
北京两个水库的桃花水母及其发生条件初探*
2023-11-06 08:01王东霞熊志明侯新觉黄振芳王忠锁
湖泊科学 2023年6期
王东霞,熊志明,侯新觉,郭 伟,黄振芳**,王忠锁**
(1:北京市水文总站,北京 100048)(2:首都师范大学生命科学学院,北京 100048)
桃花水母是温带广布的濒危淡水腔肠动物(Coelenterata),属笠水母科(Olindiidae)桃花水母属(Craspedacusta),为珍稀的古老水生物[1]。其生活史包括漫长的无性世代(螅状体,polyp)和短暂的有性世代(水母体,medusa)。螅状体体长不长于3.0 mm,着生于底质固形物或水生生物体表,难以观察,故了解较少[2]。水母体发育期约30天,性成熟阶段伞径可达20 mm左右,易于观察研究[3]。通常报道的桃花水母指的是其水母体[1-3]。
目前公开发表的桃花水母共12种,其中学界公认的仅3种,即索氏桃花水母(C.sowerbyi)、中华桃花水母(C.sinensis)和伊势桃花水母(C.iseanum)[1,4-5]。其余9种为中国学者先后发表,但近年遗传学证据提示应存在一定的同物异名[1,6]。分类体系不完善影响了相关物种的分布格局及环境相关性等进化适应机理的认知[5]。2005年后,随着DNA barcode技术的应用,18S rRNA、16S rRNA、ITS和COI等标记基因相继用于桃花水母的物种鉴定和系统学研究,对桃花水母系统学及生态学研究起到积极的促进作用,其中ITS对桃花水母物种区分度较好[6-7]。
桃花水母的传统研究多关注形态学或遗传学特征的描述和分类鉴定,注重生物学特征描述而忽视环境条件调查,注重水母体静态观察和实验而忽视其发育过程和形态变化,注重水母体发生事件的报道而忽视其发生机理的探究[5,7-8]。就水母发生发展的条件和驱动因子而言,国外学者多认可桃花水母(实为索氏桃花水母)是广域性物种,对水环境质量和生境类型敏感性低、入侵力强,高度关注其种群动态、扩张趋势及入侵机制,致力于桃花水母防范和管控技术研究[9-12]。近年来,桃花水母全球扩张呈加速趋势,索氏桃花水母已扩布到英国、德国、法国、意大利、罗马尼亚、波兰、捷克、俄罗斯、丹麦、荷兰、比利时、瑞士、日本、菲律宾、加拿大、美国、巴拿马、智利、澳大利亚和新西兰等亚洲、欧洲、美洲和大洋洲等广大温带水域[5,8-10]。相关入侵生物学和环境适应力的研究逐渐增多。诸如桃花水母与富营养化及水色的相关关系[10-11]、对营养盐垂直迁移的影响[12]、对大型浮游动物群落结构和生态系统弹性的扰动[13],以及全球变化背景下入侵扩张的风险防范等逐渐成为主流[9]。
国内学者则从受威胁物种保护的角度探究桃花水母的发生与水环境质量优劣间的关联,认可桃花水母为濒危物种,依赖清洁水体生存并认同桃花水母为清洁水体指示物种[1,5]。多数报道称相关水体桃花水母的发生应归因于水质改善,而桃花水母发生频次的降低则是环境污染和人为扰动所致[14]。问题在于,这些报道多未涉及环境因子和生境特征的专业调查和分析,清洁水指示种的提法缺乏数据支持和必要的论证。桃花水母与水环境质量相关性研究亟待加强。
2022年9月上旬北京怀柔水库和黄松峪水库同时发现桃花水母,课题组于9月9日和10日开展了实地调查和样本采集。基于有限样本的形态学和分子系统学分析,本文首先完成2个水库样本的物种鉴定和相关物种核校,进而结合水生态常规监测中获取的理化指标和浮游动物数据,探究桃花水母发生的潜在驱动因子,并就激发桃花水母螅状体启动有性生殖的环境条件提出了温变胁迫假说(temperature stress hypothesis)和营养限制假说(nutrition limitation hypothesis),以期为系统开展桃花水母生态学研究、揭示水母体发生发展规律提供借鉴。
1 材料和方法
1.1 研究区域
黄松峪水库(40°14′58.22″~40°15′46.04″N, 117°15′4.45″~117°15′55.68″E)是北京市平谷区东北部泃河上游的一座中型水库,位于黄松峪乡境内。黄松峪水库总库容0.1×108m3,流域面积49 km2(图1)。怀柔水库(40°18′40.89″~40°20′46.03″N, 116°35′28.44″~116°37′56.94″E)为北京市怀柔区境内的大型水库,为京密引水渠重要调蓄水体,南水北调中线通水后,该水库演变成向密云水库反向调水的重要调蓄水体。怀柔水库总库容1.4×108m3,流域面积540 km2(图1)。2个水库均位于温带半湿润季风气候带,年均气温10.5℃,年降雨量640 mm。夏季温热多雨,适宜水生生物生长,冬春寒冷少雨,水面冰封,水生生物活性降低或休眠。
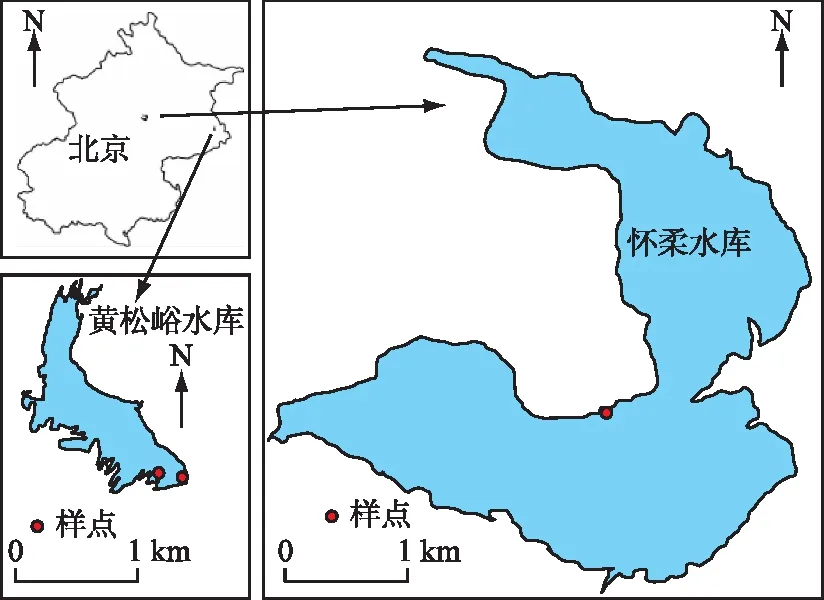
图1 北京桃花水母样本采集地Fig.1 Sampling sites of Caspedacustas spp. in Beijing
1.2 样本采集、培养实验分析
桃花水母样本采集于2022年9月9-10日。以密眼手抄网拉近目标个体后连水舀起置入带盖容器内增氧运输回实验室。室内用5000 mL玻璃烧杯,见光常温培养观察水母活动节律、伞径大小和性腺形态。通过照片和视频计数水母触手数目。桃花水母平衡囊形态观察和数目计数采用装片显微观察并计数。触手上刺丝囊疣的分布状况同样通过显微观察并拍照记录[5,8]。共观察计数桃花水母17只,其中怀柔水库8只,黄松峪水库9只。
桃花水母物种鉴定参考和振武(2005)的分类体系,主要依据刺丝囊疣形状和排列方式、缘膜内平衡囊形状及数目、生殖腺形状和触手数目及级数等特征[1]。DNA条形码辅助鉴定采用核基因组ITS(ITS1-5.8S rDNA-ITS2)碱基序列片段[6,15]。
水母DNA 提取前,以匀浆缓冲液浸泡95%乙醇保存的桃花水母过夜,重复2次,以去除水母体内外夹带的其它生物及粘附的环境DNA。组织DNA提取采用磁珠法动物组织基因组DNA提取试剂盒 (天根生化科技有限公司)。扩增ITS的引物为ITS-F1: GTCGTAACAAGGTTTCCGTAGG和ITS-R1: GGTAGTCTTGCCTGATCTGAGG,扩增片段长度约750 bp。扩增反应体系总体积50 μL,包括去离子水20 μL,2×Taq MasterMix (天根公司)25 μL,上下游引物各1 μL(10 μmol/L),模板DNA 3 μL(浓度100 μg/L)。扩增反应条件为94℃变性4 min;94℃变性50 s,55℃退火50 s,72℃延伸90 s,35个循环;72℃延伸10 min,4℃保存[15-16]。
反应产物于1.5%琼脂糖凝胶电泳检测后,送睿博兴科生物技术有限公司测序。经过双向测序获得校正拼接后的序列与 GenBank 数据库中收录的桃花水母ITS 基因进行序列相似性比对; 利用 MEGA X软件计算序列间的相对遗传距离,构建邻接系统树(Neighbor joining tree),聚类分析北京桃花水母与已发表桃花水母的遗传关系,各分支的置信度经Bootstrap 1000 次循环检验。
1.3 水环境因子采样和实验分析
浮游动物定量样品采自水面下0.5 m水层,以有机玻璃采水器采集5 L水样,置水样瓶中加75 mL鲁哥试剂固定。室内静置48 h后虹吸去除上清,分三步沉淀浓缩后得30 mL置50 mL标本瓶中加1.5 mL甲醛液定容至30 mL,4℃保存。浮游动物鉴定和计数时,先充分摇匀沉淀浓缩的样本保存液,快速吸取1 mL置1 mL浮游生物计数框中于Leica DM2500显微镜100×完成镜检计数。每样本重复两次镜检计数,取两次计数平均值计算各物种个体密度并依据个体生物量换算浮游动物总生物量密度。详细实验方法参考文献[17]。
水质理化指标同样测定水面以下0.5 m水层水样。水温(WT,℃)、溶解氧(DO,mg/L)、pH等指标采用溶氧仪(YSI Pro ODO)和pH计现场测定。总氮(TN,mg/L)和总磷(TP,mg/L)取1.5 L水样调节pH至2.0,带回实验室以多参数水质测定仪(6B-2000,盛奥华)24 h内完成测定。生化需氧量(BOD,mg/L)采用BOD自动检测仪完成培养和测定。水环境理化指标测定详细方法参考文献[18]。数据的统计分析和制图均于Microsoft Excel 2010完成。
2 结果与分析
2.1 北京桃花水母形态学鉴定
怀柔水库产桃花水母伞体为扁半球形,伞径13~21 mm之间,平均(15.8±3.1) mm。伞缘触手数目变动于266~398之间,平均(342±48)个,其中4个主辐触手明显长于其它触手(图2a)。触手上具刺丝囊疣,呈圆锥状,环状螺旋排列(图2b)。胃大,垂管长,伸达缘膜孔之外,较生殖腺略长。垂管末端为口,具4片较大的唇。生殖腺4个,着生在主辐管与垂管联结处,呈大囊状,色浅近白色(图2a、d)。缘膜内平衡囊管状,88~189个,平均(140±27)个(图2e)。综合以上重要形态特征,鉴定怀柔水库桃花水母为广布物种索氏桃花水母(Craspedacustasowerbyi)。
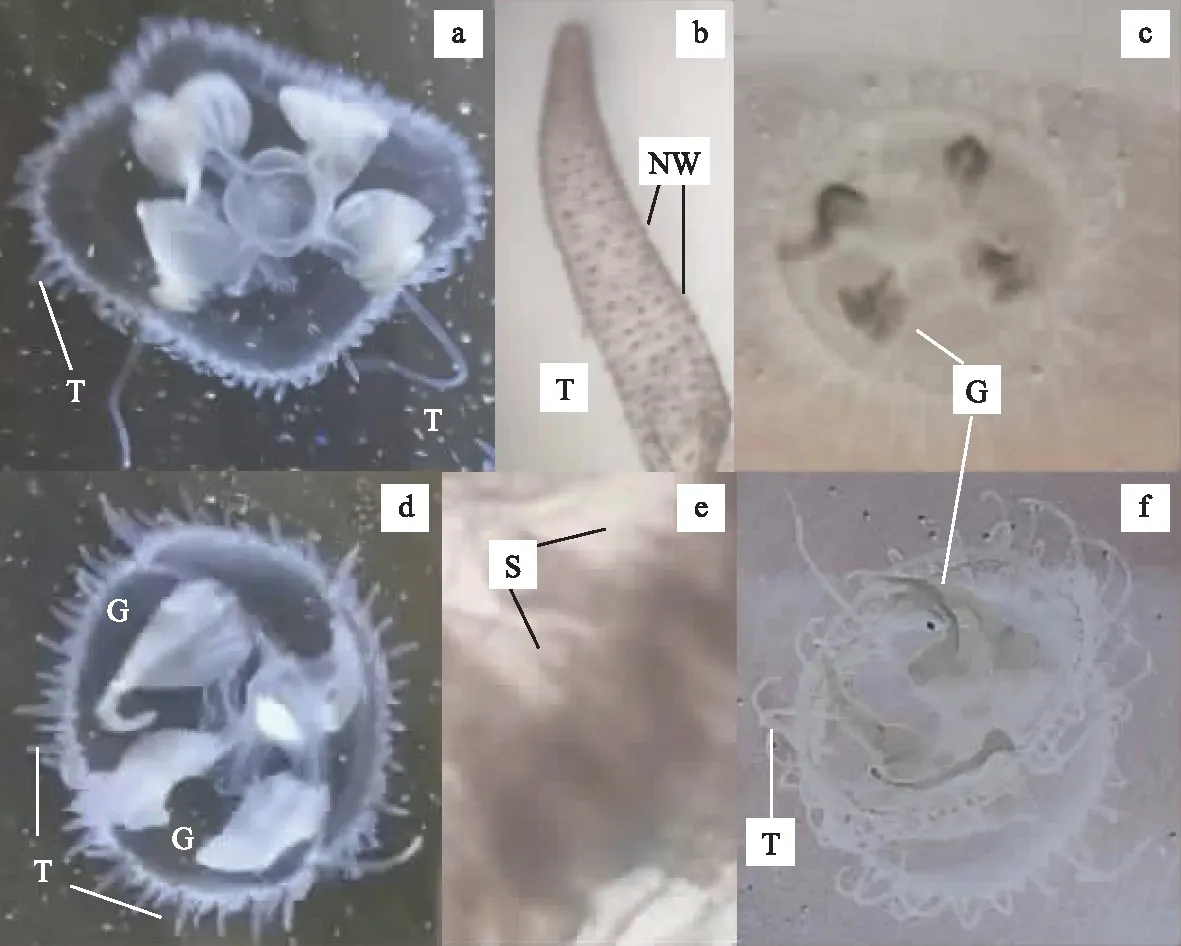
图2 北京怀柔水库和黄松峪水库桃花水母主要鉴别特征(a、d示怀柔水库索氏桃花水母,c、f示黄松峪水库嘉定桃花水母,b示触手上刺丝囊疣排列方式,e示平衡囊形态。T-触手,NW-刺丝囊疣,G-性腺,S-平衡囊)Fig.2 Taxonomic traits of the jellyfishes sampled from the Huairou and Huangsongyu Reservoirs of Beijing
黄松峪水库产桃花水母形态特征与怀柔水库非常相似,其伞径略小,变动于11~18 mm之间,平均(13.2±1.9) mm。伞缘触手数目变动于260~281之间,平均(268±5.4)个,其中4个主辐触手明显长于其它触手(图2f)。刺丝囊疣圆锥状,环状螺旋排列触手上(图2b)。缘膜内平衡囊管状,132~136个,平均(134±2)个。生殖腺小囊状,口径较小,略呈细长带形,灰色或淡绿色(图2c、f)。综合以上重要形态特征,鉴定黄松峪水库桃花水母为嘉定桃花水母(C.kiatingi)。
2.2 北京桃花水母分子系统学地位
ITS基因测序结果显示北京怀柔水库2个样本间遗传距离小于0.001(遗传相似性100%),与已报道的索氏桃花水母和信阳桃花水母(C.xinyangensis)遗传相似性分别为100%和99%(表1,图3)。黄松峪水库样本与怀柔水库样本间遗传距离为0.210(遗传相似性97.9%),与已报道的嘉定桃花水母(C.kiatingi)和秭归桃花水母(C.ziguinsis)遗传相似性分别为100%和95%。黄松峪样本与嘉定桃花水母遗传距离介于0~0.002 之间,与秭归样本距离为0.037;怀柔水库样本与大连产索氏桃花水母以及鼋头渚和安吉产信阳桃花水母的遗传距离介于0~0.008之间(表1)。

表1 北京产桃花水母与其它地区桃花水母ITS基因遗传距离*Tab.1 Genetic distances between Craspedacusta spp. from Beijing and other areas based on ITS gene sequence

图3 北京与其它地区桃花水母ITS遗传关系邻接系统树(菱形标为北京样品)Fig.3 Phylogenetic tree of Craspedacusta ITS in Beijing and other areas based on neighbor-joining method(The rhombus shows the Beijing sample)
系统发育聚类分析显示秭归产短手桃花水母(C.brevinema)实为中华桃花水母同物异名,青城山产四川桃花水母为嘉定桃花水母同物异名,鼋头渚和安吉产信阳桃花水母是索氏桃花水母同物异名,而秭归产秭归桃花水母可能是不同于近缘种嘉定桃花水母的新的物种(图3)。据此确定北京怀柔水库样本(BJ2和BJ3)与鼋头渚、安吉和大连产桃花水母同为索氏桃花水母(C组),而黄松峪水库样本(BJ1)与郫县、云台山、虎峪、德阳和青城山产桃花水母同为嘉定桃花水母(B组)。B组和C组间遗传相似性高于A组中华桃花水母,表明索氏桃花水母和嘉定桃花水母亲缘关系较近,而与中华桃花水母进化分化较早(图3)。
2.3 桃花水母发生的环境条件
怀柔水库和黄松峪水库属于中小型水库,库区环境较为稳定。2022年度监测的水环境理化指标中pH、DO、BOD、TP和TN等因子年内稳定,桃花水母发生前后未显示明显的起伏变动,对桃花水母发生(启动有性生殖)的影响有限,但水温和饵料生物丰度呈快速变化,具有潜在驱动作用(图4)。
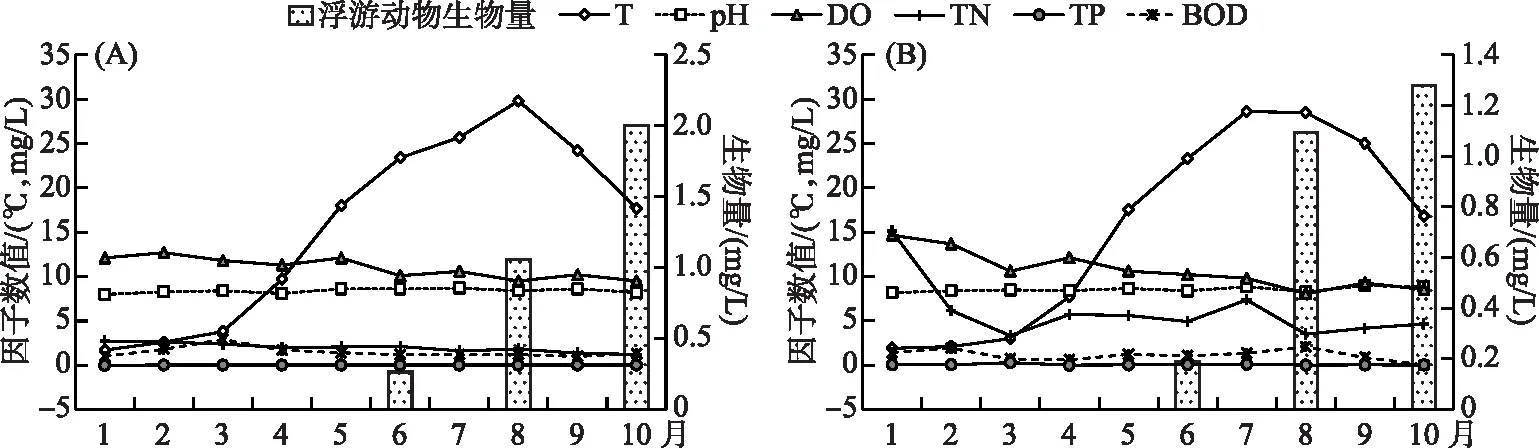
图4 怀柔水库(A)和黄松峪水库(B)主要环境因子和浮游动物生物量动态Fig.4 Seasonal dynamics of major environmental factors and zooplankton biomass in Huairou (A) and Huangsongyu (B) Reservoirs
2022年怀柔水库表层水温峰值发生在8月上旬(29.8℃),其后快速下降,至9月上旬跌至24.2℃。黄松峪水库表层水温峰值发生在7月底,8月上旬水温开始下降,至9月初降至25℃。2个水库桃花水母发生时水温均处于快速下降进程,且跌破25℃。一个月内降幅近5℃。降温或成为桃花水母有性生殖的关键环境胁迫。
桃花水母主食的轮虫、枝角类和桡足类等大型浮游动物的存量在夏秋季快速增长。怀柔水库和黄松峪水库浮游后生动物生物量在生长季之初(6月初)仅0.26和0.19 mg/L,至8月份分别增长到1.06和1.09 mg/L 并保持增长趋势,至晚秋10月初分别达2.00 和1.28 mg/L(图4)。由此推断9月上旬两库区浮游后生动物生物量分别高于1.50和1.20 mg/L。充足的食源或诱导桃花水母启动有性生殖。
3 讨论
3.1 桃花水母的分布格局
桃花水母全球分布集中在温带地区,但各物种的分布范围和环境适生性存在明显的差异。受国内物种分类不确定性影响,桃花水母物种生物地理格局尚不明确[5]。国外的桃花水母品种单一,物种分布情况比较明显。除日本有伊势桃花水母外,欧洲、美洲、大洋洲、非洲和亚洲其它国家记录的桃花水母均为单一的索氏桃花水母[1,5,9]。国内发表的11个桃花水母中,得到ITS支持的除索氏桃花水母和中华桃花水母外,还有嘉定桃花水母[15]。
本研究依据桃花水母形态学特征,辅以核基因ITS序列比对确认了怀柔水库索氏桃花水母和黄松峪水库嘉定桃花水母均为国内常见种[1]。实验中发现桃花水母性腺特征多变,缺乏精确的量化参数,是导致形态学分类中物种误定的关键原因[1,14,19]。有意义的是,基于ITS碱基序列多样性分析,为2个水库桃花水母物种鉴定提供了遗传学佐证[15-16,20]。嘉定桃花水母和索氏桃花水母在北京分布的信息为桃花水母属物种多样性格局研究提供了更丰富的生物地理特征数据。
ITS分析结果在确认怀柔水库索氏桃花水母的同时,将原记录于无锡鼋头渚和湖州安吉等江浙地区的信阳桃花水母同样归入索氏桃花水母。这一结果揭示出国内索氏桃花水母可能广泛分布于东部浅山地带到广阔平原地带。其分布范围基本覆盖了浙江、江苏、安徽、贵州、湖南、湖北、河南、山东、北京直至辽宁等有桃花水母记录的大部分地区[1,5,14-16]。适生生境既包括湖泊、水库、河流等自然水体,也包括公园水坑、水池甚至废弃铁锅等人工生境,表现出较高的环境耐受性[1,5,21-22]。
嘉定桃花水母北京黄松峪种群无疑为该物种纬度最高的分布记录。黄松峪种群和长江上游诸多种群均发生在山区水库等自然水体,表明其适生生境比索氏桃花水母单一。国内学者发表的其它杭州桃花水母的地理分布格局有待进一步确定分类地位后加以分析[1,5]。
3.2 桃花水母的环境适应力和环境指示价值
国内大多数报道都把桃花水母的发生与水质优良、水环境优越相联系,认为桃花水母是清洁水体的指示物种,但相关研究又很少描述和分析水域的水文和水质状况[1,14-16]。桃花水母对水环境的耐受性及其发生的临界条件尚缺乏充分的了解。本研究中,北京2个水库的桃花水母同时发生于2022年9月上旬,但2个库区的主要水质因子在此前后并未呈现明显的“水质优化”的趋势。对照地表水环境质量标准(GB 3838-2002),怀柔水库常年处于地表Ⅱ类水状态,而黄松峪水库则为地表Ⅲ类水,TN等指标甚至高于地表Ⅳ类水限值。常规水质指标TN、TP、BOD和DO等指标指示为中营养-轻度富营养水体。桃花水母对水质清洁的需求似乎并不明显。
Jankowksi等指明索氏桃花水母发生在富营养化水体[23],Caputo等揭示富营养化和大型浮游甲壳类丰度有利于水母生长和生境扩张[11,13],Marchessaux等认为富营养化和水色加深更有利于桃花水母的存活和扩张[10]。这些研究和本文的结果一样支持桃花水母更偏爱食物丰富的富营养水体而非清洁水体[22]。
上述的桃花水母生境类型多半也难以满足“清洁”水体的条件。诸如人工池塘、采石场、公园(社区)水池、屋顶水池甚至废弃铁锅等狭小水面、鱼类稀少、水流不畅的静水水体在气温较高的夏秋季难保水质“清洁”[1,5,21-23],而长江中下游的城市水体[5],以及江浙一带地表水体能维持“清洁”状态也是小概率事件[15]。因此,桃花水母生活水体的水环境条件差异巨大,既有清洁水体,也有富营养化水体,其中不乏劣于北京怀柔水库和黄松峪水库的生境。因此,桃花水母对水环境质量耐受范围较广,为广适性物种。相比于清洁水体,桃花水母更偏爱适口食物丰富的中营养水体。
3.3 桃花水母发生的环境驱动力
基于北京地区2个水库桃花水母发生条件和文献报道的桃花水母生境条件的总结,本研究结合动物种群调节及行为生态理论,提出桃花水母形成水母体行有性生殖的两个驱动力假说,借以探究桃花水母发生的环境条件。
1)温变胁迫效应(temperature shock effects)。不同于水质因子,水温和食源因素与桃花水母的发生相关明显。9月上旬,北京地表水体水温快速下降,黄松峪和怀柔水库表层水温降至25℃以下,降温胁迫很可能是刺激桃花水母转行有性生殖以应对后续不利低温的适应性对策。有性生殖优越于无性生殖的一个重要表现就是可以借助合子度过不良季节以拓展自己的分布范围,提升自身的环境适应力[24]。这种生殖方式的转换是低等水生生物应对环境变化的重要生活史对策(life history strategy)。当水温骤升或骤降时,无性世代的螅状体启动有性生殖形成水母体并进而形成合子以度过随之到来的干旱或严寒季节。桃花水母的地理分布范围表明其温度耐受范围较窄,对温度变化较为敏感。中国、日本、美洲、澳洲和中、西欧等温带水体是桃花水母的常见分布区,而近赤道的热带和南北极未见报道[1-5,14-16,21-23]。狭窄的分布带揭示出桃花水母对温度条件具有较高的敏感性,难以耐受热带水体高温,也不能耐受寒带冬季低温。因此,纬度较低的南方水体(尤其是浅水小水面)春季快速升温和秋季快速降温都可能诱发桃花水母发生,而北京等较高纬度的水体不具备夏季高水温条件,则秋季降温可能是桃花水母发生的重要驱动因素。
2)营养限制效应(nutrition limitation effects)。野生动物多在食源丰富的季节完成交配和生殖,从而提高生殖成功率[24]。桃花水母启动有性生殖后,其水母体快速增大、加之运动耗能,其食物摄取显著高于水螅体[22,25-27],且个体越大,食量越大[3]。食源充足才启动有性生殖有利于其有性生殖成功[23]。桃花水母以小型水生动物为食,其主食对象为0.4~1.4 mm的轮虫(Rotifera)、枝角类(Cladocera)和桡足类(Copepoda)等浮游动物,以及0.1~3.0 mm的底栖动物至鱼类幼体[22,27]。在种群较大时,桃花水母的摄食会导致浮游甲壳类象鼻溞(Bosminaspp.)和网纹溞(Ceriodaphniaspp.)等密度的显著下降[13,21,26]。水体生产力水平和喜食动物生物量高低对桃花水母有性生殖的时节应有决定性作用[22]。
富营养化水体夏秋季初级生产力水平较高,具有维持高密度浮游动物的条件。低纬度地区的浅水水体,春季回温较快,浮游植物和浮游动物的快速增长可以满足桃花水母发生的营养需求[21-23]。高纬度水体春季回温较慢,需到夏秋季方才具备食源丰富的营养条件[18,22]。北京地区冬春低温,水生动物休眠,生长季从5月底开始。表层水温18℃以上时大型浮游动物恢复生长并于夏秋季生物量达峰[16-17]。北京记录的桃花水母发生时间都在7月底至9月初,符合浮游动物等饵料生物丰度变动的季节性规律,支持营养限制的理论推断。
致谢:本文的水质和水生物采样调查和实验分析得到水生态组李世国、冯璧钧、梁琪琪、杨晓颖和郝天赐的大力协助,库区调查得到怀柔和黄松峪水库管理处大力支持,一并表示感谢。
猜你喜欢
农村青少年科学探究(2023年4期)2023-07-30
海南热带海洋学院学报(2021年5期)2021-11-07
森林工程(2021年4期)2021-08-23
儿童故事画报(2020年8期)2020-10-30
文苑(2019年14期)2019-08-09
海峡姐妹(2018年4期)2018-05-19
天天爱科学(2017年12期)2018-01-31
速读·中旬(2016年9期)2017-05-09
创新作文(小学版)(2017年23期)2017-04-04
四川师范大学学报(自然科学版)(2015年3期)2015-02-28
