
2022年极端干旱下洞庭湖区水体营养状态变化及改善对策*
2023-11-06 08:01王丹阳韩锦诚汤显强
湖泊科学 2023年6期
王丹阳,韩锦诚,黎 睿,汤显强**
(1:长江科学院流域水环境研究所,武汉 430010)(2:流域水资源与生态环境科学湖北省重点实验室,武汉 430010)(3:江西省水利科学院江西省鄱阳湖水资源与环境重点实验室,南昌 330029)
在河-湖系统中,水、沙是氮、磷、碳等关键生源要素输送的载体和动力,流速、流量、含沙量等水沙状况决定生源要素的赋存形态、迁移转化过程和输送通量,从而影响水生动植物生长所需的营养条件,一定程度上控制水生态系统[1]。尤其对于水沙交换年内变化显著的河-湖来说,由河流向湖泊的水沙输送是维持湖泊流速及自净能力、影响湖泊外源生源要素输入比重、控制湖泊在激流-静水生境间切换的关键[2]。
水文连通是水沙输移的条件,纵、横、垂向水文连通分别控制上下游、水体-岸坡、地表-地下之间的物质能量迁移,河湖连通则是河道与湖泊交换水沙及生源要素的必经之路[3-4]。极端气象事件作用于河湖连通条件,改变江湖关系,影响湖泊水环境和水生态。如在长江中游,19世纪中叶前荆江向洞庭湖分流仅通过虎渡、华容两河,1860和1870年两次强降雨引发的特大洪水后,上荆江藕池、松滋两口先后决堤,形成松滋、虎渡、藕池、华容四口分流入湖局面,极大增加了洞庭湖分水分沙能力[5]。以往学者多将极端气象事件放在更大的年内乃至多年尺度上,作为长时间序列的一部分开展研究,而较少关注气象条件直接胁迫下的湖泊水生态环境[6],尤其是对于其受水文连通条件变化的影响缺乏研究。
洞庭湖拥有长江流域最为复杂的江湖关系,其北由松滋、虎渡、太平三口接纳荆江分流,南面汇集湘、资、沅、澧四水,经东、西、南洞庭调节后由城陵矶注入长江。此外,建国以来大规模围湖造田、兴修堤垸也切断了原有的自然水文连通,许多河道流量、流速乃至流向都受人工涵闸控制[7]。在此背景下,2022年汛期洞庭湖区遭遇了极端旱情,7-9月长江流域累积降雨量不足200 mm,为1961年有完整资料以来历史同期最低。受其影响,洞庭湖水系来水量偏少近七成,水位持续消退,汛期反枯,8-9月水位均为历史同期最低,其中城陵矶站于9月15日跌至19.98 m,较历史同期偏低7.41 m。
鉴于此,本研究于2022年极端干旱事件中,在荆江-洞庭湖区河道、湖泊和堤垸开展现场勘察和水样采集工作,并结合历史数据收集,旨在(1)阐明河-湖系统中主要生源要素的赋存形态和分布特征,以及浮游植物的组成和丰度,揭示极端干旱条件下洞庭湖水生态环境状况;(2)对比极端干旱条件与历史湖泊水生态环境异同。
1 材料与方法
1.1 研究区域
洞庭湖区指荆江河段以南,四水尾闾以下,高程<50 m的湖泊水网,总面积19195 km2,其中天然湖泊约2625 km2,洪道1418 km2,其余为受堤防保护的垸区。湖区人口密集,稻田、鱼塘众多,水生动植物和越冬候鸟种类丰富。湖区水生态环境不仅关系区域生产生活可利用水资源的质量,也影响其维持生物多样性等湿地功能的发挥。
近代以来,洞庭湖区水文连通的变化主要受两方面人类活动影响。一是长江干流河段调整及水利工程建设削减三口分流分沙[5,8]。1960s下荆江系统裁弯后,三口分流、分沙比分别由裁弯前的29.4%和35.4%下降至18.8%和21.7%;2003年三峡工程运用以来,清水下泄冲刷下游河床,三口分流、分沙比进一步降至11.6% 和20.6%,且年均断流天数显著增加。二是湖区围湖造田和堤垸建设缩减湖泊面积、切断自然水系[7]。为发展农业和减轻防洪压力,建国后湖区大规模蓄洪垦殖、堵支并流合垸,湖泊容积由1949年的293×108m3减少至1995年的167×108m3,圩垸内外水体被堤防隔开,连通完全受涵闸启闭控制。
1.2 采样测试
于2022年7月22-24日、9月25-30日、10月5-7日,在53个点位采集水样:三口分流前和洞庭湖汇入后的上、下荆江干流河道;松滋、虎渡、藕池、华容河的口门、中段和尾闾;湘、资、沅、澧四水尾闾;东、西、南洞庭湖;钱粮湖垸、大通湖垸、南汉垸等堤垸内的河道、沟渠、坑塘(图1)。其中k/s-11和12为松滋、虎渡两河与澧水合流后由草尾河汇入洞庭湖的断面。7月份采样集中在垸区,9-10月份集中在除垸区外水体。
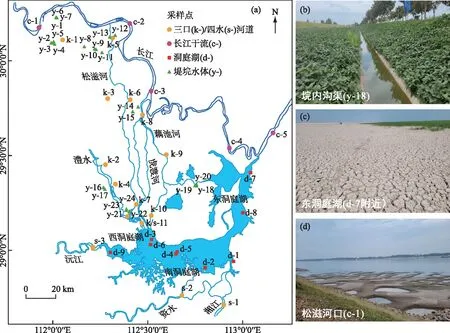
图1 采样点分布及部分样点实景Fig.1 Sampling sites and photos of some sites
1.3 数据收集与统计分析
历史时期洞庭湖区水生态环境数据部分来自2020年9月、2021年4月团队在荆江、洞庭湖的采样(图1);部分来自已发表文献和年鉴、报告等:以“洞庭湖”、“环境”为关键词检索知网,以“dongting”、“environment”为keyword检索Web of Science。TN、TP、Chl.a、综合营养状态指数(TLI)、浮游植物历史数据时段分别为1999-2017、2008-2017、2013-2017、1991-2017、1989-2017年;水情数据来自湖北省水文水资源中心(http://113.57.190.228:8001/web/Report/RiverReport)和湖南水文公众服务一张图(http://yzt.hnswkcj.com:9090/#/)。
根据序列数据是否服从正态分布,组间数据相关系使用Pearson或Spearman相关系检验判断;差异性使用t-检验或Wilcoxon秩检验判断。在α=0.05水平解释统计显著性。生源要素通量为浓度与流量之积,由于极端干旱影响下,采样期间断面流量变化较小,流量数据采用采样日平均流量。以Chl.a、TN、TP为评价指标,使用TLI判断各类水体营养状态,方法参照杨梅玲等[9]。数据处理使用SPSS软件。
2 结果
2.1 水体主要生源要素及叶绿素a浓度
三口河系中,总氮浓度在0.67~1.54 mg/L之间,以溶解态为主,占比一般超过为70%,颗粒态较少(附表Ⅰ)。硝酸盐氮是溶解态氮的主要组分,氨氮次之,二者比例约为10∶1;总磷浓度在0.15~0.26 mg/L之间,以颗粒态为主,占比约90%,溶解态较少,松滋河浓度明显高于虎渡、藕池河。氮、磷浓度在三口河系呈尾闾>中段>口门的梯度分布,尤其在松滋河最为显著。溶解态有机碳和叶绿素a浓度分别在31~44 mg/L和2.8~5.7 μg/L之间,不同河流、不同河段之间无显著差异。四水尾闾中,总氮浓度在0.44~0.67 mg/L之间,平均0.55 mg/L;总磷浓度在0.18~0.41 mg/L之间,平均为0.32 mg/L。氮以溶解态为主,硝态氮、氨氮比重约8∶1,磷以颗粒态为主。溶解态有机碳和叶绿素a浓度分别在28~36 mg/L和2.8~4.1 μg/L之间。相比三口河系,四水氮、磷浓度显著较高,但碳、叶绿素a浓度差异不大。
长江干流总氮、总磷平均浓度分别为0.90和0.23 mg/L,在5个采样断面间没有显著差异。洞庭湖总氮浓度在0.52~0.63 mg/L之间,平均为0.57 mg/L,东、西、南洞庭湖间无显著差异,但明显低于长江干流;总磷浓度在0.39~0.49 mg/L之间,平均为0.45 mg/L,明显高于长江干流。溶解态有机碳浓度长江和洞庭湖均在38 mg/L左右,叶绿素a浓度长江和东洞庭湖较高,西、南洞庭湖较低。在洞庭湖垸内,溶解态有机碳浓度在沟渠、鱼塘、荷塘间差异较小,均在30 mg/L左右波动,氮、磷和叶绿素a浓度则呈现荷塘>鱼塘>沟渠的梯度分布,尤其池塘中溶解磷浓度可达沟渠中的4~6倍。相比垸外湖泊和河道,垸内碳浓度略低,氮、磷浓度显著偏高,叶绿素a浓度大致处于同一水平。
2.2 水体浮游植物种类及丰度
洞庭湖区水体浮游植物共检出蓝藻、绿藻、硅藻、隐藻、甲藻、裸藻6个门类下49个藻种,其中东、西、南洞庭湖检出6门37种,长江及三口四水河道中藻类多样性略低,为6门30种(附表Ⅱ)。河、湖水体中均以硅藻门和绿藻门为主,但藻种存在一定互斥性,如细鞘丝藻、裸藻在河道水体中均有检出,但未在湖泊中发现,粗刺藻、囊裸藻等则只存在于湖泊中。藻类生物量以长江最高为1.551 mg/L,洞庭湖次之,平均1.337 mg/L,三口四水较低(图2)。除南洞庭湖和四水中浮游植物生物量以甲藻为主外,其余水体中绿藻和硅藻占总生物量的大多数。

图2 洞庭湖、三口四水、长江中浮游植物生物量Fig.2 Phytoplankton biomass in Lake Dongting, three outlets and four rivers, and Yangtze River
3 讨论
3.1 湖区生源要素通量
采样期间,枝城站平均水位38.05 m,松滋、虎渡、太平口断流,荆江与三口河系的纵向水文连通切断,三口分流量为零,河道口门处有积水但无流速,因而荆江向三口河系输送的氮、磷、碳通量均为0(图3)。洞庭湖向荆江输送的氮、磷、碳通量分别为2.3、0.83和67 kg/s,其中来自三口口门至尾闾区间(含澧水)的占比分别为10.4%、18.1%和21.6%,来自四水的占比分别为22.6%、33.7%和41.6%,来自湖区自源生产的占比分别为67%、48.2%和36.8%。
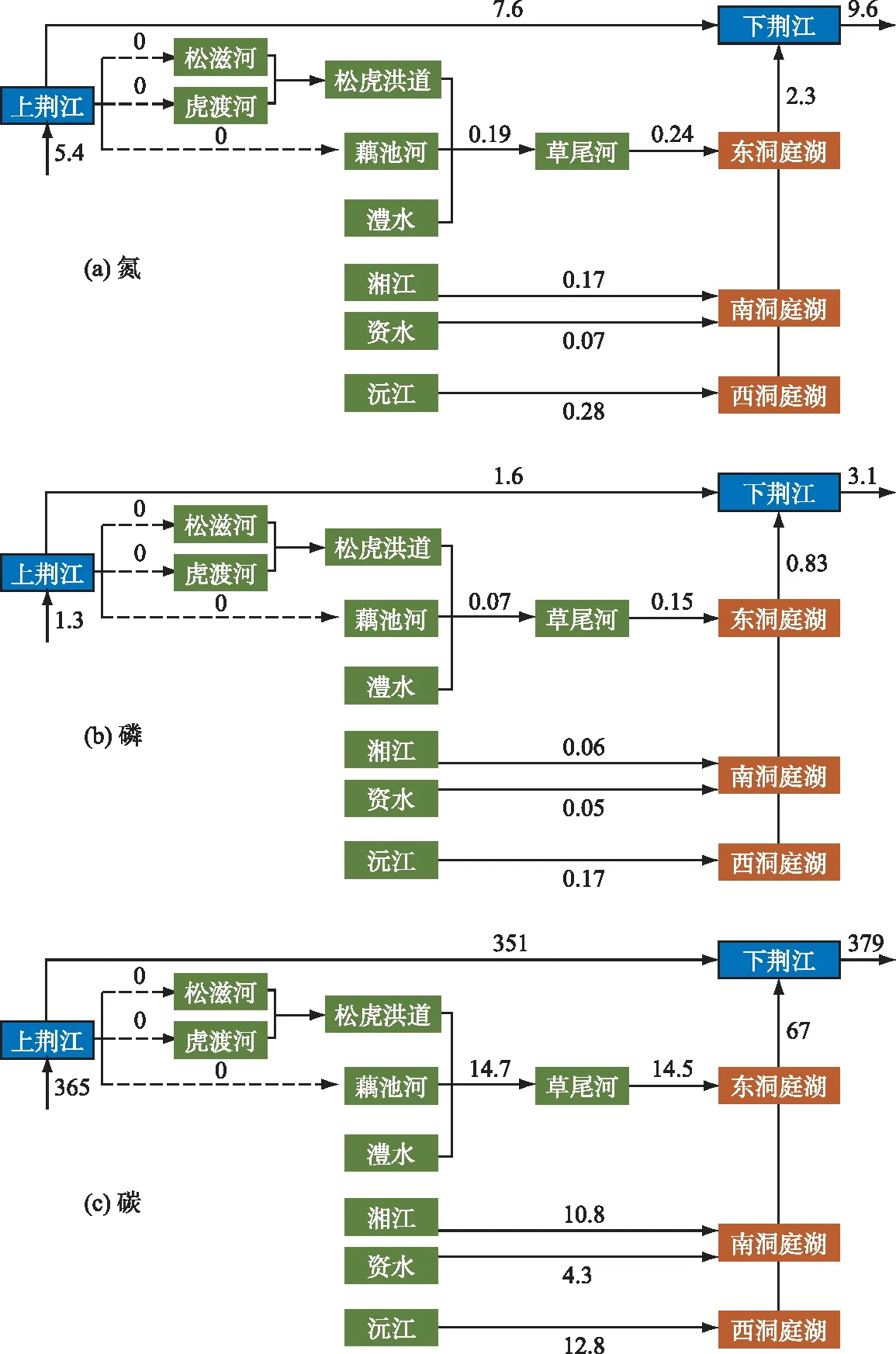
图3 洞庭湖区氮、磷、碳输送通量(单位:kg/s)Fig.3 Fluxes of nitrogen, phosphorus, and carbon in Lake Dongting area
生源要素通量直接由浓度与流量决定,因而河湖之间纵向水文连通越强,流量越高,通量就越大[10]。在极端干旱状态下的洞庭湖区,流量较大的湘、资、沅、澧四水(平均约249 m3/s)向湖泊输送的生源要素远超过流量较小的松滋、虎渡、藕池河(平均不足20 m3/s),且三口河道中生源要素只来自本地积累,无长江补给。这使得洞庭湖区完全成为长江的生源要素源,无法通过三口分流发挥汇的功能,可能加重城陵矶以下江段氮、磷、碳等的负荷。
外源输入量的调整影响内、外源生源要素的相对比重。在极端干旱下的洞庭湖,由于三口四水补给减少,更多的生源要素来自湖区生物地球化学循环下的自源生产,因而由东洞庭湖汇入长江的氮、磷中,自源比重均超过或接近50%。内、外源相对贡献的变化可能改变生源要素组分。以碳为例,在外源输入占比较高的丰水期,河岸带地表和地下径流在汇入河道的过程中,沿程冲刷、交换、携带土壤颗粒中沉积的老碳(微生物量碳含量较高),导致湖泊内微生物碳占比较大[11],而在以自源生产为主的枯水期,湖泊碳则可能主要来自湖底凋落物及表层浮游动植物死亡后的分解[12]。
3.2 垸内生源要素富集
洞庭湖区堤垸内的沟渠、水塘等各类水体中,溶解态氮、磷、碳浓度都显著高于外部河湖,综合营养状态指数也处于较高水平,尤其荷塘TLI超过了50,是所有水体中唯一达到富营养状态的区域(图4)。已有研究中也有报道类似现象,长江中游洞庭、鄱阳湖区,氮、磷浓度在外河一般可以达到III类水标准,而圩垸内则多为劣V类,如在鄱阳湖饶河联圩,圩内车马湖、农沟和圩外饶河的TP浓度依次为0.17、0.15和0.07 mg/L[13]。
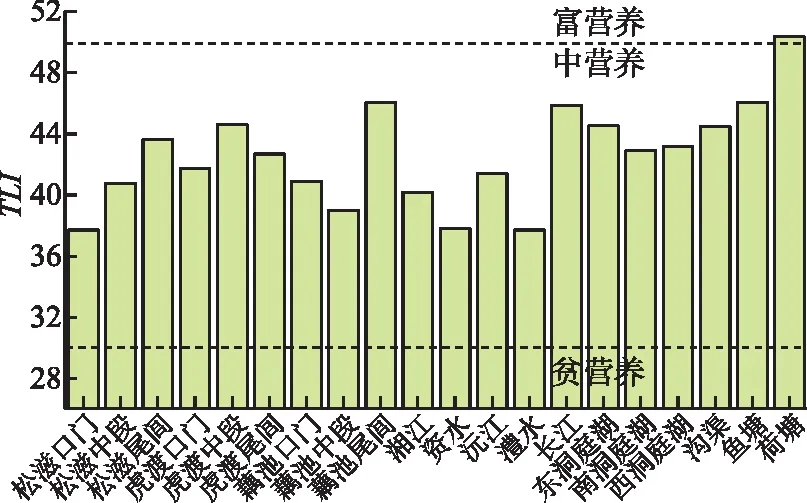
图4 洞庭湖区综合营养状态指数Fig.4 Comprehensive trophic level index in Lake Dongting area
垸内生源要素循环受垸区防洪和用水策略影响。一是“内外河分开”,加固已有堤防工程,内水无法参与外循环,易在闸口附近形成回水区造成更严重污染[14]。二是“排灌渠分开”,规定了灌渠由外河经涵闸引水经各级水渠进入田面和鱼塘,排水渠由田面、鱼塘、洼地经各级水渠过涵闸排向外河。由于外河水质较好,而田间和鱼塘排水含有大量来自化肥施用和鱼类排泄物的营养盐,排灌分开往往造成营养盐浓度沿灌-用-排路径依次升高,如在江陵县三湖农场垸区,TP浓度在灌溉渠、田边围沟和垸内排水干渠(清南渠)分别为0.08、0.10和0.17 mg/L[15]。
修建堤坝和涵闸,人为控制自然河湖与垸区水体的交换,是造成垸内生源要素富集的根本原因。当涵闸开启时,水体沿1-2-3路径完成内外水循环,外河补给内水,垸内周期性换水,排泄来自5、8、11的生源要素(图5)。一旦涵闸关闭,内外水循环路径被切断,无法闭环,而垸内水体仍源源不断地通过地表和地下径流汇集来自农田、居民生活等的面源污染,造成生源要素在垸内水体积累。遭遇极端干旱时,外河水位下降,涵闸提水困难,1、3、17路径被切断,失去水文连通,垸内氮、磷、碳等浓度升高。
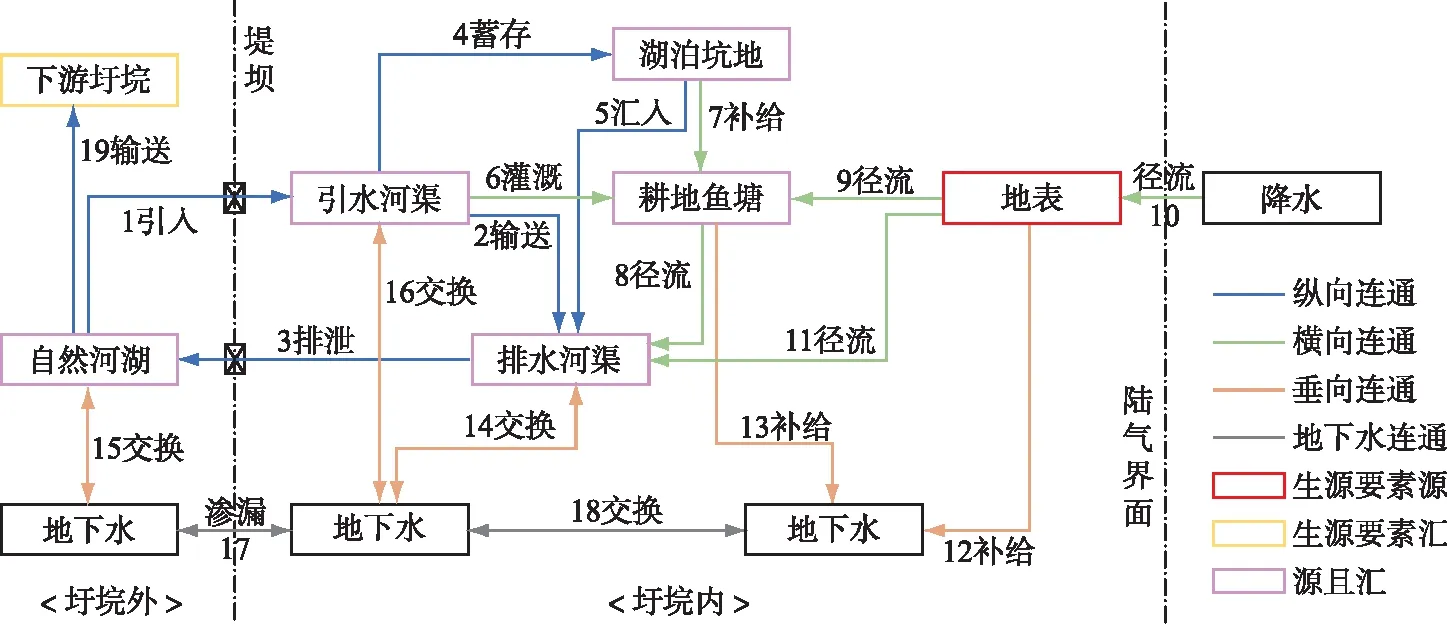
图5 堤垸内外生源要素迁移路径Fig.5 Paths of biogenic element transport between inside- and outside-polders
3.3 与历史水平对比
将本次采样数据与论文、报告等公开发表历史数据对比,2022年极端干旱时段,洞庭湖水体中总氮浓度处于历史低位。过往数据显示,总氮在洞庭湖总体呈枯水期(1.74 mg/L)>平水期(1.65 mg/L)>丰水期(1.49 mg/L)的规律,但在2022年9月,总氮浓度不仅低于往年枯水期的平均水平,甚至比绝大多数年份的丰、平水期浓度还要低(图6a)。9月份极端干旱时,三口断流,洞庭湖失去来自长江的氮补给,而城陵矶段主航道依然畅通,湖内自源生产的氮仍可以不断排出;另一方面,洞庭湖近岸多被开垦为耕地,施用较多氮肥,干旱时由于水位下降,近岸土壤出露,不再向水体释放氮。因此,水中氮收入减少而支出稳定,可能改变了湖区的氮收支,导致了较低的水体中氮浓度。
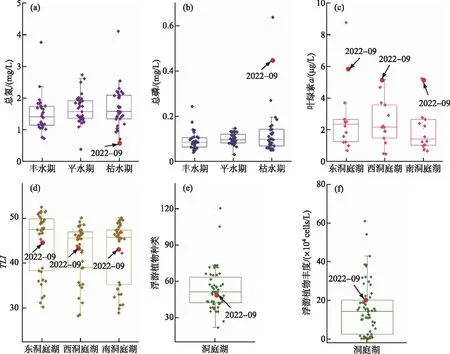
图6 洞庭湖2022年极端干旱时段与历史时期氮、磷、叶绿素a、TLI、浮游植物丰度对比Fig.6 Nitrogen, phosphorus, Chl.a, TLI, and phytoplankton abundance in Lake Dongting in the extreme drought of 2022 compared with history
2022年极端干旱时,洞庭湖总磷浓度处于历史阶段高位,与总氮相反(图6b)。三峡工程运用以来,大量泥沙被拦截在库区,下游清水下泄,由三口入湖的泥沙含量锐减,由于磷主要以颗粒态附着于泥沙之上,因而入湖磷通量也有所减少。即便如此,在外源磷输入随三口四水入湖水量而下降的极端干旱条件下,湖内磷浓度仍远超往年丰、平、枯水期平均值,说明磷可能多来自湖内自源生产。在各类水体中,藻类生物量与Chl.a浓度呈显著正相关,与各生源要素浓度无显著相关性。由于水位下降、水深变浅导致水体透光率上升,同时流速放缓,夏季温度升高,极端干旱创造了浮游植物的适宜生长环境,造成相比历史阶段具有较高的浮游植物生物量和叶绿素a浓度(图6c、f)。植物残体死亡后破碎分解,可能是导致颗粒磷以及总磷浓度较高的原因[16]。尽管极端干旱时洞庭湖失去了长江的汇入,但其浮游植物种类(图6e)处于历史平均水平,并未显著下降,说明在失去外源输入的情况下,湖区自生生源要素的浓度和组成仍能满足各类藻种生长所需。计算综合营养状态指数低于历史阶段不同水期的平均值(图6d),这可能与总氮浓度较低而权重较高有关。
3.4 干旱影响下水生态环境改善对策
在气候变化和人类活动的双重影响下,未来洞庭湖可能经历更大程度、更高频率的干旱事件[17],带来水体富营养化等潜在威胁。长久之计,一是通过河道工程建设和三峡调度运用,改善江湖关系,恢复水文连通,加快长江与洞庭湖换水速率,提高湖泊水面率,从而增强水体自净能力,降低氮、磷等污染物浓度[18-19];二是继续退耕还湖,修复沿湖经常性被淹没的高氮、磷浓度的土壤,减少内源污染释放。但是,洞庭湖区的复杂江湖关系和艰巨防洪任务决定了湖区任何江湖关系的调整首先应考虑防洪安全。考虑一些河道工程可能会给江湖关系带来难预料、难逆的变化,应将治理富营养化的水环境目标与防洪、用水目标结合综合调控[20]。
垸区以稻田种植为主,化肥施用使大量氮磷等进入水体,而堤垸的防洪功能又限制了环境基础设施的建设,受污染水体难以得到治理,可利用水资源量减少。在防洪优先的前提下,汛期主动蓄洪,建设水源库、净化水质,补给枯季用水是协调水生态环境改善与水灾害防治、水资源利用的可行措施。建议在近外河堤的低洼处、围洼筑堤、建设水库,水库背倚外河堤坝,内设两闸门,分别连通引水河渠和排水河渠。水库在汛前腾空,汛时开闸分洪,汛末蓄水。一方面恢复圩垸水面率,汛期既发挥蓄洪作用,又有序引导洪水入垸,避免淹没生产生活设施;另一方面枯水期或外河涵闸关闭时,由于水库连通引、排水渠,圩垸内仍可完成水循环闭环,保持水体自净能力,削减污染的同时为社会经济活动提供水资源。
与主动蓄洪的相近概念和实践有很多。1980s长江中下游圩垸已开展围洼滞蓄,将圩垸内高地来水导入低地筑矮堤圈成的滞蓄区临时储存,枯水期抗旱并发展多种经营。2010年,湖南省水利厅提出除加强堤防和蓄洪建设外,更应考虑蓄水保水和生态保护等新机制,通过扩内湖、建低坝和综合枢纽工程等方式,利用蓄洪区低洼地,根据垸内生产和环境用水需求有计划地进水和吐水,蓄水深1~1.5 m,每年蓄洪10×108~15×108m3[21]。可见,将水库作为圩垸内长期水利设施,不仅使之发挥蓄洪和供水功能,还利用水库连通圩垸内各水体,使外河涵闸关闭时内水仍保持一定流速和净化能力。如果能将环境水库建设与垸内各级河道、沟渠的护岸水土保持措施和水体生物净化措施结合,则有望实现水环境保护与水灾害防治、水资源利用的目标统一与协调。
4 结论
2022年夏季,洞庭湖区遭受极端干旱,松滋、虎渡、藕池三口水系断流,湘、资、沅、澧四水来水减少,湖泊水位下降,面积萎缩。在此影响下,洞庭湖总磷和叶绿素a浓度达到历史高值,浮游植物生物量也超过历史平均水平,但总氮浓度却处于历史低位。受洞庭湖-长江水文连通削弱影响,洞庭湖来自三口的氮、磷、碳等生源要素通量减少,来自四水和湖内自源生产的比重增加;受人为控制自然河湖与垸区水体交换的影响,垸内水体换水不畅,富营养化程度高于外部自然河湖。改善湖区水文连通,削减内源污染释放,协调水环境与防洪、用水的关系,在垸内通过主动蓄洪建设水源库,是应对未来干旱下湖泊水生态环境问题的可行对策。
5 附录
附表Ⅰ和Ⅱ见电子版(DOI: 10.18307/2023.0624)。
猜你喜欢
小学生学习指导(低年级)(2021年10期)2021-11-01
金桥(2020年8期)2020-05-22
少儿美术(快乐历史地理)(2019年4期)2019-08-27
阅读(低年级)(2018年4期)2018-05-14
小学阅读指南·低年级版(2017年2期)2017-03-23
音乐天地(音乐创作版)(2016年4期)2016-07-28
衡阳师范学院学报(2016年3期)2016-07-10
林业与生态(2016年3期)2016-02-27
作物研究(2014年6期)2014-03-01
作物研究(2014年6期)2014-03-01
