
环境因子及生态补水对乌梁素海“黄苔”年际暴发面积的影响(1986-2021年)*
2023-11-06 08:19杨瑾晟罗菊花霍轶珍兰海今谷孝鸿史小丽
湖泊科学 2023年6期
芦 津,杨瑾晟,罗菊花,张 民,阳 振,霍轶珍,兰海今,谷孝鸿,史小丽,3**
(1:中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008)(2:中国科学院大学,北京 100049)(3:中国科学院大学南京学院,南京 211135)(4:内蒙古河套学院生态与资源工程系,巴彦淖尔 015000)(5:巴彦淖尔市乌梁素海生态保护中心,巴彦淖尔 015000)
在全球气候变化和人类活动影响加剧背景下,全球湖泊发生了不同程度的富营养化,湖泊藻类水华频次也相应增加[1]。不同于长江中下游的太湖、巢湖等典型亚热带浅水湖泊的蓝藻水华[2],附着藻类(Epiphyton)异常增殖形成的“黄苔”(Metaphyton)暴发是我国北方浅水湖泊面临的重大水生态环境问题。乌梁素海是我国北方湖泊“黄苔”暴发最典型、最严重的案例之一。“黄苔”主要由绿藻门中转板藻、水棉等丝状藻组成,幼年时期以着生藻的形式附着于沉水植物上生长,随着孢子成熟,逐渐聚集成较大团块漂浮至水面,经阳光照射后由绿色转为黄色,进而形成“黄苔”[3-4]。“黄苔”暴发会大量消耗水中氧气,降低水下光强,威胁水生动植物生存,尤其会导致沉水植被消亡、湖泊生态功能退化,进而逐步打破湖泊“草型清水”稳态,引发浮游藻类水华[5-7];同时大量植物残体腐烂还会加剧湖泊沼泽化进程[8]。“黄苔”暴发已对乌梁素海生态栖息地和渔业资源造成严重破坏,水生态环境安全和健康受到严峻挑战。
附着藻类生长与“黄苔”暴发受到诸多环境因子的影响[9]。首先,氮、磷营养盐通常被认为是附着藻类异常增殖的主要驱动因素,但乌梁素海“黄苔”和营养盐的关系还不太明确。例如,乌梁素海“黄苔”年内生消过程[10]和“黄苔”预警研究均未形成营养盐与“黄苔”关系的定论[11],而2000-2010年乌梁素海“黄苔”多年月度变化特征分析的结果表明“黄苔”与氮磷营养盐呈负相关[12]。温度升高会促进附着藻生长和增加“黄苔”生物量[13-14];风速亦被认为是春季乌梁素海“黄苔”生长的影响因子[15]。同时,附着藻生长还依赖附着基质和光照条件。草型湖泊通常透明度较高,沉水植物还能为附着藻提供附着基质[16]。特别是冠层型沉水植物茎或分枝长,植冠高大,绝大部分生物量被托举到水面附近,对光和空间竞争能力强,细碎叶片和多分枝结构也更有利于丝状藻的附着[17-18]。此外,流速[9]、水位波动[19]等水文因子也会影响附着藻的生长,但这些因素对于“黄苔”暴发的影响尚缺少相关研究。此外,为缓解乌梁素海沼泽化等环境问题,内蒙古自治区于2008年起实施生态补水工程,而由此引起的水环境变化对“黄苔”的影响还缺少科学认识和评价。
因此,本研究主要通过资料收集和卫星图像遥感解译,获得乌梁素海近35年(1986-2021年)湖体水环境质量、基本水文参数、气候气象因子、沉水植被和挺水植被面积数据,以及“黄苔”暴发年最大面积等长尺度数据,追溯乌梁素海“黄苔”暴发历程,揭示乌梁素海水环境的长时序变化特征和“黄苔”暴发的成因与规律,明晰生态补水工程对“黄苔”暴发的影响,并提出相应防控建议,以期为乌梁素海管理与保护提供科学依据。
1 材料与方法
1.1 研究区域概况
乌梁素海(40°36′~41°03′N,108°43′~108°57′E),位于我国内蒙古自治区巴彦淖尔市乌拉特前旗,河套地区灌溉系统的末端,是我国第八大、黄河流域第二大淡水湖泊,黄河流域以及内蒙古西部地区最重要的淡水资源。乌梁素海处于温带季风性气候带,年平均气温7.4~8.8℃,夏季降水占全年63.2%,年平均蒸发量和植物蒸散损耗达4.02亿m3,是全球半荒漠化地区极为罕见的大型草原湖泊。作为河套平原唯一的承泄水体,乌梁素海接纳了来自河套地区的农田灌溉水、生活污水、工业废水等污水。历史上,乌梁素海湖体不断萎缩,湖泊水域面积从1949年建国初期的799.2 km2缩减至近年的293.04 km2,水生植被泛滥,沼泽化进程加剧。为缓解现状,内蒙古自治区自2008年起实施乌梁素海生态补水工程,2008-2013年均补水量0.569亿m3,2014年后通过总排干大规模实施生态补水,年均补水量超过3亿m3。
1.2 数据获取与处理
“黄苔”生长过程在年内具有明显的周期性和持续累积的特性,且其引发的生态灾害基本建立在大规模暴发的情境下,因此本研究基于空间分辨率为30 m的Landsat TM/OLI系列卫星影像,参照相关方法[10]提取了1986-2021年“黄苔”面积变化的波峰段(暴发期)数据,以表征当年“黄苔”暴发的严重程度。同时,还参照相关方法[20]提取了历年沉水植物和挺水植物面积数据。
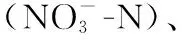

图1 乌梁素海监测样点分布Fig.1 Distribution of sampling sites in Lake Ulansuhai
1.3 数据分析
在R中使用“lm”函数分析乌梁素海环境因子的长时序变化规律,并对历年“黄苔”暴发面积的时间序列进行分段拟合;使用“cor.test”函数分析乌梁素海“黄苔”暴发面积与环境因子、气象因子和水文因子的相关性,筛选乌梁素海“黄苔”暴发的年际影响影子,并使用“lm”函数进行关系拟合,且在SPSS中进行多元线性回归分析。在R中使用“aov”和“LSD.test”函数对“黄苔”大规模暴发前后和实施生态补水工程时期主要影响因子的差异进行检验。使用“ggplot2”包对上述分析结果可视化;在所有分析中,P<0.05认为在统计学上具有显著差异。
2 结果与分析
2.1 乌梁素海水质长时序变化规律
乌梁素海水体CODCr在2013年之前处于36.3~113.9 mg/L高位波动,均值达62.8 mg/L,高于《地表水环境质量标准》(GB 3838-2002)中规定的Ⅴ类水标准(40 mg/L),为劣Ⅴ类,自2014年起稳定达标((32.7±3.9) mg/L)。1998-2021年,乌梁素海水体中TN、TP呈显著下降趋势(P<0.05)。其中,TN在2017年前浓度为1.2~11.0 mg/L,均值4.3 mg/L,基本为劣V类(>2.0 mg/L);TP在2008年由原先的劣Ⅴ类(0.09~0.41 mg/L,均值0.24 mg/L)提升至V类(0.1~0.2 mg/L),并在2014年后稳定低于0.1 mg/L。DO浓度则在2008年降至最低值(3.3 mg/L),实施生态补水工程后跃升至8.4 mg/L(2009年),并自2011年起维持在6.2~7.4 mg/L,基本反映出乌梁素海水体污染在生态补水之前不断加重、生态补水工程实施后逐步减轻的变化历程(图2)。
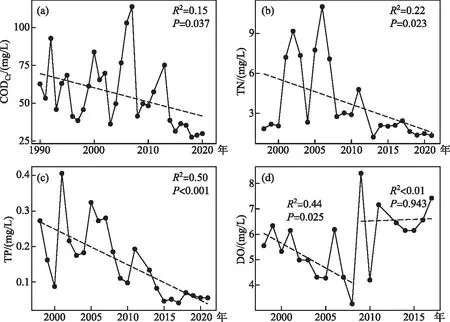
图2 乌梁素海1990-2021年(a)CODCr、(b)TN、(c)TP和(d)DO变化Fig.2 The changes of (a) CODCr, (b) TN, (c) TP, and (d)DO in Lake Ulansuhai between 1990 and 2021
然而,根据生态环境部2004年发布的《湖泊(水库)富营养化评价方法及分级技术规定》,基于TN、TP对湖体2019-2021年的综合营养状态指数(TLI(Σ))的计算结果为54.33、54.15、53.24,即乌梁素海仍处于富营养水平(TLI(Σ)>50)。因此,乌梁素海水体污染虽然在近35年呈显著下降趋势,但总体形势并不乐观。
2.2 乌梁素海“黄苔”暴发年际变化及其影响因子
近35年(1986-2021年)的历史数据显示,乌梁素海1986-1989年间“黄苔”面积均为0,直至1990年才出现“黄苔”暴发,面积为3.01 km2,至1997年基本保持此规模(多年均值为3.20 km2);1998年起,乌梁素海“黄苔”开始大规模暴发,面积达到 16.11 km2,此后一直保持高位暴发态势(1999-2008年多年均值为16.39 km2),并在2001和2008年达到最高,分别达到28.82和27.51 km2。线性拟合结果表明,2008年之前乌梁素海“黄苔”暴发面积随年份极显著增加(P<0.001)。而2008年末实施生态补水工程后,“黄苔”暴发的扩张趋势才得以抑制,2009-2021年间面积基本保持在10 km2以下((8.82±2.82)km2)(图3)。
图3 乌梁素海1986-2021年“黄苔”暴发面积变化Fig.3 The changes of the area of the metaphytic blooms in Lake Ulansuhai between 1986 and 2021
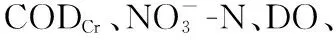
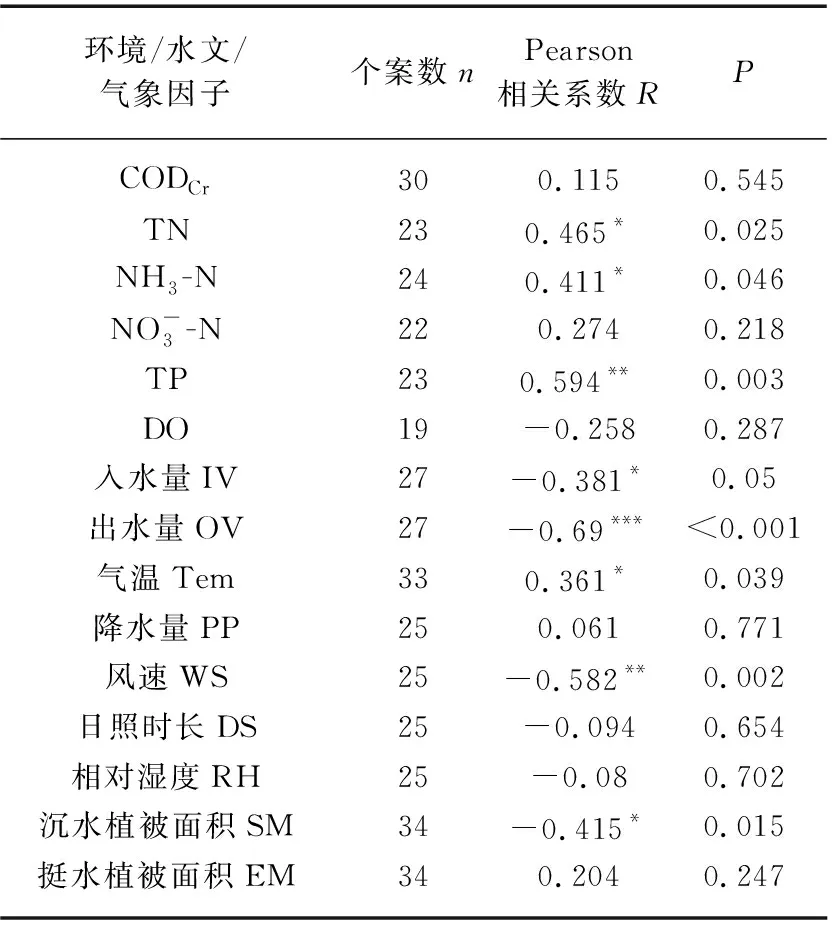
表1 乌梁素海“黄苔”暴发面积与环境、 水文和气象因子的相关性分析结果Tab.1 The results of correlation analysis between the area of metaphytic blooms and environmental, hydrological and meteorological factors in Lake Ulansuhai

表2 乌梁素海“黄苔”暴发多元回归模型结果*Tab.2 The results of multiple regression model for the area of metaphytic blooms in Lake Ulansuhai
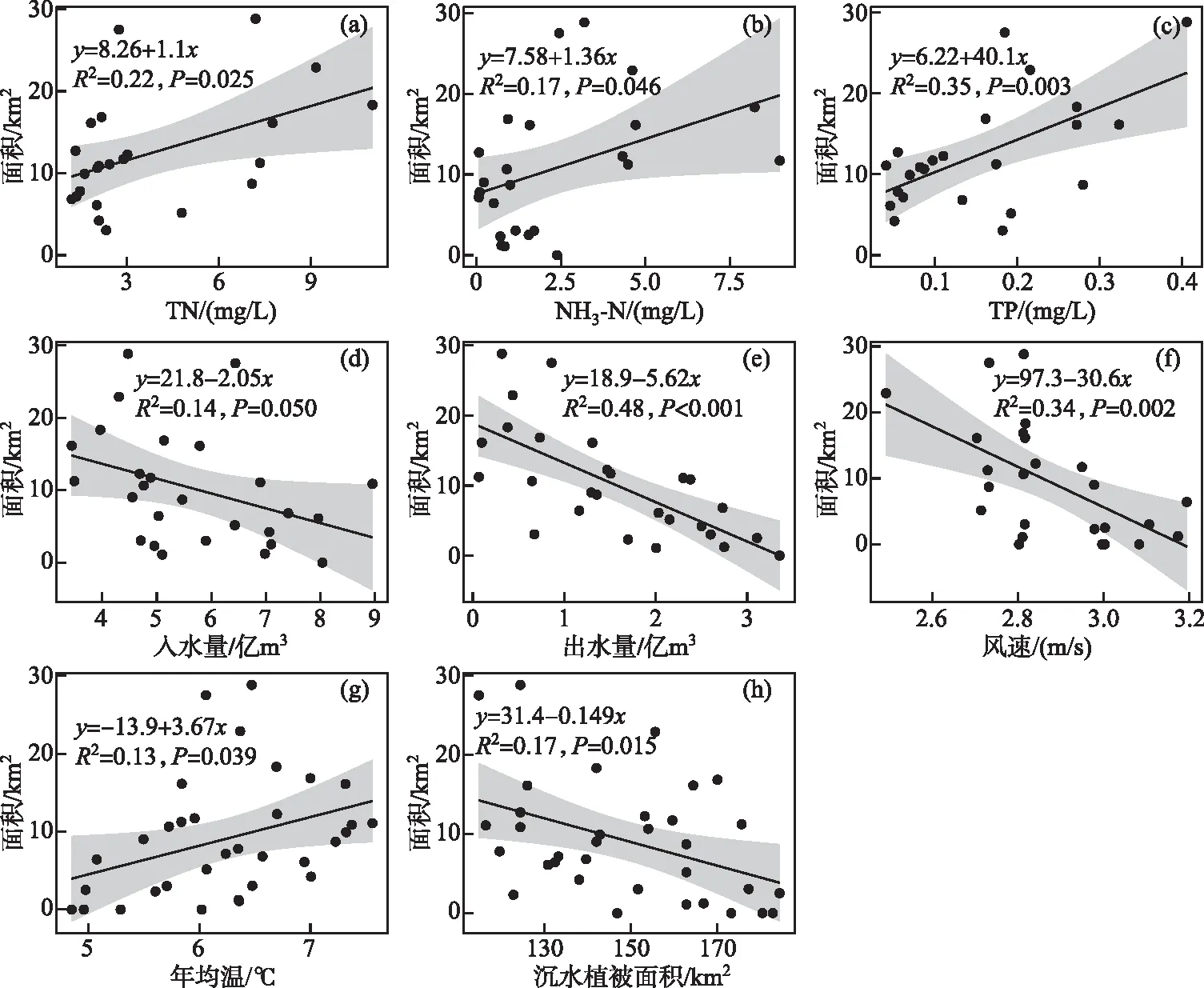
图4 乌梁素海“黄苔”暴发面积与(a)TN、(b)NH3-N、(c)TP、(d)入水量、(e)出水量、(f)风速、(g)年均温和(h)沉水植被面积的线性拟合Fig.4 The results of the linear regression between the area of metaphytic blooms and (a) TN, (b) NH3-N, (c) TP, (d) inflow volume, (e) outflow volume, (f) wind speed, (g) annual temperature, and (h) the area of submerged macrophytes in Lake Ulansuhai
2.3 乌梁素海生态补水工程实施的影响
依据“黄苔”暴发规模和生态补水工程实施情况,将乌梁素海“黄苔”暴发历程分为3个时期:无“黄苔”或小规模暴发时期(1986-1997年)、大规模暴发时期(1998-2008年)、生态补水时期(2009-2021年)。方差分析结果显示,乌梁素海不同时期“黄苔”暴发面积具有显著差异(P<0.05),表现为大规模暴发时期(1998-2008年)>生态补水时期(2009-2021年)>无“黄苔”或小规模暴发时期(1986-1997年)。乌梁素海不同时期的水环境亦发生了显著变化。其中,生态补水时期的TN、TP浓度较1998-2008年显著降低(P<0.05),出、入水量显著增加(P<0.05),且与1986-2007年无“黄苔”或小规模暴发时期的出、入水量无显著差异。而生态补水时期的沉水植被面积较1986-2007年已显著降低(P<0.05)。结果还表明,自乌梁素海“黄苔”大规模暴发开始(1998-2021年),其区域气候环境发生了显著变化,与1986-1997年(无“黄苔”或小规模暴发时期)相比,气温显著升高(P<0.05),风速显著降低(P<0.05)(图5)。
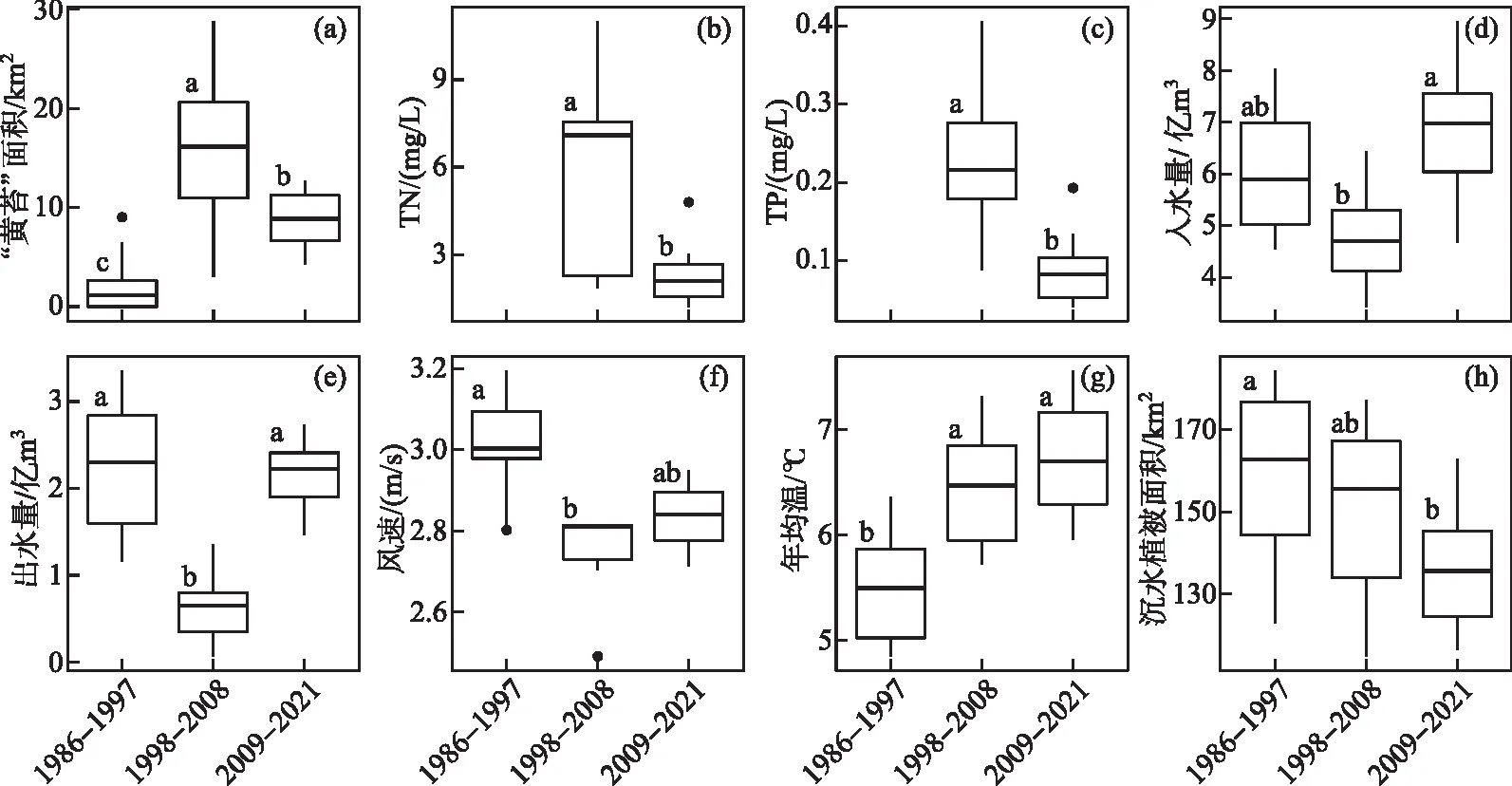
图5 乌梁素海不同时期(a)黄苔“暴发”面积、(b)TN、(c)TP、(d)入水量、(e)出水量、(f)风速、(g)年均温和(h)沉水植被面积的比较(箱体从上至下依次是75%、50%、25%分位数,箱体两端衍生线分别延伸至上极限(Q3+1.5×IQR)和下极限(Q1-1.5×IQR),黑点为潜在离群值)Fig.5 The comparison of (a) the area of metaphytic blooms, (b) TN, (c) TP, (d) inflow volume, (e) outflow volume, (f) wind speed, (g) annual temperature and (h) the area of submerged macrophytes at different periods in Lake Ulansuhai
3 讨论
3.1 营养盐和沉水植被的影响
1980s以来,受人类活动影响,尤其是乌梁素海周边工农业生产加强,大量含有TN、TP的工业废水、城镇生活污水和河套地区农业生产大量使用化肥、农药所产生的富含氮磷的农田退水排入湖体[24-25],造成乌梁素海水环境急剧恶化,氮磷含量超标,富营养化严重,可能是历史上乌梁素海暴发“黄苔”的直接诱因。本研究中,乌梁素海“黄苔”暴发面积与TN、NH3-N、TP均呈显著线性正相关(图4a~c),而这在多元线性回归模型中并未体现(表2),主要是由于在多元逐步回归算法中兼顾了模型的稳定与简便,而因子间存在显著相关关系(如TP、年均温与沉水植被面积,TN、TP、入水量与出水量等,P<0.05),因此多元回归模型最终保留了沉水植被面积和出水量2个因子。这也表明,“黄苔”、沉水植物、营养盐的内在关系较为复杂。在较低营养盐环境下,沉水植物可利用茎叶和根系分别吸收水体和沉积物营养盐生长[26],藻类因受到营养限制和沉水植物释放的化感物质抑制作用[27],通常不会异常增殖形成“黄苔”(附着藻)或浮游藻类水华;然而当水体富营养化程度增加,附着藻营养限制作用降低,逐渐形成生长优势,从而挤占沉水植被的生态位[5,7]。研究发现,沉水植物和附着藻更易利用NH3-N,但附着藻每mg氮生物量每天可吸收0.06~0.60 mg NH3-N,吸收效率是沉水植物的2~10倍[28];地中海地区Xeresa湖的研究结果表明,附着藻类生物量与TP(0~0.35 mg/L)呈显著正相关关系(对数模型,P<0.05),并在TP为0.10~0.35 mg/L时保持较高水平,沉水植被与TP呈负相关(对数模型,P<0.05)[29],因此相较于沉水植被,营养盐水平升高更有利于附着藻类的生长。
沉水植被对乌梁素海“黄苔”暴发同样具有重要影响。不同类型的沉水植被可能导致了乌梁素海和Xeresa湖的附着藻生物量对TP的响应模型的差异。一方面,沉水植物所提供的基质是附着藻生长的必要条件,否则即便水体处于富营养状态,附着藻也无法形成生长优势。加拿大11个湖泊的调查结果表明,附着藻类生物量与水体TP浓度并无显著关系,因为不同湖泊中的沉水植被类型并不相同。以两个中营养湖泊(TP约为23~24 μg/L)为例,以多分枝且叶片细碎的狐尾藻Myriophyllumspicatum为主的Massawippi湖的附着藻叶绿素a浓度显著高于以叶片宽长的美洲苦草Vallisneriaamericana为主的Champlain湖[30]。实地调研和历史资料[20]显示,乌梁素海典型的沉水植被主要有篦齿眼子菜Potamogetonpectinatus、马来眼子菜Potamogetonmalainus、狐尾藻等,这些植物叶片细碎或细长、分支密、比表面积大,是优质的附着基质,支持了乌梁素海的“黄苔”暴发。另一方面,Xeresa湖中的主要沉水植被为喜贫-中营养的轮藻属Charaspp.,由于湖体在研究时正处于富营养化程度上升期,轮藻优势衰退,附着基质的减少使得附着藻生物量对高TP(0.1~0.35 mg/L)的响应减弱;而乌梁素海的主要沉水植被为眼子菜属Potamogetonspp.,生态位较宽、耐污和适阴性强[31-33],适应较高的TP环境。本研究中,乌梁素海“黄苔”与沉水植被呈显著负相关(图4i),且沉水植被面积正显著降低(图5h),说明乌梁素海沉水植被的减少是“黄苔”暴发挤占生态位的结果,而沉水植被的持续减少可能是遏制“黄苔”暴发规模的持续扩张的原因(图3),两者互为因果。乌梁素海“黄苔”暴发是水体富营养化和湖内能够提供优质附着基质的沉水植被共同作用的结果。
虽然乌梁素海水质不断改善,氮磷浓度显著降低,但水体仍处于富营养状态(近3年TN 1.33~1.48 mg/L,TP 0.055~0.062 mg/L,TLI(Σ) 53.24~54.33), 营养盐水平仍高于TN 0.5 mg/L、TP 0.02 mg/L的藻类水华暴发边界条件[1],加之乌梁素海沉水植被覆盖度和类型仍适合附着藻定植和生长,“黄苔”规模暴发的风险依旧较大,因此需要针对乌梁素海水质和沉水植被实行更加精细化的管控。此前研究认为,乌梁素海富营养化的主要限制因子为磷[34];基于VAR模型的评价结果也表明,TP、TN为乌梁素海富营养化的主要影响因素,贡献度分别为28.78%和23.13%[35]。本研究中,乌梁素海“黄苔”对TP、TN、NH3-N均呈显著的正响应,其中TP对黄苔暴发“面积”的变差解释度达35%,高于后者的21%和17%。基于AQUATOX模型的模拟结果表明,同时削减20%~50%的入湖氮磷负荷能有效降低乌梁素海7.77%~19.54%的绿藻生物量,而单独削减20%~50%的磷负荷可降低7.96%~18.86%的绿藻生物量,与前者无明显差异[36]。上述研究支持了乌梁素海优先控磷的观点,但对于附着藻生物量是否具有同样的削减效果仍需进一步的研究验证。鉴于乌梁素海当前主要污染源为灌区的农业面源污染[37],建议加强生态沟渠[38]和人工湿地建设,截留、降低外源污染。同时,建议针对乌梁素海沉水植被类型进行适当调控,选取叶片宽大、分支较少的植被进行生态修复;还应对浅水区或静水区的生长过剩沉水植被适时收割,以减少附着藻的生长基质,同时能有效减轻植物死亡后残体腐烂造成的营养加剧等问题。
3.2 生态补水工程的影响
出入水量影响着水体更新迭代的速率和总体水位高低。较低的流速有利于附着藻的定植和生长[9],在富营养化湖泊中附着藻的生物量还随着水深的增加而降低[33]。乌梁素海生态补水工程使湖区入水量显著增加(图5d~e)。研究表明,乌梁素海生态补水明显改善了进水区的水动力,加快了水流向东、向南流动,并缩短了“黄苔”暴发严重的东大滩等湖区水龄[39]。此外,生态补水工程调引的黄河水质主要为Ⅰ、Ⅱ类(多年占比67.6%,TP<0.1 mg/L),总体上优于乌梁素海湖区水质(主要为Ⅴ类或劣Ⅴ类,多年占比92.6%,TP>0.1 mg/L)[40],因此可能得益于冲刷自净作用以及污染管控等措施,乌梁素海TN、TP浓度显著降低(图5b~c),水质发生好转。这些变化对抑制乌梁素海“黄苔”大规模暴发具有积极意义。相比之下,1998-2008年期间,乌梁素海出、入水量显著低于其他时期,水体交换速率和总体水位较低,加之营养盐浓度高、沉水植被覆盖面积大,可能进一步加剧了“黄苔”的暴发(图5)。
然而,生态补水水量的确定仍是急需回答的科学问题之一。《乌梁素海流域山水林田湖草生态保护修复试点工程实施方案》提到,基于乌梁素海水质改善目标,需对湖水每两年置换一次,即年均补水达到1.5亿m3[41];基于乌梁素海水面维持和水盐平衡目标,测算结果为每年需生态补水3亿m3以上;而基于水体交换、水力改善目标的测算研究尚鲜有报道。2018年以来,乌梁素海生态补水在3.5亿m3及以上(高于往年的0.357亿~3.2亿m3),但“黄苔”仍能在局部规模暴发,总面积无显著变化(P=0.909)。因此,建议基于“黄苔”防控目标综合考虑营养盐削减、水位维持、强化水体交换等多种需求,科学测算合理的生态补水水量,以预防“黄苔”大规模暴发。
在全球气候变化背景下,生态补水工程对乌梁素海“黄苔”暴发的抑制效果可能减弱。泛欧地区5个不同温度梯度的湖泊调查结果表明,附着藻生物量在20~28℃与水温呈显著正相关关系,意味着温度升高可能导致湖泊夏季附着藻生物量也随之增加[13]。而温度对位于寒旱区的乌梁素海影响更大,有研究报道,乌梁素海“黄苔”年内面积变化与两个月前的日均温度具有极显著的正相关性[12]。本研究亦表明“黄苔”暴发面积在年尺度上亦与气温呈显著正相关(图4g),实质上年均最低气温对其影响更大(Pearson相关系数R=0.538,P=0.006),这对未来“黄苔”防控带来了更大的挑战。风速减弱也可能会抵消生态补水工程活水的效果。研究表明,乌梁素海在无风条件下会出现大面积的滞水区,东大滩等水域水流较缓、水深较浅、沉水植被茂盛,而风场是影响这些区域湖流的主要驱动力[42],乌梁素海地区年均风速呈显著下降趋势(图5f),将会进一步加剧局部滞水或死水,从而抵消生态补水工程的活水效果。因此,建议对乌梁素海浅水、死水和滞水湖区进行有针对性地增强导流、引水活化,增强水动力、缩短水龄,以抵消风力扰动减少对“黄苔”的促进。
4 结论与展望
本文追溯了近35年乌梁素海“黄苔”自1990年出现-1998年起大面积暴发-2009年以来暴发规模得到抑制的变化历程,得到如下结论:
1)乌梁素海水体化学需氧量CODCr(P=0.090)、TN(P=0.011)、TP(P=0.001)浓度在年尺度上呈下降趋势,但仍处于富营养状态(TLI(Σ)>50)。
2)相关性分析结果表明,乌梁素海“黄苔”暴发面积与TP(P=0.003)、TN(P=0.025)、NH3-N(P=0.046)、气温(P=0.039)呈显著正相关,与出水量(P<0.001)、风速(P=0.002)、沉水植被面积(P=0.015)、入水量(P=0.050)呈显著负相关(P<0.05);多元线性回归结果表明,沉水植被面积和出水量是影响乌梁素海“黄苔”暴发面积的主要因子(R2=0.724,P=0.002)。
3)生态补水工程的实施(2009-2021年),增加了乌梁素海出入水量(P<0.05),水体中TP、TN浓度亦降低(P<0.05),同时沉水植被退化(P<0.05)减少了“黄苔”生长基质,使其暴发的扩张趋势得以遏制。
研究认为,乌梁素海当前的营养条件、基质条件仍适宜附着藻生长和“黄苔”暴发;全球气候变化背景下,乌梁素海地区气温升高和风力减弱可能会抵消生态补水工程对“黄苔”暴发的抑制效果,为未来科学有效防控带来更大挑战。研究建议,基于营养盐削减、水位保持、水力改善等目标,科学估算生态补水所需水量;实行更加严格的外源营养盐管控措施;调控沉水植被类型,适时收割过剩的沉水植被减少附着基质;通过局部导流、引水活化等措施增强滞水区、死水区水动力,以防控“黄苔”暴发。
猜你喜欢
小猕猴学习画刊(2021年10期)2021-04-18
小雪花·成长指南(2019年12期)2019-02-10
时尚北京(2018年1期)2018-01-22
水利技术监督(2017年6期)2017-12-19
都市丽人(2017年6期)2017-07-25
中国三峡(2017年4期)2017-06-06
水利科技与经济(2016年8期)2016-04-22
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
中国水利(2015年16期)2015-02-28
