文采用的太湖蓝藻水华面积的遥感反演方法见文献[30-31],选用美国NASA系统(GSFC, http://oceancolor.gsfc.nasa.gov/) 提供的MODIS-Aqua逐日影像数据。文中选取了从2017年1月-2022年12月间的共803景太湖影像经SAEDAS软件进行辐射定标、几何校正、大气校正以及人工目视筛选得到无云高质量影像作为数据源[30]。由于藻华水体在近红外波段反射率较高,而非藻华水体在近红外和短波红外的吸收较强,在MODIS影像波段下,以645、1240 nm波段为两端基点,以859 nm波段为中心波长,可以对藻华水体与非藻华水体进行辨识[31];根据此特点,基于Hu等提出的浮藻指数(floating algae index,FAI)以及水华判别阈值(FAI>-0.004)[31],对逐日的水华面积进行提取和统计。
1.5 统计分析
基于ArcGIS软件中的克里金插值法获得太湖2005-2022年营养盐浓度的空间分布,并计算了营养盐各浓度梯度的空间占比面积。根据吴雅丽等对太湖春季藻生长的磷培养实验结果,春季藻类生长的无机磷阈值为0.02 mg/L(对应的总磷阈值为0.059 mg/L)[32];总氮的阈值为0.8 mg/L[33],考虑到野外环境中藻类对水体磷的利用以及氮的季节性变化,以冬季溶解态氮磷浓度衡量湖泊中生物可利用的营养负荷[34],本文统计了冬季DTN和DTP在空间插值中分别小于0.80和0.02 mg/L的湖区面积。样本的相关性通过Pearson相关分析进行检验。基于湖区计算了每个湖区的浮游植物和微囊藻生物量均值,并对浮游植物和微囊藻生物量数据进行对数运算后以满足正态分布。
2 结果与分析
2.1 太湖2022年的气象和水文特征
2.1.1 气象因素 (1)降雨。2022年太湖累积降雨量为927.4 mm,较2005-2021年长期均值(1085.8 mm)下降了14.6%。其中汛期(5-9月)累积降雨量为401.1 mm(表1),较长期均值减少了42.5%,遭受了严重干旱;其中以5月降雨量最少,月累积降雨量仅为10.16 mm,是历史同期均值的11%。7月和8月累积降雨量分别为77.0和41.7 mm,低于历史同期均值的58.2%和63.9%。(2)气温。太湖2022年平均气温为18.0℃,高于历史同期的气温均值(17.3℃)。(3)光强。2022年光合有效辐射年均值为28.26 mol/(m2·d),高于历史年均值(25.41 mol/(m2·d))。其中5-9月的光合有效辐射明显高于历史均值。(4)风速。在近年全球风速下降的环境下,太湖2022年的风速也呈现出进一步降低趋势,全年平均风速为1.95 m/s,其中春季平均风速为2.09 m/s,8月份平均风速最低,为1.65 m/s;其次是5月份,平均风速为1.79 m/s。

表1 2022年降雨量与降雨天数Tab.1 Rainfall and rainfall days in 2022
2.1.2 水文因素 (1)水位。与降雨量相对应的,从4月开始水位陆续下降(图2),2022年汛期平均水位为3.11 m,显著低于同期历史均值水位(3.35 m)。8月水位均值仅为3.00 m,为2005年以来最低均值。(2)水温。太湖2022年平均水温为20.4℃,高于历史同期水温均值(19.0℃)。其中春季(3-5月)水温19.8℃,较2021年(19.6℃)和2020年(19.7℃)变化不大;太湖夏季(6-8月)平均水温达32.3℃,超过历史(2005-2022年)同期的10.2%;水温超过30℃的天数达30天。
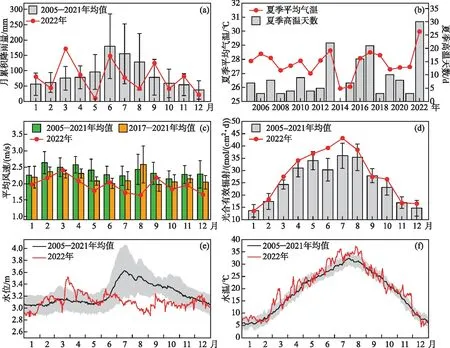
图2 2022年太湖流域的气象和水文特征:(a)月累积降雨量;(b)夏季平均气温;(c)平均风速变化;(d)光合有效辐射;(e)日水位变化;(f)日水温变化Fig.2 Characteristics of meteorological and hydrological in Lake Taihu Basin in 2022: (a) monthly cumulative rainfall; (b) average air temperature in summer (from June to August); (c) monthly average wind speed; (d) monthly mean of daily photosynthetically active radiation; (e) daily water levels; (f) daily water temperature
2.2 2022年的太湖藻情
2.2.1 蓝藻水华面积与叶绿素a浓度 从遥感获取的蓝藻水华面积结果来看,在水华高发季(5-9月),2022年蓝藻水华的平均范围和最大面积较近5年相比均有明显的下降趋势,水华的平均面积为 189 km2,观测到最大面积为528.9 km2。其中,5月份的水华面积为91.3 km2,明显偏小,仅为近5年同期水华面积均值(454.6 km2)的1/5;全年来看,10月份的平均水华面积最大,达280.4 km2,最大面积也是出现在10月。2022年水华季节超过500 km2的水华次数比例为3.6%,明显低于近5年平均比例(19.8%)。从北太湖的Chl.a变化来看,2022年为52.6 μg/L,是2017年以来的最低值,Chl.a最高值出现在8月,为79.8 μg/L。其中5-7月均明显低于近5年平均浓度,5月的Chl.a均值为29.7 μg/L,较近5年同期(92.8 μg/L)下降68.0%(图3)。
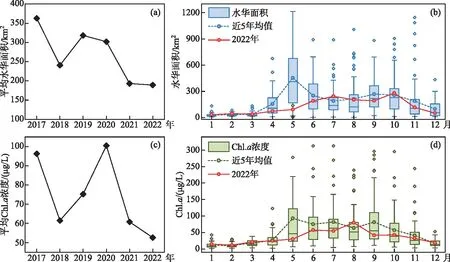
图3 2022年蓝藻水华面积与Chl.a浓度变化:(a)2017-2022年水华季节(5-9月)太湖平均水华面积的逐年变化;(b)2022年太湖逐月水华面积;(c)2017-2022年水华季节(5-9月)北太湖平均Chl.a浓度的逐年变化;(d)2022年北太湖逐月Chl.a浓度变化Fig.3 Variation of cyanobacterial bloom area and chlorophyll-a concentration in 2022:(a) average annual bloom area of Lake Taihu in bloom season (May to September) from 2017 to 2022; (b) monthly bloom area of Lake Taihu in 2022; (c) average annual chlorophyll-a concentration in North Lake Taihu during the bloom season from 2017 to 2022; (d) monthly chlorophyll-a concentration in North Lake Taihu in 2022
2.2.2 微囊藻生物量 2022年春季的总藻生物量(Ba)为2.38 mg/L,同样是处于近5年的最低值。微囊藻(Microcystisspp.)是太湖春夏季蓝藻的最主要优势种群之一,其生物量(Bm)在相当程度上指示了蓝藻水华的强度。2022年春季的微囊藻生物量明显下降,为0.32 mg/L,为近10年来的最低值,且微囊藻在总藻生物量中的占比明显下降,由近5年的70.1%下降到13.3%(图4)。
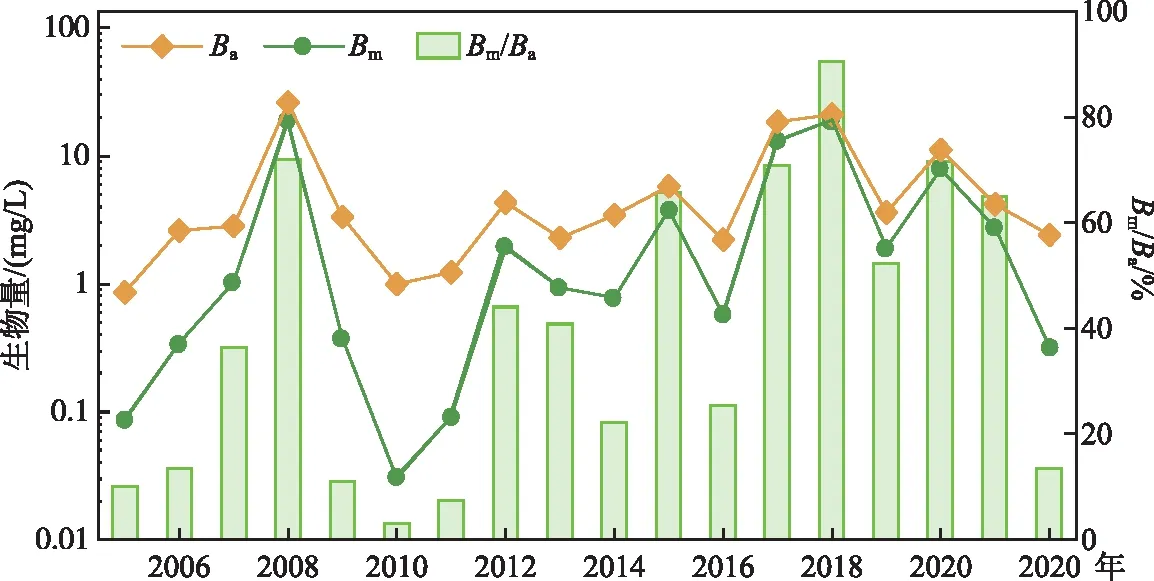
图4 2005-2022年太湖春季(5月)总藻生物量(Ba)和微囊藻生物量(Bm)的变化Fig.4 Variable of total algae biomass (Ba) and Microcystis algae biomass (Bm) in May in Lake Taihu from 2005 to 2022
2.3 氮磷浓度的长期变化特征
2.3.1 太湖氮磷浓度的时间变化 2005-2022年,太湖TN浓度呈现明显下降趋势,TP浓度呈现出波动下降趋势,其中以冬春季下降最为明显。在2022年,全湖TN和TP年均值分别为1.41和0.084 mg/L,均为2005年以来的最低值,较近5年均值(2017-2021年)分别下降了30.6%和27.3%。溶解态氮磷作为藻类生长可直接利用的养分物质来源,也与总氮、总磷表现出相似的变化趋势,其中冬春季的DTP在2010年后呈现出较为稳定的下降趋势,夏季的DTP在2016-2017年前后出现反弹后也呈现出下降趋势。2022年全湖的DTN和DTP年均值分别为0.83和0.028 mg/L,冬季(2月)的DTN和DTP浓度均为2005年以来的同期最低值,分别为1.36和0.017 mg/L(图5)。
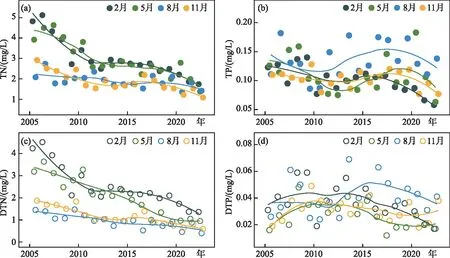
图5 2005-2022年太湖TN、TP、DTN和DTP浓度的逐季节变化趋势(其中趋势线由局部加权回归拟合)Fig.5 Seasonal variation of total nitrogen, total phosphorus, total dissolved nitrogen and total dissolved phosphorus concentrations in Lake Taihu during 2005-2022
2.3.2 太湖溶解态氮磷浓度的空间分布 空间插值结果表明,在2022年冬季(2月),全湖除西北竺山湾湖区外,大部分湖区的DTP均低于0.020 mg/L,总面积占比达到79%,是为2010年以来的最大范围。DTN小于0.80 mg/L 的湖区面积也在近2年明显增加,2022年冬季,DTN较高的湖区主要分布在西北和西南沿岸区,其中DTN低于0.80 mg/L 的面积占比达到22%(图6)。
2.4 微囊藻生物量的影响因素
利用随机森林量化主要环境因素对微囊藻和总藻生物量影响的相对重要性(图7)。分别选取2005-2022年春夏季水温(Temp)、累积降雨(RF)、平均风速(AWS)、水位(WL)、光强(PAR)、总氮(TN)、总磷(TP)以及冬季溶解性氮磷的进行分析(氮磷的下标表示月份,水温是对应的春夏季均值)。结果表明:春季的TP5、Temp、AWS是影响微囊藻和总藻生物量最主要的因素,与DTP5共同解释了48.8%的春季微囊藻生物量变化。TP5和DTP2与总藻类生物量的相关性系数分别为0.47和0.36;TP5、Temp、DTP2和AWS共同解释了56.7%的总藻生物量变化。
在8月份的夏季,与微囊藻生物量相关性最高的是TP和TN浓度,相关系数分别为0.54和0.50,其中8月份的TP8、TN8、2月份的DTN2以及WL的变化解释了48.4%的夏季微囊藻生物量变化;8月的TP8、TN8也与总藻生物量表现出较高的相关性(图7)。
3 讨论
3.1 太湖2022年藻情偏轻的主要驱动因素
3.1.1 春季低磷浓度对微囊藻生物量的影响 根据随机森林分析结果的重要性排序,春季的总磷、水温和风速是影响太湖春季微囊藻时空变化的重要因素。通过对2005-2021年间微囊藻与总磷、水温和风速之间的对比分析可以看出,总磷浓度与温度和微囊藻生物量之间有较好的正向关系(线性回归:lg(Bm)=5.25+5.79lg(TP),R2=0.54)。太湖梅梁湾的营养盐添加实验发现,春季无机磷浓度在0.02 mg/L 时(对应TP浓度为0.059 mg/L),藻类生长速率达到最大值,低于此值时藻类的生长速率与生物量会受到磷浓度制约,并且随磷浓度的增加而显著增加[32]。2022年冬季太湖DTP浓度为0.017 mg/L,DTP小于0.02 mg/L的湖区范围达79%,即相当一部分的湖区的藻类生长速率受到磷的限制;春季TP浓度为0.063 mg/L,也明显低于往年同期值,低磷浓度是造成春季微囊藻生物量较低的主要原因。
此外,影响微囊藻生物量的另外2个重要因素——水温和平均风速,分别与微囊藻生物量呈现出正向和负向关系(图8)。其中水温的增加有利于蓝藻的快速生长与代谢[3-4,17],而低风速有利于浅水湖泊的水下光环境的改善,并促进藻颗粒在水中的垂直分层。在强光照和低风速的环境下,蓝藻可以利用细胞结构中的伪空胞上浮到表层获得充足的光照,并获得优势生态位[35]。已有的研究表明,太湖风速的下降有利于藻类的生长与积累[36-37],促进藻类的“光生态位”提升和对营养物质的利用效率[38]。2022年春季,水温为19.8℃,风速为2.09 m/s,均在一定程度上有利于微囊藻生物量的增加,而低磷浓度限制了藻类对水温以及风速可能出现的正向响应(图8)。
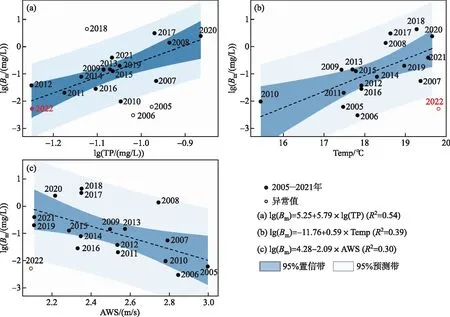
图8 太湖春季微囊藻生物量(Bm)与主要驱动因素间的关系:(a)微囊藻生物量与春季总磷的线性回归关系;(b)微囊藻生物量与春季水温的线性回归关系;(c)微囊藻生物量与平均风速(AWS)的线性回归关系Fig.8 Relationship between major drivers and Microcystis biomass (Bm) in spring in Lake Taihu: (a) linear regression relationship between Microcystis biomass (Bm) and total phosphorus (TP) in spring; (b) linear regression relationship between Microcystis biomass (Bm) and water temperature (Temp) in spring; (c) linear regression relationship between Microcystis biomass and average wind speed (AWS) in spring
3.1.2 夏季微囊藻生物量对营养盐的响应 2022年夏季太湖流域经历了长期的高温。一般来说,高温能显著促进蓝藻的生长优势[3,18]。遥感水华的面积监测结果表明,太湖8月的蓝藻水华面积较5月有明显增加,但与多年夏季平均值基本持平。相关分析和随机森林的结果也表明,在2022年8月,水体的TP和TN浓度与微囊藻和藻类的生物量变化密切相关。微囊藻监测结果中,以草型生态系统为主的东太湖和胥口湾未出现微囊藻,其余点位微囊藻生物量与TP浓度在空间上也呈现出显著的正相关关系(图9,拟合方程 lg(Bm)=2.55×ln(lg(TP)+1.82),R2=0.73)。在西北的竺山湾以及部分西南湖区点位,微囊藻生物量较高;这与入湖区沉积物的内源释放有关。竺山湖等藻型区,沉积物中含有较高的活性磷[39],高温和藻华过程会共同刺激活性磷从沉积物向水体的迁移释放,补给藻生长所需要的磷[14,40-41]。而在湖心区和贡湖湾,沉积物中的活性磷较低,前期藻类生物量较低,藻类对沉积物内源磷释放的“泵吸”作用较弱[14];加上干旱少雨,外源入流补给的营养盐主要停留在西部湖区,难以向东推移,导致水体总磷浓度维持较低浓度,限制了微囊藻的过度增殖。总氮浓度也呈现出类似的空间分布,在湖心区、贡湖湾以及梅梁湾湖区处于较低水平,而在竺山湾和部分西南湖区点位较高,与微囊藻生物量的分布一致。
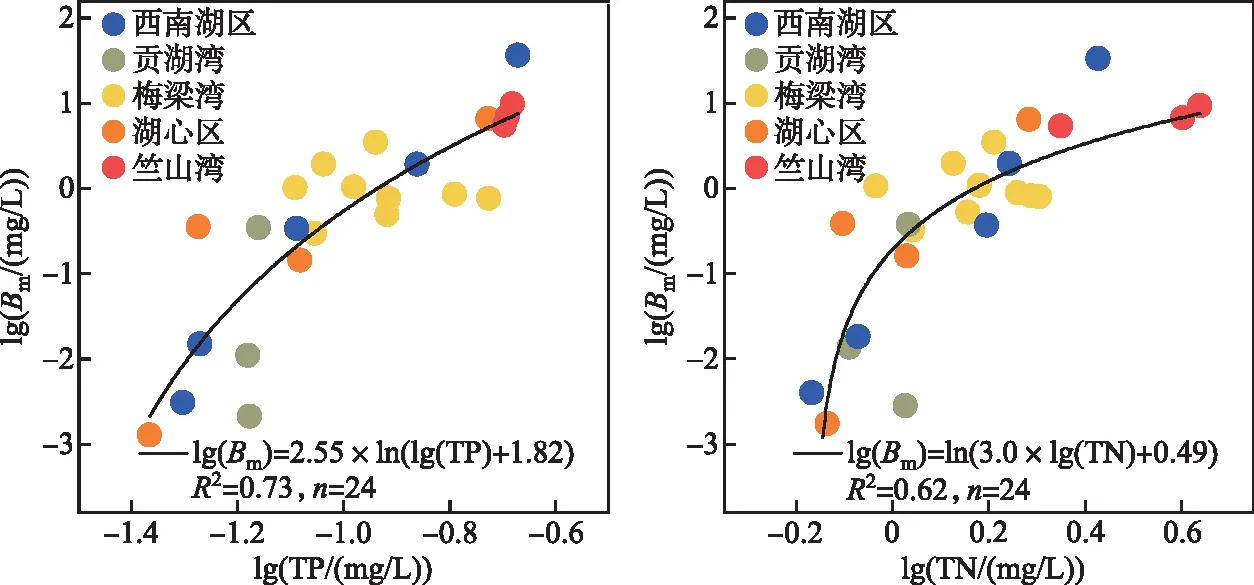
图9 太湖夏季不同点位TP和TN与微囊藻生物量(Bm)的关系Fig.9 Relationship between total phosphorus and Microcystis biomass (Bm), as well as between total nitrogen and Microcystis biomass (Bm), at different sampling sites in Lake Taihu during the summer
3.2 局部湖区蓝藻生长受到营养盐限制的原因
3.2.1 营养盐浓度的持续总体下降 湖体的氮磷变化通常具有连续性,春季水体的氮磷浓度受到冬季溶解态氮磷浓度的影响。随机森林结果表明,春季和夏季的微囊藻生物量均在一定程度上受冬季溶解态氮磷的影响,冬季溶解态氮磷浓度是湖泊中生物可利用的重要营养部分[34]。通过对冬季溶解态氮磷与春夏季总磷浓度的相关分析发现,DTP2与TP5和DTP5之间具有较高的相关性,相关系数分别为0.71(P<0.01)和0.83(P<0.01);DTN2也与TN5和DTN5之间具有高的相关性,相关系数分别为0.86(P<0.01)和0.80(P<0.01)(表2)。这表明春季湖体的TN和TP浓度受到冬季湖体的DTN和DTP浓度很大的影响。2022年冬季,太湖的DTN和DTP较低,其中DTP低于0.02 mg/L的范围占到全湖的79%,DTN低于0.80 mg/L 的范围达到22%,这影响了春季湖内的氮磷浓度。
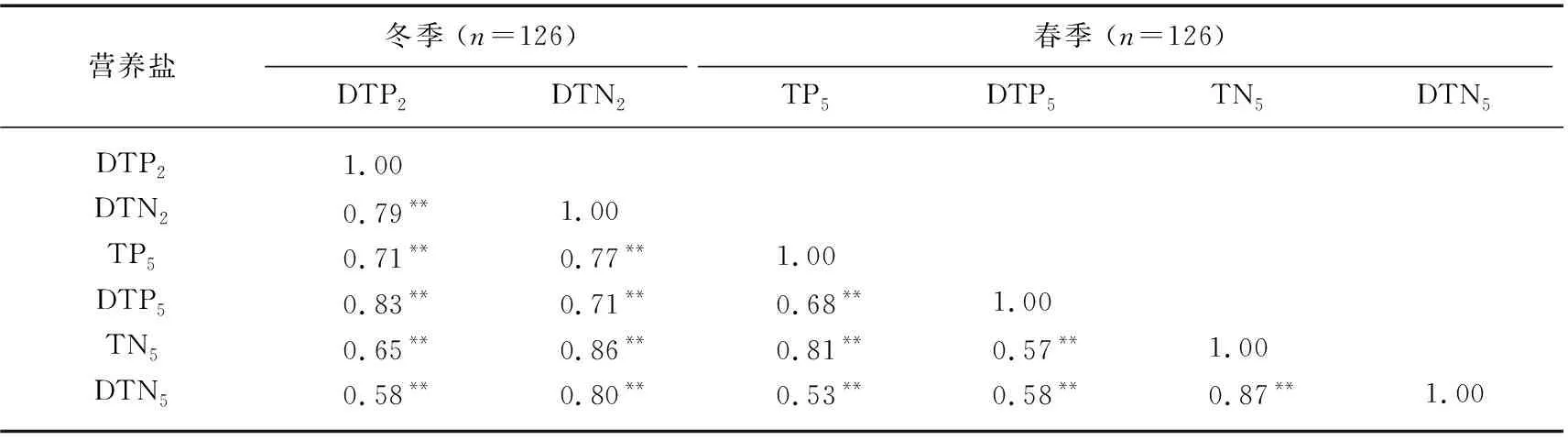
表2 太湖春季和冬季营养盐浓度的相关性分析结果Tab.2 Correlation analysis of nutrient concentration between spring and winter in Lake Taihu
3.2.2 干旱降低了春夏季的外源营养盐补给 冬春季的外源负荷对生长期湖内的藻类生物量至关重要。在伊利湖,春季入湖的有效磷负荷被用于定量预测夏季蓝藻水华强度[42]。对于太湖而言,河道输入的TP负荷是水体的主要外源磷来源。TP负荷受到入湖水量的影响很大[11],后者主要是由降雨量控制[43-44]。根据降雨量估算的环太湖入湖水量,以及2016-2022年太湖主要环湖河道水质调查,获得了2016-2022年环湖河道的入湖负荷(图10)。结果表明,2022年春季(3-5月)TN和TP的入湖负荷分别为8077.8和273.7 t,较2016-2021年均值分别下降了18.0%和35.2%;汛期(5-9月)TP和TN负荷整体下降了57.5%和60.3%。其中5月受到水量的限制,入湖的TP和TN负荷仅为211.7和11.7 t,分别下降了93%和94%,不足常年的1/10。入湖负荷的下降进一步维持了全湖较大湖区范围内的低氮磷浓度状况,从而对藻类的生长产生了限制效应。
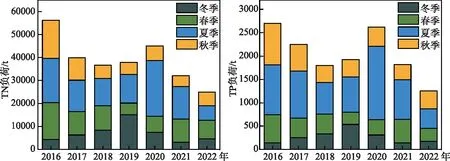
图10 2016-2022年太湖入湖河道的氮磷负荷Fig.10 External input total nitrogen and total phosphorus loads of Lake Taihu from 2016 to 2022
相类似地,2010年和2011年也先后经历了较为严重的干旱,2010年累计降雨量776 mm,低于均值的28%;其中夏季降雨量仅198 mm,而2011年主要为春季干旱,3-5月累计降雨量110 mm,低于同期均值的55%。相对应地,2010年和2011年5月TP浓度分别为0.098和0.077 mg/L,处于较低水平(图5),2010年和2011年春季的藻类生物量分别为0.99和1.23 mg/L,微囊藻生物量分别为0.03和0.09 mg/L,均表现出明显下降(图4)。因此,对于太湖这种对外源营养盐负荷依赖性较高的浅水湖泊而言,春季的干旱少降雨会影响到湖内的磷的动态,从而影响春季的藻情变化,尤其是微囊藻生物量的变化。
3.2.3 低风速可能减少了沉积物营养盐的动态补给 风力诱导的湖泊沉积物再悬浮过程是浅水湖泊沉积物营养盐向水中动态补给的重要过程[45]。研究表明,在台风期间,水柱中悬浮颗粒物浓度和总磷浓度会随着风速增加而显著上升[46]。秦伯强等对太湖的观测结果表明,导致沉积物悬浮所需的临界切应力大致相当于风速4 m/s时产生的底泥扰动力;当风速达6.5 m/s以上时,太湖沉积物将发生大规模悬浮,同时将间隙水中的溶解性营养盐快速释放[47];李一平等的结果表明在梅梁湾较浅的水域,风速为3 m/s时,对应的临界切应力就可使梅梁湾底泥起悬[48]。分析太湖春季的风速与水体悬浮物浓度的变化可以发现(图11),太湖春季平均风速与水体平均悬浮颗粒物浓度间具有一致的变化趋势(相关系数R=0.69)。在2022年,风速继续表现出下降趋势,尤其5月的平均风速(均值为1.79 m/s)为2005年来同期最低值,水体悬浮物浓度仅为29.6 mg/L,表明水体的再悬浮过程较弱,一定程度上降低了底泥悬浮引起的内源营养盐补给,维持了湖泊水柱中的营养盐的较低状态。尽管低风速有利于湖泊光环境的改善[37],但低风速在一定程度上也减少了沉积物对水体的营养盐补给过程,在前期水体微囊藻生物量较低的背景下,低营养盐水平对藻情的影响可能是主导性的。
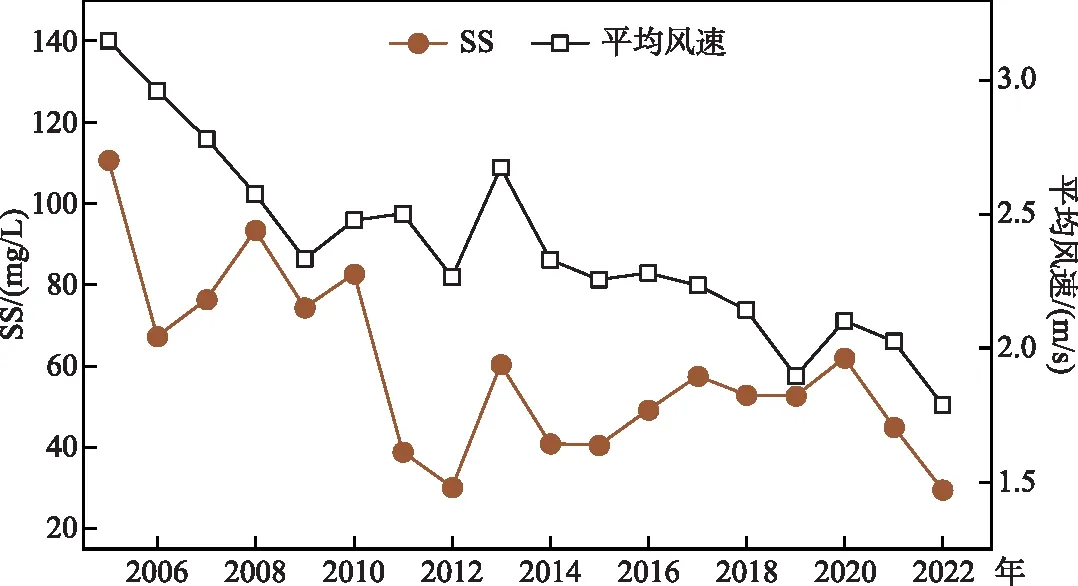
图11 2005-2022年春季平均风速与SS的变化Fig.11 Average annual wind speed and suspended particulate matter in spring from 2005 to 2022
3.3 干旱导致太湖营养盐浓度降低的管理启示
干旱通过改变湖泊营养盐的动态进而对湖泊的藻情变化产生影响。本研究表明,从2021年开始,太湖的氮磷低值区开始增大,2022年,79%的湖区冬季DTP低于0.02 mg/L;加上春季入湖负荷的下降,使得5月份全湖的TP为2005年来最低值,藻类的生长受到了抑制。在2021年,冬季全湖的溶解态氮磷也处于较低状态,但由于降雨较多,2021年春季和夏季TP入湖负荷分别是2022年的1.85和2.04倍,为藻类的生长起到了及时的补给作用,因此春季的氮磷浓度高于2022年(图5)。而2022年的连续干旱背景下,汛期外源TN和TP负荷分别下降了57.5%和60.3%,且在5月入湖负荷仅为常年的10%,导致除了竺山湾等入湖口湖区外,较大的湖区的养分维持在较低水平,不仅对春季蓝藻的生长起到了明显的抑制作用,并且在极其高温的夏季,太湖蓝藻水华的面积没有显著高于往年。随机森林结果表明总磷和总氮是最重要的因素,微囊藻生物量与总氮、总磷的浓度在空间上具有较高的一致性,表明大范围湖区的低营养盐浓度对藻类的生物量起到了限制效应。而降雨等气象因素对藻类生物量有着较大的影响;春夏季的外源补给对藻类生长季的范围与强度起到决定性的作用。建议对蓝藻水华的监测可以从冬季(2月)开始,关注水体中易被藻类利用的溶解态氮磷浓度,同时关注其在空间上的分布,从而将蓝藻水华控制在一定湖区范围内不扩张。
营养盐控制是蓝藻水华防控的根本途径。2022年的干旱天气通过减少入湖水量极大的削减了的外源的氮磷负荷,再次验证了外源削减对蓝藻控制的根本性与可行性。在气候变化的环境下,温度的升高与风速的降低,都在很大程度上支持了蓝藻水华向更适宜扩展的方向发展。浮游植物物种多样性的下降同样表明太湖微囊藻等优势藻种对营养盐的利用效率呈现出上升的趋势[49]。因此,氮磷的控制可能需要严格的阈值来应对这种环境的变化以及未来高温气候等带来的蓝藻水华风险。
最后,健康稳定的水生态系统离不开健康的水生态结构和稳定的食物网。沉水植被的修复有利于提升水体透明度、削减风浪、抑制水体沉积物的再悬浮和沉积物的内源释放等过程,并为浮游动物、底栖动物以及鱼类等提供适宜的栖息地。从根本上进一步降低蓝藻水华强度,还需要进一步调整湖泊鱼类群落结构,通过下行效应提高对浮游植物生物量的控制能力。结构不合理的食物链可能会对藻类生物量的控制起到负面作用,如有研究表明底栖和浮游动物食性鱼类生物量增加,不利于藻类生物量的控制[50]。
4 结论
1)在2022年严重高温干旱的气象背景下,太湖蓝藻水华面积的平均范围及最大面积与近5年相比处于较低状态,其中5月份水华面积、叶绿素a浓度、微囊藻生物量均明显下降。
2)太湖氮磷浓度近年来保持下降趋势,2022年氮磷浓度降至历史低值;加上干旱引起的外源负荷下降,较大范围湖区的磷浓度维持在较低水平,是2022年太湖的春季藻类生物量较低的主要原因。较低的外源负荷一定程度上也缓冲了夏季高温可能造成的水华大面积反弹现象。
3)气候因素对湖泊蓝藻水华态势的影响不可忽视。2022年极端干旱环境下的太湖藻情变化,再次表明了外源负荷对春夏季藻情的重要影响。在此背景下,严格的外源控制是必要且有效的。此外,还需警惕干旱后的强降雨对河道蓄积的污染物的冲刷而对湖泊水质可能造成的负面影响。
致谢:感谢中国科学院太湖湖泊生态系统研究站监测队伍提供的相关数据;感谢太湖站蓝藻水华预测预警课题组提供的分析思路!
猜你喜欢
当代水产(2021年8期)2021-11-04
当代水产(2019年8期)2019-10-12
当代水产(2019年9期)2019-10-08
水利技术监督(2017年6期)2017-12-19
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
西北园艺(果树)(2015年1期)2015-02-21
癌变·畸变·突变(2014年2期)2014-03-01
湖南农业科学(2014年18期)2014-02-27
华东理工大学学报(自然科学版)(2014年3期)2014-02-27
