
青藏高原东南缘高山湖泊藻类响应气候变化和大气沉降的长期特征*
2023-11-06 08:19陈光杰孔令阳陈小林黄林培
湖泊科学 2023年6期
朱 云,陈光杰,孔令阳,李 静,陈小林,李 平,马 欠,周 起,黄林培,任 雁
(云南师范大学地理学部,云南省高原地理过程与环境变化重点实验室,昆明 650500)
气候变化和持续增强的人类活动已成为影响湖泊环境变化的重要驱动因子[1-2],而在未受到人类活动直接影响的高纬度、高海拔地区,越来越多的证据表明高山湖泊出现了明显的生态系统变化[3-4]。监测记录显示,高山地区的增温效应较其他地区更为显著[5],如阿尔卑斯山地区自1980年以来增温幅度可以达到全球的2~4倍[6]。同时,化石燃料燃烧、化肥施用等产生的污染物质已通过大气环流输送到高海拔地区[7-8],成为影响高山湖泊环境与生态系统健康的重要环境压力[9]。高山湖泊是高山生态系统的重要组成部分,一般具有较低的生物生长速率、独特的物种生态位和单一的营养结构,其生态系统对气候变化和营养输入的响应十分敏感[4,10]。大量研究表明,气候变化和大气污染沉降首先会改变高山湖泊水体的理化特征,进而对藻类等生物生长和群落结构产生直接影响[11-12]。其中,增温会直接造成湖泊水温升高、热力分层时间延长、水位波动增强等[13]。Rühland等研究表明,增温导致深水湖泊热力分层加强后有利于湖中小型藻类(如小环藻)占优[14]。持续的增温还会导致高山湖面冰封时间减少、光照条件改善,均有利于浮游藻类的繁殖[15]。同时大气氮沉降过程持续输送了氮素等营养盐,在贫营养水平的高山湖泊中会显著促进藻类的生长,甚至会改变水体营养盐的配置并从氮限制转变为磷限制[16]。而在酸中和能力弱的高山湖泊中,大气氮沉降还会造成水体酸化并导致湖泊生物量减少、生物多样性下降[17-18]。
在区域增温和大气沉降的背景下,流域特征、地质背景、湖盆形态等类型不同的高山湖泊中表现出了差异的生态响应模式[19-20]。位于树线以上的湖泊通常具有流域侵蚀作用强、水体溶解有机碳(DOC)浓度低等特点,而较低的DOC难以有效地阻挡紫外线辐射并保护水生生物[21],因此位于树线以上的高山湖泊生物群落的变化要可能比树线以下的高山湖泊更为显著[20]。当大气活性氮沉降到地面后,具有面积小、植被覆盖低、土壤层发育薄特点的流域可以在较短的时间内达到氮饱和,随后流域多余的活性氮会直接输入湖泊,在此类流域中的高山湖往往对大气氮沉降的响应更为敏感[9]。而在环境特征相似的流域中,小型湖泊水体的混合作用较强[22]、生物多样性低[23],会对气温变化和营养盐输入做出快速响应[19],呈现出比大型湖泊更为敏感的响应模式[22,24]。青藏高原东南缘地区是我国增温幅度最显著的地区之一[25],区域内分布着大量的小型高山湖泊且多为冰川遗迹湖。监测结果显示,南亚和东南亚等地区的大气污染物质可通过印度夏季风输送到我国西南地区,青藏高原东南缘地区的平均氮(N)沉降速率约为40 kg/(hm2·a),是我国陆地N沉降速率的2倍左右[9,26]。已有研究表明,该地区的大气沉降等过程已对高山湖泊的环境特征和藻类演替产生了重要影响[17,23]。同时已有研究将高山湖泊在藻类响应模式上存在的区域异质性主要归咎于流域特征差异,而对于湖泊类型在其中的影响作用研究仍较少[9,20,23]。
基于以上分析,本研究选择青藏高原东南缘地区位于树线以下、具有不同水深的3个小型弱碱性湖泊为研究对象,通过钻孔测年、理化特征与藻类等多指标的时间序列分析,首先重建过去300年来3个高山湖泊的环境变化与藻类响应历史。进一步结合区域气候定量重建和氮沉降等数据,对比分析在气候变化和大气沉降背景下高山湖泊藻类响应的模式异同,以期获得该地区湖泊藻类变化的同步性特征及水深等湖泊类型的调节作用,可为全球变化持续的情景下预测高山湖泊生态系统变化的未来趋势提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原东南缘地区,盖公错纳、沃迪错、碧沽天池3个高山湖泊均分布在云南省迪庆藏族自治州香格里拉县。其中,盖公错纳海拔4214 m、距离香格里拉市约40 km,沃迪错海拔3842 m、距离香格里拉市约52 km,碧沽天池海拔3840 m、距离香格里拉市约25 km (图1a)。3个高山湖泊都位于树线以下(目前林线高度约4300 m),盖公错纳和沃迪错流域土地类型以裸地为主,碧沽天池流域土地类型以森林为主(表1)。盖公错纳最大水深约为39.4 m,水体呈弱碱性(pH=8.03),透明度为1.1 m且营养水平低,采样时水生植物分布极少(覆盖度<1%);流域面积为5.61 km2,湖泊面积为0.14 km2(流域和湖泊面积之比约为40.1)。盖公错纳缺乏入湖河流,出水口位于湖盆东部,湖泊水位高时有出流,为开放型湖泊。沃迪错最大水深约为20.7 m,水体清澈,透明度达10.1 m,水体呈弱碱性(pH=8.12),采样时水生植物分布极少(覆盖度<1%);流域面积为3个湖泊中最大,达36.04 km2,湖泊面积为0.17 km2(流域和湖泊面积之比约为212)。沃迪错湖盆西侧分布有2条入湖河流,两个湖区之间相连通有湖水交换,有1条出湖河流分布于湖盆东侧,为开放型湖泊。碧沽天池最大水深约为1.7 m,清澈见底,水体呈弱碱性(pH=7.94),营养水平低,湖中有少量水生植物分布;流域面积为6.41 km2,湖泊面积为0.16 km2(流域和湖泊面积之比约为40.1)。湖泊无河流输入且无出水口,为封闭型湖泊。

表1 盖公错纳、沃迪错和碧沽天池基本环境特征Tab.1 Key environmental characteristics of Gaigong Cuona, Wodi Co and Bigu Tianchi
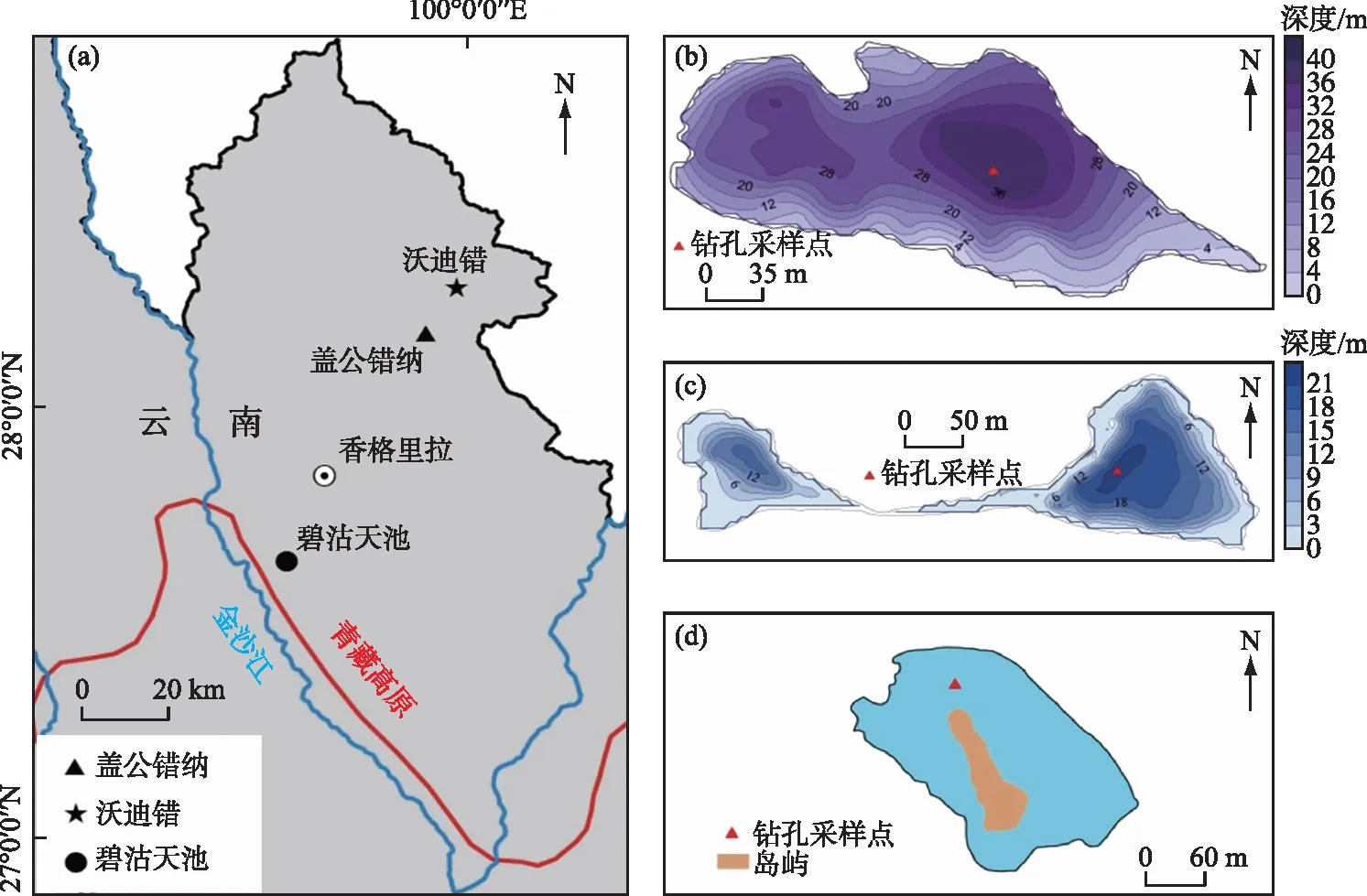
图1 湖泊地理位置(a),盖公错纳(b)、沃迪错(c)和碧沽天池(d)等深图及采样点位置Fig.1 Maps showing lake location (a), bathemetry and sampling sites of Gaigong Cuona (b), Wodi Co (c) and Bigu Tianchi (d)
3个湖泊所在区域主要受西南季风和南支西风的影响。冬半年(11-4月),受南支西风急流的影响,气候偏冷干;夏半年(5-10月)受西南季风的控制,降水充沛,气候温暖湿润。香格里拉市气象站点(海拔3450 m; 数据来源于国家气候中心http://ncc.cma.gov.cn/cn/)近62年来的温度和降水气象数据显示,6-9月温度最高,12-2月温度低于0℃;年降水量为596.3 mm,主要集中在6-9月,占全年降水量的77% (图2a)。
1.2 样品采集
2019年8月利用Hon-Kajak沉积物重力钻在盖公错纳湖心39.4 m深处(图1b),采集长度为112 cm的沉积物柱状样品。现场对其顶部10 cm按0.5 cm间隔,10 cm以下按1 cm间隔进行分样,盖公错纳钻孔共获得122个样品。2019年8月利用Hon-Kajak沉积物重力钻在沃迪错最深处(图1c),采集长度为41 cm的沉积物柱状样品。2018年7月利用Hon-Kajak沉积物重力钻在碧沽天池最深处(图1d),采集长度为100 cm的沉积物柱状样品。现场对钻孔顶部5 cm按0.5 cm间隔,5 cm以下按1 cm间隔进行分样,沃迪错钻孔共获得46个样品,碧沽天池钻孔共获得105个样品,将3个湖泊样品分别装进自封袋中低温密封避光保存。本文根据年代模型结果,重点分析了盖公错纳钻孔上部18 cm长度、沃迪错钻孔上部41 cm长度、碧沽天池钻孔上部47 cm长度的样品。湖泊钻孔采样时,均在现场使用多参数水质监测仪YSI对湖泊水温、pH、溶解氧(DO)浓度和电导率等进行测量,并采集湖泊水样低温保存带回实验室进行总磷(TP)和总氮(TN)等水化学指标分析。
1.3 实验室分析
实验室内对沉积钻孔进行了210Pb和137Cs年代测试,使用美国Canberra公司生产的高纯锗探测器测定放射性核素210Pb和137Cs比活度,应用CRS模型(the constant rate of supply model)建立了沉积物样品年代序列[27-28]。分析的沉积物理化和藻类代用指标包括粒度组成、TN含量、氮同位素(δ15N)、藻类色素、硅藻等。
称取0.3~0.5 g沉积物干样,经前处理去除碳酸盐和有机质后,使用马尔文激光粒度仪(MS-2000)测量沉积物粒度,本研究主要使用粒度数据中的中值粒径。沉积物TN含量及δ15N的测定使用FLASH HT Plus元素分析仪及流动质谱仪(MAT-253)完成,精度分别为±1.0%和<0.1‰。沉积物色素含量测定参考Swain的方法[29]: 称取0.4 g沉积物干样放入50 mL聚乙烯离心管中,加入90%丙酮25 mL,摇匀后静置10 h以上,用离心机在3500 r/min速度下离心10 min,将上清液过滤至100 mL容量瓶。同样操作反复萃取3次,将萃取液都过滤到容量瓶中,最后用90%丙酮定容至100 mL取10 mL萃取液,使用岛津UV-1750紫外可见分光光度计测定666 nm波长的吸光值,即为叶绿素的特征吸收峰。在样品比色皿中加入2滴稀硝酸,酸化2 min,在同一波长处测量吸光值,即为自然叶绿素的吸光值。使用参考文献中提供的公式计算叶绿素(CD)含量和自然叶绿素(NC)含量。CD含量可以用来指示湖泊的初级生产力变化[29],而NC含量是指未分解的叶绿素,它指示了沉积物中的色素保存程度[30]。
沉积物硅藻样品的分析参照标准方法进行处理[31]。主要步骤包括:称取0.3 g的沉积物干样于烧杯中,加入10%的盐酸去除碳酸盐,反应一段时间后加入30%的双氧水并在80℃的水浴锅中加热4 h去除有机质,处理好的样品倒入离心管用离心机在1200 r/min的速度下离心4 min,吸去上清液加入蒸馏水继续离心4次至样品呈中性,离心好的样品用Naphrax@树胶制片。硅藻属种的鉴定在徕卡显微镜(Leica DM2500)1000倍油镜下完成,鉴定方法参考的是Krammer和Lange-Bertalot[32]的分类系统,每个样品计数不少于400个。所有实验均在云南省高原地理过程与环境变化重点实验室完成。
1.4 气象监测和气候重建数据收集
1.5 数据分析
首先应用限制性聚类分析(CONISS)方法对硅藻地层图进行聚类分析[37],并使用Broken-stick模型识别硅藻群落结构显著变化的阶段。应用去趋势对应分析(DCA)确定硅藻群落变化梯度的最大长度(SD),3个湖泊DCA主轴1长度都小于2,故选用主成分分析(PCA)的方法识别硅藻群落变化的主要特征,使用Broken-stick模型对主成分分析的主轴进行选取。应用冗余分析(RDA)的方法识别驱动硅藻群落变化的主要环境梯度,分析前对环境数据进行标准化处理,选择显著性(P<0.05)的环境因子,剔除其中膨胀系数(variance inflation factor,VIF)大于20的环境因子并生成RDA排序图。进一步开展方差分解分析用于3个高山湖泊的驱动因子及其强度比较,方差分解分析中将驱动群落变化的关键因子分为3类环境梯度,其中粒度、重建降水等代用指标指示水文变化,沉积物TN反映氮沉降等作用下营养盐的影响,温度重建指标指示气温梯度。将筛选的环境因子用于简约模型选择并生成韦恩图(Venn),从而定量识别不同环境因子对硅藻群落的相对驱动强度。本研究中,聚类分析、PCA、RDA、方差分解等在R 4.1.0中完成,沉积物地层图在C2 1.7.2中完成。
2 结果
2.1 岩芯年代序列
根据沉积钻孔210Pb、137Cs比活度和CRS模型建立了3个高山湖泊的钻孔年代序列(图3)。盖公错纳沉积钻孔中210Pb的比活度从表层到底部总体呈下降趋势,137Cs比活度峰值出现在10.5 cm位置。根据CRS深度-年代模型建立盖公错纳钻孔的年代学序列,结果表明17.5 cm深度的钻孔代表了过去约293 a;沉积速率出现从钻孔底部向上先增加而在深度3.75 cm以上部分开始下降的长期特征,平均沉积速率为0.15 cm/a。沃迪错沉积钻孔中210Pb的比活度从表层到底部总体呈减少趋势,137Cs比活度峰值出现在11.5 cm深度的位置。根据CRS深度-年代模型建立了沃迪错钻孔的年代学序列,结果表明40.5 cm深度的钻孔覆盖了过去约277 a的沉积序列;沉积速率呈现持续增加的趋势,平均沉积速率为0.18 cm/a。碧沽天池沉积钻孔中210Pb 的比活度从表层到底部总体呈减少趋势,137Cs比活度在13.5 cm深度的位置达到峰值。根据CRS深度-年代模型建立了该钻孔的年代学序列,结果表明46.5 cm深度的钻孔覆盖了过去约321 a的沉积序列;沉积速率呈现先缓慢增加后快速增加的趋势,平均沉积速率为0.22 cm/a。
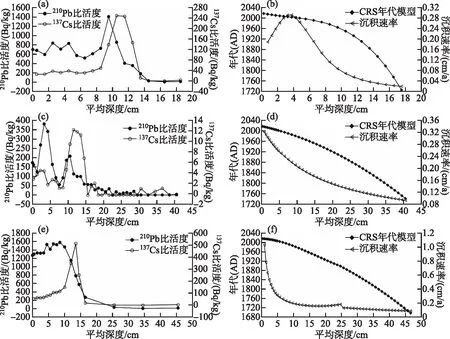
图3 盖公错纳(a,b)、沃迪错(c,d)、碧沽天池(e,f)钻孔210Pb和137Cs比活度、CRS年代模型和沉积速率Fig.3 Depth profiles showing specific activities of 210Pb and 137Cs, CRS age model and sedimentation rate for sediment cores of Gaigong Cuona (a,b), Wodi Co (c,d) and Bigu Tianchi (e,f), respectively
2.2 3个湖泊沉积物主要理化指标与藻类色素变化特征
近300年来,3个高山湖泊沉积物主要理化指标与藻类色素等记录均存在较为明显的变化(图4)。盖公错纳沉积物主要理化指标与藻类色素的结果显示(图4a),盖公错纳沉积钻中值粒径变化范围为6.3~10.4 μm,在2005年之前中值粒径呈现缓慢减小的趋势,自2005年后中值粒径出现快速减小。沉积物TN含量在5.0~9.7 mg/g(均值±标准差为(7.3±1.0) mg/g),TN含量变化具有明显的阶段性特征,1953年之前TN含量变化较小,基本保持在7.6 mg/g左右,1953-1988年TN含量上升,呈高浓度,1989-2019年TN含量下降,呈低浓度。盖公错纳沉积钻叶绿素浓度在0.17~0.83 μg/g org ((0.37±0.17) μg/g org),叶绿素含量总体呈上升趋势,显著上升期出现于1991年后。自然叶绿素表现为从钻孔底部到表层逐渐增加的趋势。
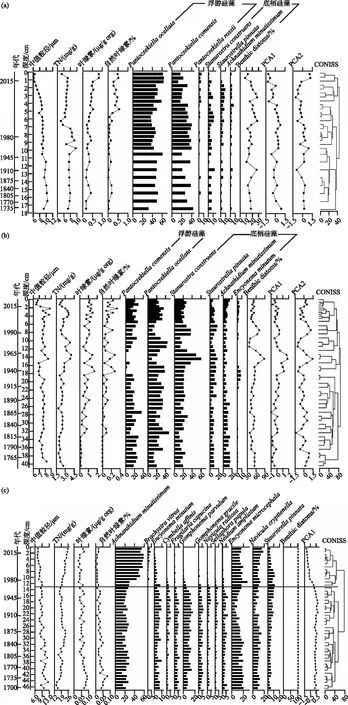
图4 盖公错纳(a)、沃迪错(b)和碧沽天池(c)钻孔理化指标、藻类色素、优势硅藻属种(相对丰度>5%)组合、硅藻群落PCA样品得分的地层变化图及聚类分析结果Fig.4 Stratigraphic profiles showing long-term variations in key physico-chemical proxies, algal pigments, dominant diatoms (species with relative abundance >5%), diatom PCA scores with clustering analysis results for Gaigong Cuona (a), Wodi Co (b) and Bigu Tianchi (c), respectively
沃迪错沉积物主要理化指标与藻类色素的结果显示(图4b),沃迪错沉积钻中值粒径变化范围为6.5~8.8 μm,近300年来中值粒径呈现波动变化,没有明显的阶段性特征。沉积物TN含量为1.8~4.0 mg/g((2.7±0.5) mg/g),近300年来TN含量总体呈上升趋势,在1956-1983年期间,TN含量出现快速上升形成一个高浓度阶段。沃迪错沉积钻叶绿素含量为0.11~1.71 μg/g org ((0.76±0.44) μg/g org),叶绿素含量总体呈上升趋势,在1938-1962年期间,叶绿素含量出现快速上升。沃迪错沉积钻自然叶绿素呈现从钻孔底部到表层逐渐增加的趋势。
碧沽天池沉积物主要理化指标与藻类色素的结果显示(图4c),碧沽天池沉积钻中值粒径变化范围为7.2~12.6 μm,近300年来中值粒径呈持续减少的趋势。沉积物TN含量在13.8~25.6 mg/g ((20.0±2.7) mg/g), TN含量呈现先增加后减少最后增加的趋势,1968年后碧沽天池TN含量开始快速上升。碧沽天池沉积钻叶绿素浓度在0.03~0.09 μg/g org ((0.06±0.02) μg/g org),叶绿素浓度总体呈下降趋势。碧沽天池沉积钻自然叶绿素呈现从钻孔底部到表层逐渐减少的趋势。
2.3 3个湖泊的硅藻群落组合及其变化特征
沉积物记录显示,3个高山湖泊中硅藻群落的优势物种及其变化特征存在明显差异(图4)。水深达39.4 m 的盖公错纳硅藻群落以浮游种为主(占比达82%),水深达20.7 m的沃迪错硅藻群落浮游种和底栖种各占约50%,水深达1.7 m的碧沽天池硅藻群落则以底栖种为主,占比几乎达99%。聚类分析(CONISS方法)结果显示,近300年来盖公错纳和沃迪错硅藻群落变化无明显阶段性特征,而碧沽天池自1968年后硅藻群落结构发生明显转变,硅藻种极细微曲壳藻(Achnanthidiumminutissimum)相对丰度上升成为最大优势种,硅藻种微小异极藻(Gomphonemaparvulum)和小肉丝藻(Encyonemaminutum)相对丰度出现下降。盖公错纳钻孔中共鉴定出硅藻21个属,95个种;优势种为眼斑小环藻(Pantocsekiellaocellata)、科曼小环藻(Pantocsekiellacomensis);近300年来底栖硅藻比呈现波动变化。沃迪错钻孔中共鉴定出硅藻23个属,118个种;优势种为眼斑小环藻(Pantocsekiellaocellata)、科曼小环藻(Pantocsekiellacomensis)、连结脆杆藻(Staurosiraconstruens);底栖硅藻比在1951-1968年和1987-1996年2个时期出现上升。碧沽天池钻孔中共鉴定出硅藻16个属,78个种;优势种为极细微曲壳藻(Achnanthidiumminutissimum)、小头桥弯藻(Encyonopsismicrocephala)等;近300年来湖泊一直以底栖硅藻为主。
硅藻群落数据的主成分分析(PCA)和零模型检验结果表明,盖公错纳和沃迪错硅藻群落数据共识别出2个PCA主轴显著,碧沽天池硅藻群落数据识别出1个PCA主轴显著。相关分析结果显示(图5),盖公错纳硅藻PCA1得分变化与中值粒径变化显著正相关(r=0.56,P<0.01),沃迪错硅藻PCA1得分变化与TN变化显著正相关(r=0.52,P<0.001)。碧沽天池硅藻PCA1得分变化与TN变化显著负相关(r=-0.38,P<0.01),与中值粒径变化显著正相关(r=0.30,P<0.05),与气温变化显著负相关(r=-0.38,P<0.05)。
2.4 主要环境压力及驱动强度识别
为了识别影响藻类变化的主要环境梯度及其驱动强度,将主要环境因子分组进行方差分析(图6)。其中粒度组成、降水定量重建数据指示了湖泊水动力、降水等水文过程,沉积物TN浓度反映营养水平的影响,温度定量重建指标指示了气温梯度。方差分解的结果显示(图6),驱动3个湖泊硅藻群落变化的环境因子和驱动强度存在差异,且碧沽天池硅藻群落的方差分析解释量最高。其中,TN的独立作用对沃迪错(6.5%)的影响强度均高于碧沽天池(4.8%)和盖公错纳(3.9%),水文波动的独立影响主要出现在碧沽天池(4.5%)和盖公错纳(3.5%),气温的独立作用仅在碧沽天池(4.8%)中较为明显,而TN、水文、气温3类因子的相互作用在碧沽天池(4.1%)和盖公错纳(2.7%)中较强。初级生产力的长期变化上,盖公错纳中气温的独立驱动作用较强(8.9%)且与水文因子的相互作用最明显(28.1%);TN的独立作用在沃迪错中最为显著(25.2%),且与气温呈显著的相互作用(17.5%);在浅水型的碧沽天池中TN和水文因子均产生了较强的独立影响(11.9%和7.6%)及相互作用(19.3%),且TN、水文、气温3类因子的相互作用在碧沽天池中也最为显著(5.5%)。

图6 过去300年来盖公错纳(a,d)、沃迪错(b,e)和碧沽天池(c,f)中主要环境梯度驱动硅藻群落(a,b,c)和叶绿素浓度(d,e,f)的方差分解结果Fig.6 Variation partitioning results showing the proportion of three key environmental gradients in explaining the temporal variance of diatom assembalges (a,b,c) and chlorophyll pigments (d,e,f) for the past 300 years in Gaigong Cuona (a,d), Wodi Co (b,e) and Bigu Tianchi (c,f), respectively
3 讨论
3.1 3个湖泊硅藻组合特征及驱动因素的对比分析
在不同水深梯度下,3个高山湖泊的硅藻群落组合存在明显差异。碧沽天池水深为1.7 m左右,硅藻群落以底栖种为主,其中硅藻种A.minutissimum是该湖最大的优势种。硅藻种A.minutissimum是常见的广布种,在3个高山湖中都有出现,它能耐受较宽的环境梯度且对于水温波动和营养水平变化都有很高的适应性[38-39]。盖公错纳硅藻群落以浮游种为主(占比达82%),且小型Pantocsekiella为优势种属。当气温上升、湖泊热力分层加强时,小型Pantocsekiella通常更具物种竞争优势[14]。同时水中光照条件也是影响小型Pantocsekiella生长的一个重要因素,透明度上升和湖面冰封持时缩短都可能导致小型Pantocsekiella丰度上升[15]。盖公错纳水深为39.4 m左右,是3个高山湖泊中最深且热力分层最为稳定的水体,为小型Pantocsekiella生长提供了适宜的水动力和生境条件。沃迪错中底栖硅藻和浮游硅藻约各占一半,底栖硅藻优势种主要为S.construens和Staurosirellapinnata,2个硅藻种生态习性相似,它们都能耐受较宽的环境梯度,在不同的水温梯度和营养梯度下都能很好的生存[15,40],所以在高山湖泊中较为常见[15,23]。在同为深水型湖泊的盖公错纳和沃迪错中,优势硅藻种属相似但在硅藻群落的组合上存在一定差异。硅藻种S.construens和Staurosirellapinnata都是体型比较小的底栖硅藻,这类硅藻在水流扰动下常可以悬浮在水体中[41],所以在深水湖泊盖公错纳和沃迪错中2个硅藻种都属于优势硅藻。但在沃迪错中硅藻种S.construens和Staurosirellapinnata的占比要高于盖公错纳,沃迪错水深为20.7 m左右、沿岸带面积大于盖公错纳,同时沃迪错湖水透明度较高(10.1 m),这为底栖硅藻生长提供了良好的条件。
深水与浅水湖泊的对比分析发现,硅藻群落的长期演替呈现了不同的阶段性特征。如在2个深水湖泊中硅藻群落近300年来无明显的阶段性变化特征,而碧沽天池自1968年后出现了显著的群落结构转变。方差分解结果显示(图6),驱动3个高山湖泊硅藻群落变化的环境因子存在较大差异。盖公错纳的硅藻群落变化主要受到水文变化和TN的影响,气温主要通过与其他环境因子的共同作用驱动了硅藻群落变化。盖公错纳硅藻以浮游的小型Pantocsekiella占多,小型Pantocsekiella的生长主要受到热力分层、光照、营养盐的共同限制[14-15,42]。水动力变化会影响湖泊热力分层的稳定性,气温上升则会加强湖泊热力分层的厚度和持续时间[13],水文和气温因子可能主要通过湖泊热力分层的变化来影响盖公错纳的硅藻群落构建。20世纪以来,青藏高原东南缘地区的气温加速上升[33],加强了该区域深水湖泊的热力分层,有利于小型Pantocsekiella属硅藻继续占优,这可能是2个深水型湖泊中硅藻群落结构未发生明显转变的部分原因。此外,Pantocsekiella属种组合变化还可能与营养盐物质的增多有关。湖泊添加实验表明,小型Pantocsekiella只有在N充足的情况下才能响应湖泊热平衡的变化[42]。沃迪错硅藻PCA1得分与TN变化呈显著正相关(r=0.52,P<0.001),表明了TN对物种组成的驱动作用。硅藻种S.construens和P.comensis是沃迪错的2个优势种,其中S.construens作为广布种可以在营养水平较高的水体中生存[38],而P.comensis是在水体营养水平较低的情况下常见的贫营养种[43]。1956-1968年期间随着TN含量上升,同期沃迪错的硅藻种S.construens相对丰度出现上升、P.comensis相对丰度出现下降,反映了硅藻群落对营养盐变化的快速响应。同时,硅藻种S.construens可能还受到气温波动的直接影响。已有研究发现,气温下降导致的湖面冰封时间延长时,耐低光的S.construens会更具生存优势[15,44]。因此1956-1968年的降温时段(图2b) 可能延长了沃迪错的湖面冰封时间,从而促进了硅藻种S.construens相对丰度上升。而在浅水湖泊碧沽天池,硅藻群落变化同时受到水文变化、TN、气温三者的共同影响,分析发现硅藻群落PCA1得分变化与TN变化呈显著负相关(r=-0.38,P<0.01)。硅藻种A.minutissimum作为碧沽天池的优势种,通常能耐受较宽的环境梯度,尤其对水温和营养水平变化有较强的适应性[38-39]。因此,1968年开始研究区的温度上升(图2b)和水体营养盐的升高(图4c)可能均促进了硅藻种A.minutissimum相对丰度上升。同时硅藻种A.minutissimum喜好附着生长在水生植物上[45],区域增温等作用直接促进了水生植物的生长和分布,从而有利于A.minutissimum等底栖物种的增加。
3.2 3个湖泊初级生产力变化特征及驱动因素的对比分析
青藏高原东南缘地区的结果显示,不同水深的高山湖泊初级生产力变化特征存在差异,且浅水湖泊中沉积物色素可能更容易受到降解作用的影响。沉积物叶绿素指标可以用来恢复湖泊初级生产力的变化历史[29],盖公错纳和沃迪错钻孔叶绿素浓度总体呈上升趋势(图4a和图4b),可能反映了湖泊初级生产力的上升。近300年来碧沽天池钻孔叶绿素浓度则总体呈现持续下降的趋势(图4c),可能反映了沉积物色素在浅水湖泊中受降解作用的影响较强。大量研究表明,藻类色素的保存和分解程度会受到光照、温度、氧气等环境条件的直接影响[30,46-47]。在热力分层较为稳定的深水湖泊中,沉积物-水界面通常会形成一个低光、低温、缺氧的底部环境,从而有利于色素保存且不易分解[46]。自然叶绿素可以用来反映沉积物中叶绿素的降解程度[30],在浅水湖泊碧沽天池中自然叶绿素数值较低且呈下降的趋势,反映了沉积物叶绿素降解作用的持续增强。因此在浅水湖泊中,沉积物-水界面通常出现高光照、溶解氧水平高且水温波动较大的底部环境,均不利于沉积物色素的保存[47-48]。
方差分析结果表明(图6),驱动3个湖泊初级生产力变化的环境因子也存在较大差异。如影响盖公错纳生产力长期变化的环境因子主要为气温和水文条件,而沃迪错的藻类生产力变化与TN和气温有关。本研究中的高山湖泊都具有较长的冰封期,气温上升可以提高水温和延长生长季[49],促进湖泊藻类生长和初级生产力上升。水文变化会影响深水湖泊的热力分层状况,进而影响湖泊藻类的生长。稳定的热力分层有利于表层漂浮藻类获得充足的光照[49],同时热力分层增强还会减弱上下层水体的营养交换[13,49]。而在浅水型的碧沽天池中水温波动更为快速、沉积物-水界面溶解氧水平更高,因此沉积物色素受到降解作用影响更为明显。方差分解结果可能更多反映的是影响色素降解的环境因素,如气温上升会提高水温、缩短湖泊冰封期并增加光照时长,进而加速色素降解,水文波动频繁会影响湖泊的混合状态和溶解氧水平而对色素降解造成更大影响[46]。
3.3 青藏高原东南缘地区高山湖泊藻类响应大气沉降和气候变化的模式差异
在气候变化持续的背景下,青藏高原东南缘地区高山湖泊的营养水平波动与区域大气沉降过程显著相关。现代监测记录表明,青藏高原地区存在着明显的大气沉降过程与持续增强的长期特征[50]。研究结果的对比分析发现(图7),3个高山湖泊的沉积物δ15N信号自工业革命以来均出现偏负变化的总体趋势,且与该地区太极湖等高山湖泊的δ15N和Hg浓度指标记录的氮沉降过程和大气传输模式[17]基本一致。同时上述沉积物记录的大气沉降过程也与格陵兰[36]、喜马拉雅山[35]等地区冰芯记录的大气沉降模式具有较好的同步性(图7),表明了大气沉降作用是3个高山湖泊氮素持续输入的重要来源[8-9]。该研究区域的已有研究表明,高山湖泊氮素输入和营养水平变化主要受到大气氮沉降作用的长期影响[9,17]。而盖公错纳和沃迪错钻孔δ15N信号在后期的偏正趋势,可能是受到内源湖泊藻类增加等的影响[17]。初步结果表明,盖公错纳现生浮游植物的δ15N值大致在3.9‰~5.0‰。同时除少部分大气活性氮直接沉降到湖泊水体外,绝大部分大气活性氮首先沉降到流域地表,然后通过地表径流等过程最终汇入湖泊。因此,高山湖泊沉积物的δ15N信号会同时受到大气沉降、流域地表和湖泊生物地球化学等多个过程的综合影响[23]。

图7 盖公错纳、沃迪错和碧沽天池钻孔δ15N记录(本研究)、太极湖钻孔δ15N和Hg含量变化[17]、格陵兰冰芯δ15N和硝酸盐浓度[36]和喜马拉雅山冰芯硝酸盐浓度[35] 变化的对比分析Fig.7 Comparative profiles showing temporal changes of sediment δ15N in Gaigong Cuona, Wodi Co and Bigu Tianchi (this study), sediment δ15N and Hg in Taiji Lake[17], ice-core δ15N and nitrate concentrations in Greenland[36] and ice-core nitrate concentrations in Himalayan[35]
在低纬高原地区,高山湖泊藻类对环境变化的响应模式呈现出了与水深等湖泊形态显著相关的变化特征。本研究结果表明,3个高山湖泊中氮营养盐输入对硅藻群落的驱动作用总体强于气温的影响。对比分析发现,近300年来深水湖泊中硅藻群落变化无明显阶段性特征,而浅水湖泊中硅藻群落结构自1968年后发生明显转变。同时,驱动硅藻群落变化的环境因子在3个湖泊之间存在一定差异。如随着湖泊水深的下降,TN等营养因子对硅藻群落的驱动作用逐渐增强,且3个湖泊中浅水湖泊碧沽天池硅藻群落对环境变化的响应最敏感。在湖泊面积相近的背景下,水深更浅意味着湖容更小,对气温的缓冲作用和营养盐输入的稀释作用更弱,藻类对环境变化的响应可能更为敏感[19,22],指示了硅藻群落转变的环境阈值可能与水深等湖泊类型相关。初级生产力变化也表现出和硅藻群落类似的湖泊差异性。深水湖泊盖公错纳和沃迪错初级生产力总体呈上升趋势,而碧沽天池由于水深较浅其沉积物色素受到降解作用的影响更强,出现了叶绿素浓度下降的长期趋势。此外,水深较浅的沃迪错初级生产力开始上升的时间(约1938年)要早于水深较深的盖公错纳(1991年左右),且驱动盖公错纳和沃迪错初级生产力变化的环境因子也存在差异。如2个湖泊初级生产力的变化都受到气温和水文波动的影响,但在水深更浅的沃迪错TN对初级生产力的驱动作用变得更强;沃迪错在3个湖泊中具有最大的流域和湖泊面积之比,可能出现了较强的流域TN输入负荷和较高的水体营养水平(表1)。因此,湖泊类型与流域特征可能对高山湖泊的藻类演替特征产生了较强的调节作用,从而导致了生态系统响应模式在湖泊间呈现出显著差异。
4 结论
在区域增温和大气沉降作用显著的青藏高原东南缘地区,高山湖泊的藻类响应模式出现了与湖泊和流域特征相关的异质性特征。近300年来,2个深水湖泊(盖公错纳和沃迪错)硅藻群落变化无明显阶段性特征,而浅水湖泊碧沽天池硅藻群落结构自1968年后发生明显转变。进一步分析发现,盖公错纳中硅藻群落变化主要受到水文波动影响,而沃迪错和碧沽天池中硅藻群落变化主要受到TN影响,且TN对浅水湖泊硅藻群落的驱动作用更为显著。同时,盖公错纳和沃迪错初级生产力总体呈上升趋势,且沃迪错初级生产力开始上升的时间要早于盖公错纳;而浅水湖泊碧沽天池叶绿素浓度总体呈持续下降趋势,可能反映了区域增温下增强的藻类色素降解作用。驱动初级生产力变化的环境因子也呈现明显的湖泊差异,其中盖公错纳受水文和气温影响、沃迪错与TN和气温有关,而碧沽天池与水文波动和TN相关。由此可见,高山湖泊藻类对区域环境变化的响应特征和环境阈值可能与水深梯度、流域面积等明显相关,而浅水湖泊的藻类指标对区域增温和大气沉降作用可能具有更高的敏感性。
猜你喜欢
大自然探索(2023年7期)2023-11-14
课堂内外(小学版)(2023年9期)2023-10-11
椰城(2022年5期)2022-05-18
书香两岸(2020年3期)2020-06-29
黑龙江水利科技(2020年8期)2020-01-12
少儿美术(快乐历史地理)(2019年4期)2019-08-27
小学生作文(中高年级适用)(2018年6期)2018-07-09
阅读(低年级)(2018年4期)2018-05-14
上海建材(2018年1期)2018-04-18
宝藏(2018年1期)2018-01-31
