
Lnc46198半番鸭胚胎期特异性表达及对鸭胚成纤维细胞增殖和凋亡的影响
2023-11-09 14:09章琳俐辛清武缪中纬朱志明朱春红邱君志黄勤楼郑嫩珠
中国农业大学学报 2023年11期
李 丽 章琳俐 辛清武 缪中纬 朱志明 朱春红 邱君志 黄勤楼 郑嫩珠*
(1.福建省农业科学院 畜牧兽医研究所/福建省畜禽遗传育种重点实验室,福州 350013;2.福建省农业科学院,福州 350013;3.中国农业科学院 家禽研究所,江苏 扬州 225125;4.福建农林大学 生命科学学院,福州 350002)
半番鸭又称骡鸭,是家鸭与番鸭的属间杂交产物,具有很强的杂种优势,如抗逆性强、生长速度快等,但半番鸭不育,没有实际种用价值。若想获得半番鸭苗,只能饲养其父母本并采用人工授精技术,但亲本间自然交配受精率低,仅为15%~45%,极大限制了半番鸭产业发展。属间杂交不育是一个非常复杂的性状,其中雄性不育可表现为性腺发育缺陷,如精索或睾丸发育不全。除了父母本染色体差异及生理生化影响,半番鸭性腺发育可能由多个基因控制。因此半番鸭性腺发育异常研究可为探究属间杂交不育机理提供理论参考依据。
长链非编码RNA(Long non-coding RNA,lncRNA)虽未直接编码蛋白,但可在多种生物学过程中发挥作用[1-4],并主要通过靶基因或miRNA途径影响多种生物学过程,如Wang等[5]研究了鸡卵巢中与性成熟杂种优势相关的lncRNA和mRNA的系统图谱。Jiang等[6]发现lncPGCR/miR-6577-5p/Btrc途径可促进鸡原始生殖细胞(Primordial germ cells,PGCs)生长发育。lncPGCAT-1靶向ILF3激活JNK信号通路可促进鸡PGCs形成[7]。
前期研究中,李丽等[8]通过测序发现TCONS_00246198(简称lnc46198)在不育半番鸭性腺中特异性高表达,推测lnc46198可能靶向17β-HSD3基因抑制生殖细胞发育进而影响半番鸭胚胎期性腺发育。然而,尚未有lnc46198对鸭细胞分化发育调控机制的相关研究。因此,本研究拟测定lnc46198在半番鸭不同发育时期胚胎性腺中的表达规律及在半番鸭不同组织中表达情况,同时构建过表达载体初步分析lnc46198对鸭胚成纤维细胞的增殖和凋亡的影响,旨在探索lncRNA功能及对半番鸭性腺发育的影响,可为半番鸭生殖细胞发生机制提供基础资料,为半番鸭种质创新提供指导。
1 材料与方法
1.1 试验材料
鸭胚成纤维细胞购自湖南丰晖生物科技有限公司,过表达载体(pcDNA3.1-3 Flag质粒)和MEM α购自美国英杰生命技术有限公司,Trypsin-EDTA(0.25%)、PlatinμmTMPCR SμperMix High Fidelity、GeneArt®Seamless Cloning and Assembly Enzyme Mix、LipofectamineTM 3000 Transfection Reagent、TRIzol®Plus RNA Purification Kit和SuperScriptTMIII First-Strand Synthesis SuperMix均购自美国赛默飞世尔科技公司,QIAqμick PCR &Gel Cleanμp Kit购自德国凯杰公司,感受态细胞DH5α购自上海唯地生物技术有限公司。
Power SYBR®Green PCR Master Mix购自德国应用生物系统公司,Cell Counting Kit-8购自日本同仁化学研究所,Annexin-V-FITC Apoptosis Detection Kit购自sigma-aldrich(上海)贸易有限公司,PowerUpTMSYBRTMGreen Master Mix购自德国应用生物系统公司,TUNEL BrightRed Apoptosis Detection Kit购自美国诺唯赞生物科技股份有限公司。
1.2 半番鸭鸭胚lnc46198特异性表达分析
收集番鸭和北京鸭人工授精获得的半番鸭鸭胚,雌雄鉴定后选择雄性样本,在体视显微镜下采集雄性半番鸭10、15、20和25 d不同胚龄的性腺组织,同时采集12.5 d胚龄大脑、心脏、肝脏、肺脏、肌肉、肾脏和睾丸。提取总RNA,逆转录后cDNA贮存在-20 ℃备用。利用RT-qPCR技术测定半番鸭不同组织和不同时期lnc46198相对表达量。根据lnc46198测序序列,内参基因为GAPDH,引物序列:F:GGAGCTGCCCAGAACATTATC,R:GCAGGTCAGGTCCACGACA。采用Primer Premier 6.0和Beacon designer 7.8软件设计定量PCR引物,由生工生物工程(上海)股份有限公司合成。lnc46198引物序列:F1:CGGCTGATTCCGTTATGGGTAT,R1:CAGGGCTTCCTCCACCTTT,扩增长度98 bp,退火温度60 ℃。20 μL反应体系:cDNA 1.0 μL,SDW 8.0 μL,Power SYBR®Green Master Mix 10.0 μL,正向引物/10 μmol/L 0.5 μL,反向引物/10 μmol/L 0.5 μL,反应条件:95 ℃ 1 min;95 ℃ 15 s,63 ℃ 25 s,40个循环。每个样品重复3次,相对表达水平以2-ΔΔCt进行统计分析。
1.3 lnc46198-pcDNA3.1-3 Flag重组载体构建
以半番鸭性腺组织提取RNA,后逆转录成cDNA,以cDNA为模板设计引物,lncRNA-F和lncRNA-R为引物,引物序列:lncRNA-F:TTGGTACCGAGCTCGGATCCGTTAGATGATGTTTAGTTAGTTTAA,下划线代表BamHI酶切位点,lncRNA-R:CTGTGCTGGATATCTGCAGAATTCCAGAAATAAGAGCCTGTTTAA,下划线代表EcoRI酶切位点,扩增长度3 525 bp,退火温度60 ℃。对lncRNA CDS序列进行PCR扩增获得序列,50 μL扩增反应体系:SDW 1.0 μL,lncRNA-F (10 μmol/L) 1.0 μL,lncRNA-R (10 μmol/L) 1.0 μL,cDNA2.0 μL,Platinμm®PCR SμperMix,High Fidelity 45 μL,反应条件:预变性94 ℃,2 min,94 ℃,30 s,60 ℃,30 s,68 ℃,3.5 min,35个循环。In Fusion进行载体和目的片段的重组,PCR扩增产物采用QIAqμick PCR &Gel Cleanμp Kit进行切胶回收(具体参照试剂盒说明书),然后采用GeneArt®Seamless Cloning and Assembly Enzyme Mix进行重组载体的无缝克隆。lncRNA和pcDNA3.1-3 Flag融合10 μL反应体系:SDW 2.0 μL,lncRNA Purified product 2.0 μL,pcDNA3.1(BamHI/EcoRI)(50 ng)1.0 μL,GeneArt®2X Enzyme Mix 5.0 μL,反应条件:室温放置20 min;冰上放置2~3 min,然后全部转化感受态细胞DH5α,挑选单克隆菌落进行摇菌培养,筛选阳性菌液送生工生物工程(上海)股份有限公司进行重组载体测序。
1.4 鸭胚成纤维细胞转染试验
1.4.1鸭胚成纤维细胞培养和传代
鸭胚成纤维细胞采用MEM α培养液(含10% FBS以及双抗)进行复苏,在37 ℃、5% CO2条件下进行培养至汇合度90%左右;加入Trypsin-EDTA(0.25%)进行消化,以5×105个/mL细胞密度接种6孔板进行传代。
1.4.2lnc46198过表达质粒的细胞转染
鸭胚成纤维细胞以5×105个/mL密度接种6孔板,在37 ℃、5% CO2孵育箱中过夜;待细胞汇合度约为70%时,换成新鲜完全培养液,按照LipofectamineTM 3000说明书分别转染lnc46198-pcDNA3.1过表达质粒以及pcDNA3.1对照质粒,8 h后更换完全培养液;转染48 h后收集细胞提取总RNA用于后续qPCR分析过表达效率。
1.5 RT-qPCR检测lnc46198过表达效率
收集过表达后的DEF细胞和对照细胞,采用TRIzol®Plus RNA Purification Kit提取总RNA,由DNase I去除残余的基因组DNA,然后按SuperScriptTMIII First-Strand Synthesis SuperMix说明书合成cDNA。以GAPDH为内参,采用PowerUp SYBRTM Green Master Mix试剂盒进行RT-qPCR扩增。引物序列:F2:CAAATCTAGGCAACACCACA,R2:CAGCAACAATCATAGGAGGC,扩增长度94 bp,退火温度60 ℃。20 μL反应体系:cDNA 4.0 μL,SDW 5.0 μL,Power SYBR®Green Master Mix 10.0 μL,正向引物/10 μmol/L 0.5 μL,反向引物/10 μmol/L 0.5 μL,反应条件:95 ℃ 1 min;95 ℃ 15 s,63 ℃ 25 s,40个循环,每个试验重复3次,每个样品设置3个复孔,其相对表达量采用2-ΔΔCt法进行计算。
1.6 CCK8法检测过表达lnc46198对鸭胚成纤维细胞增殖的影响
增殖和凋亡是细胞功能研究的2个基本的研究现象,且两者也可以相互应验。将转染pcDNA3.1对照质粒以及lnc46198-pcDNA3.1过表达质粒24 h后的鸭胚成纤维细胞进行消化收集,细胞计数并调整细胞密度。按每孔2×103/100 μL细胞密度接种96孔板,待细胞贴壁后记为0 h,继续培养48 h加入10 μL CCK8,在37 ℃、5% CO2条件下孵育2 h后用酶标仪检测450 nm处的吸光值,每组均进行6复孔重复。
1.7 流式细胞术分析过表达lnc46198对鸭胚成纤维细胞细胞凋亡的影响
为确定过表达lnc46198对鸭胚成纤维细胞准确的凋亡率,本试验将转染pcDNA3.1对照质粒以及lnc46198-pcDNA3.1过表达质粒48 h后的鸭胚成纤维细胞进行消化收集,预冷PBS漂洗细胞2次。加入300 μL预热的无EDTA-胰酶消化3~5 min,然后加入预热 200 μL含血清培养基终止反应;同时加入500 μL PBS清洗残余的细胞进入到2 mL离心管。离心(1 500 g,5 min),弃去上清;加入1 mL预热PBS轻轻吹打重悬,离心同上;再次加入1 mL PBS重悬,离心同上。每管加入500 μL Binding Buffer,轻轻重悬细胞。先加入5.0 μL Annexin V-FITC和10 μL PI,颠倒混匀,室温避光放置5~15 min,然后进行流式细胞分析。
2 结果与分析
2.1 lnc46198在半番鸭胚龄时期性腺和不同组织中的表达
lnc46198在半番鸭胚龄时期性腺中特异表达结果如图1所示,结果表明,在所有检测胚龄性腺中均出现表达,且随着胚龄增加lnc46198在性腺中相对表达量有升高趋势,表明lnc46198具有时间特异性。lnc46198在半番鸭12.5 d胚龄性腺和不同组织中特异性表达结果显示(图2),lnc46198在半番鸭12.5胚龄性腺中的表达量极显著高于其他组织(P<0.01),推测lnc46198可能在鸭性腺中发挥作用。
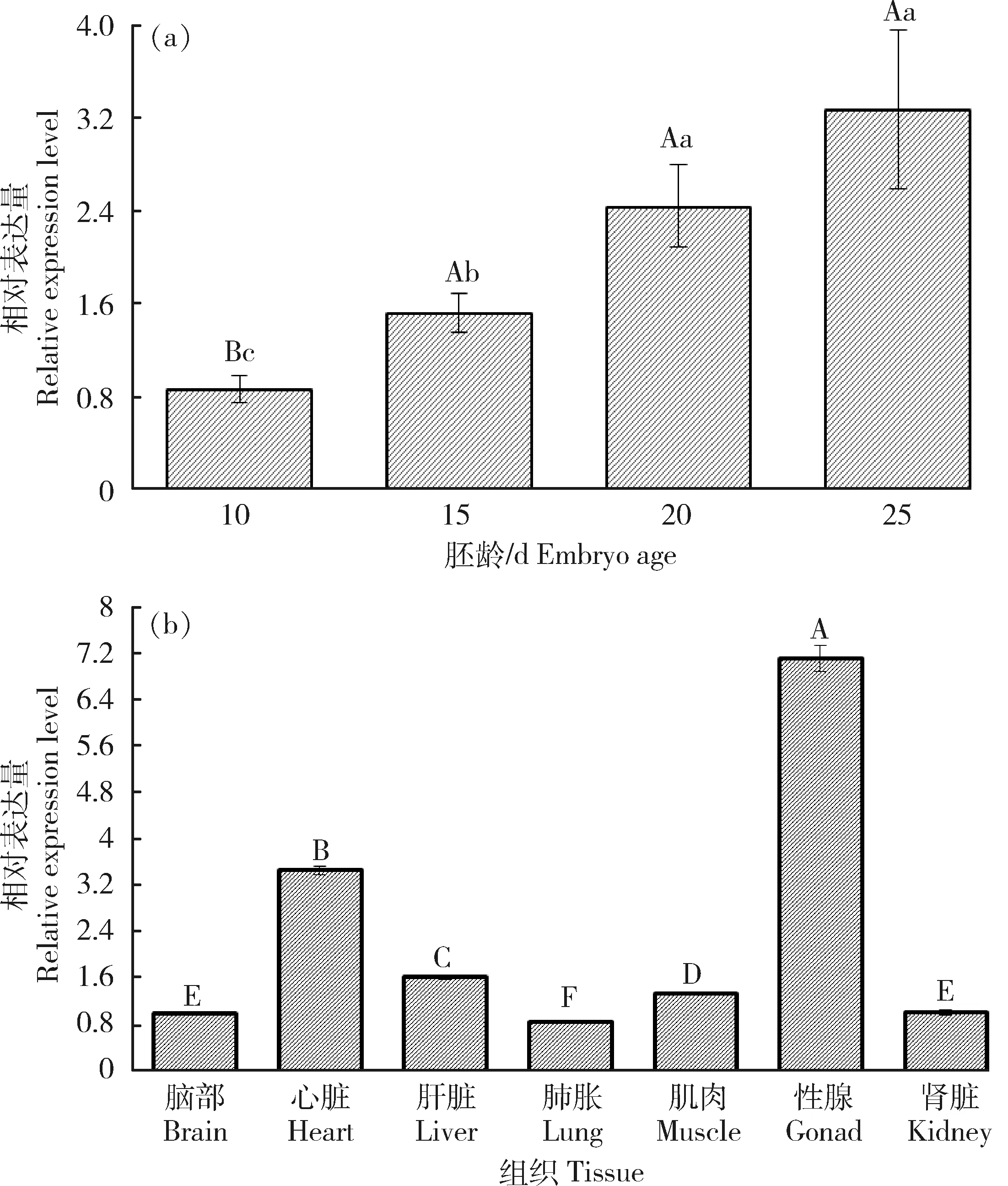
(a)半番鸭不同胚龄性腺;(b)半番鸭不同组织。

M:DL5K DNA marker(100、250、500、750、1 000、2 000、3 000、5 000 bp)
2.2 lnc46198-pcDNA3.1-3 Flag重组载体构建
BamHI和EcoRI酶双酶切割lncRNA片段,采用全基因合成,lnc46198全长插入到pcDNA3.1-3 Flag,lncRNA PCR扩增序列结果如图2(a)所示,lnc46198 PCR扩增产物约3 525 bp。重组载体测序结果如图2(b)所示,结果显示重组载体含有序列与lnc46198一致,表明重组载体构建成功。
2.3 lnc46198过表达分析
对鸭胚成纤维细胞系进行培养和传代,传至第三代细胞。鸭胚成纤维细胞转染lnc46198-pcDNA3.1-3 Flag过表达载体,转染后48 h lnc46198过表达效率结果如图3所示,与转染对照组pcDNA3.1相比,lnc46198的表达水平上调了29.28倍(P<0.01),表明lnc46198过表达成功。
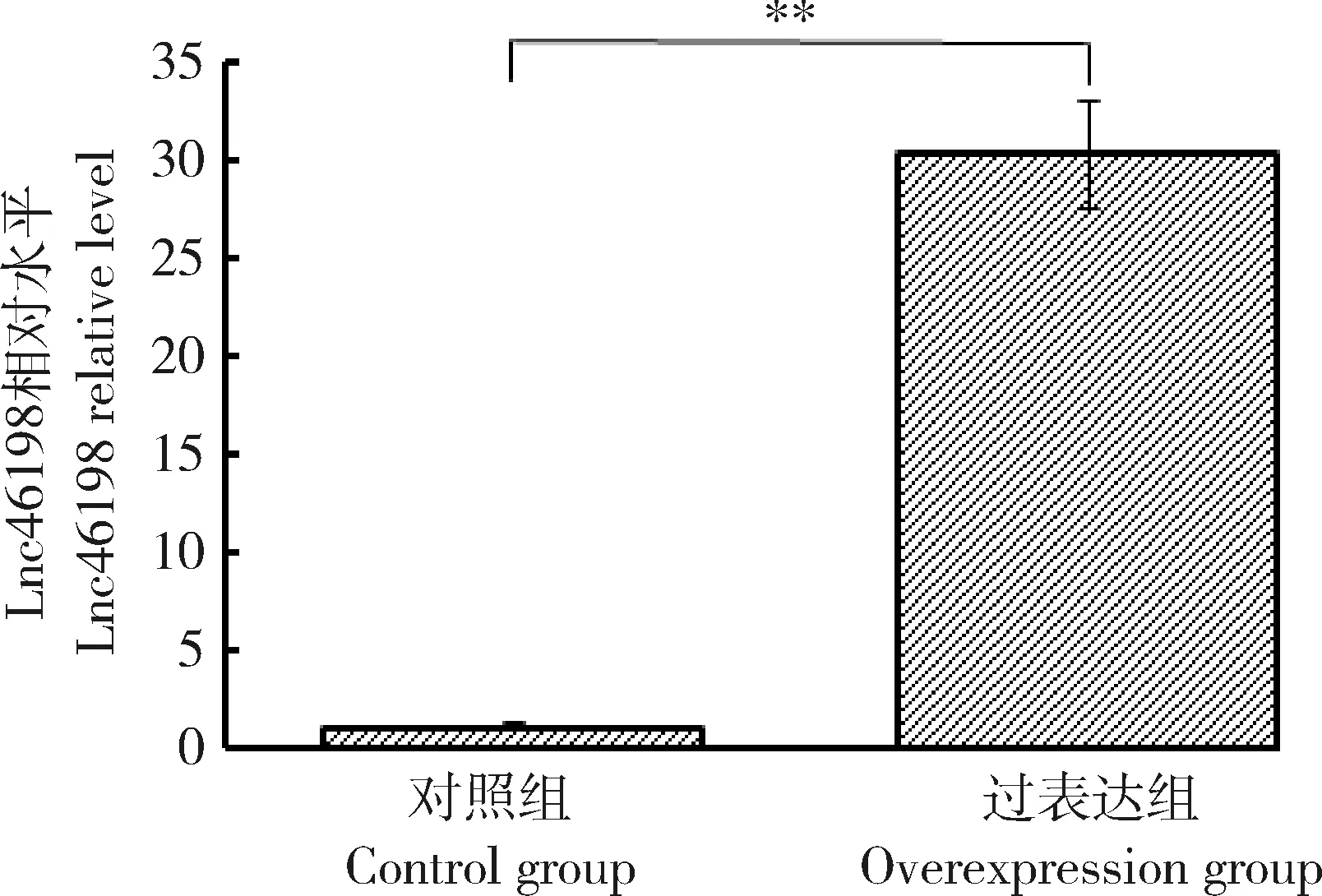
**P<0.01表示差异显著。下同。
2.4 过表达lnc46198对鸭胚成纤维细胞增殖活力的影响
为探究lnc46198对鸭胚成纤维细胞的影响,转染pcDNA3.1对照质粒以及lnc46198-pcDNA3.1过表达质粒,CCK8法检测过表达lnc46198后鸭胚成纤维细胞的活力情况。结果显示(图4),与空白组以及pcDNA3.1组比较,过表达该lnc46198后,鸭胚成纤维细胞活力极显著降低(P<0.01)。表明过表达lnc46198可抑制鸭胚成纤维细胞的增殖活力。
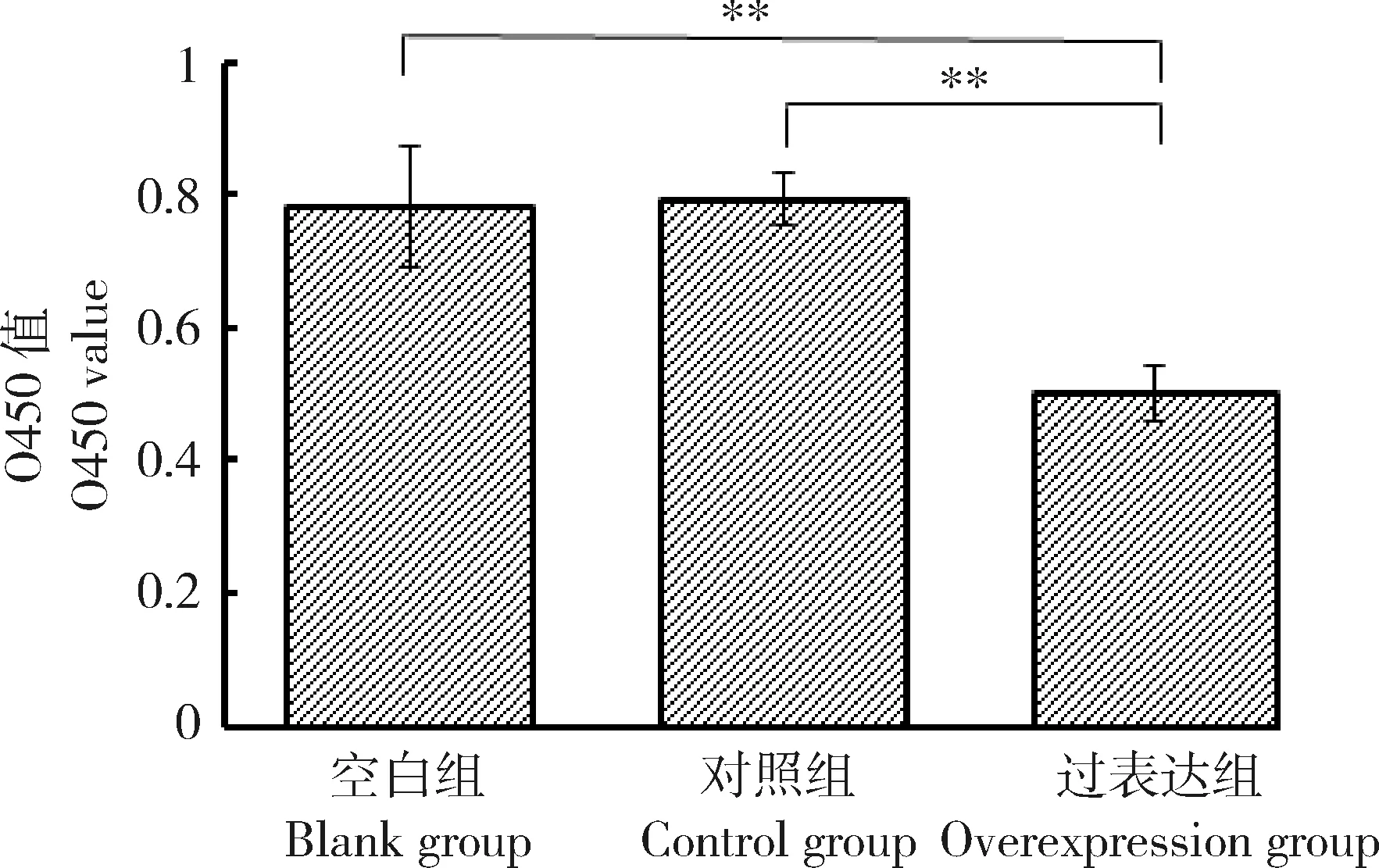
图4 过表达lnc46198对鸭胚成纤维细胞增殖活力的影响
2.5 流式细胞分析过表达lnc46198对鸭胚成纤维细胞凋亡的影响
为探究lnc46198对鸭胚成纤维细胞凋亡的影响,鸭胚成纤维细胞转染pcDNA3.1对照质粒以及lnc46198-pcDNA3.1过表达质粒,利用Annexin V-FITC流式检测过表达lnc46198后鸭胚成纤维细胞的凋亡情况,结果显示(图5(a)),与pcDNA3.1对照组相比,过表达lnc46198后,鸭胚成纤维细胞凋亡增多,过表达组鸭胚成纤维细胞凋亡率是pcDNA3.1对照组的7.30倍(图5(b),P<0.01),表明过表达lnc46198可促进鸭胚成纤维细胞凋亡。
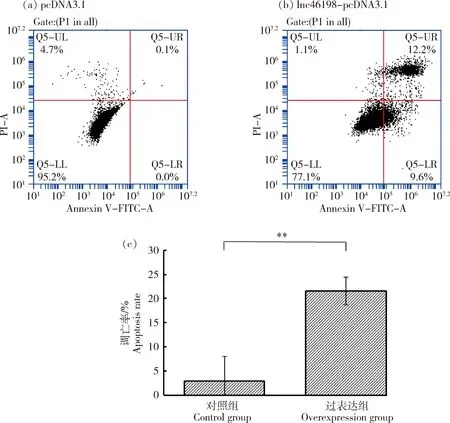
(a)、(b)细胞凋亡散点图;(c)细胞凋亡率。
3 讨 论
大多数lncRNAs在组织分化发育过程中,都具有明显的组织或时间表达特异性。Wichman等[9]研究表明lncRNA动态表达对小鼠精子发生和生育发挥潜在作用。lncRNAs失调可能造成雄性低精子数或不育症。Liu等[10]研究发现lncRNA在城口山地鸡胚胎中期E12的表达模式和其他时间点均不同。Bao等[11]发现雄性种系发育过程6个关键时间点lncRNA的动态表达调控。本研究对半番鸭胚胎不同发育时期性腺中lnc46198的表达量进行检测分析,结果发现,在所有检测胚龄性腺中lnc46198均有表达,且随着胚龄增加lnc46198的表达量也逐渐升高,表明lnc46198具有较高的时间特异性。这与夏雪平等[12]研究一致,夏雪平等发现lncRNA 54770.30在黄鳝卵巢至精巢转变过程中表达量呈上升趋势,提示lncRNA 54770.30可能参与黄鳝性腺发育过程,lncRNA的表达规律可为研究其功能提供参考依据。同时,本研究还测定了lnc46198在半番鸭不同组织中的表达,结果显示lnc46198在半番鸭性腺中的表达量极显著高于其他组织,即在性腺中特异性表达,推测lnc46198可能在半番鸭性腺发育中发挥作用,这与前人的研究结果相近,如黄子妍等[13]认为MSTRG.15568.9与SPAG4基因具有较明显的组织表达特异性,并靶向SPAG4基因参与雄性精子发生与精子活力调控;Jastrzebski等[14]则发现多个lncRNA在火鸡输精管、睾丸和附睾中表达,并认为其可能参与雄性性腺发育。以上研究结果提示lncRNA表达具有组织特异性。另外,本研究中lnc46198的时间和组织特异表达趋势与缪中纬等[15]研究中17β-HSD3基因的组织特异性相近,二者表达模式一致,提示lnc46198可能与靶基因17β-HSD3共同参与生物学过程,如影响睾酮合成或睾丸间质细胞发育[16-18]。
Hu等[19]发现lncRNA TCONS_00814106可通过miR-1343调控猪颗粒细胞的增殖和凋亡,Wu等[20]认为lnc13814通过apla-miR-145-4途径促进鸭颗粒细胞的凋亡,Liang等[21]研究也表明lncRNA在小鼠精子发生特定阶段与基因表现出协同变化,上述研究均表明lncRNAs可通过miRNA或转录后水平调控基因功能。lncRNA可通过调控基因上游调控机制参与动物性腺或胚胎发育,Spga-lncRNA[22]和Gm2044[23]等研究发现关键lncRNA Gm2044可抑制小鼠精原细胞增殖进而参与小鼠精子发生过程。Zou等[24]认为lncRNAs的特异性剂量补偿机制可能导致鸡卵巢不对称发育。Zhang等[25]发现lnc5926调控胚胎基因组激活基因(Embryonic genome activation,EGA)表达进而影响山羊早期胚胎发育。因lnc46198为新筛选的未知lncRNA,需根据靶基因功能预测lncRNA功能,本研究中lnc46198靶基因为17β-HSD3,研究表明17β-HSD3基因是类固醇激素合成途径中的关键基因,其对睾丸激素合成和雄性繁育性状至关重要[26-27],若缺陷可能造成性腺发育障碍[28-30],因而推测lnc46198靶向17β-HSD3基因参与半番鸭性腺发育。lncRNA异常表达则可影响细胞增殖、分化或凋亡作用,本研究过表达lnc46198后发现其可抑制鸭胚成纤维细胞增殖活力,并促进鸭胚成纤维细胞凋亡,进一步验证了lnc4619抑制细胞增殖的作用。Hu等[31]认为lncRNA-Gm2044在转基因小鼠精原细胞中过表达不会影响睾丸形态和生育能力,但会阻断精子生成,并可作为诊断不育的分子靶标。Liang等[32]研究认为过表达lncRNA-Gm2044或敲除miR-335-3p可以降低转录因子A-MYB对突触复合体蛋白1基因(Synaptonemal complex protein 1,SYCP1)表达和细胞增殖的影响。同时,lncRNA还可能影响其他类型细胞的增殖或凋亡活动,Sun等[33]研究表明lnc90386敲除可显著抑制鸡胚成纤维细胞凋亡和炎症因子,并促进鸡胚成纤维细胞增殖。杨光等[34]发现了一条新的lncRNA lncbMD,结果表明干扰lncbMD后能够促进牛骨骼肌卫星细胞的增殖,孟珊等[35]通过干扰试验发现猪肌肉组织中显著高表达的lncRNA-6617可促进猪肌内前体脂肪细胞分化,上述研究与本研究结果均表明干扰、过表达或敲除lncRNA可影响细胞活动,但其作用方式不尽相同。本研究初步分析lnc46198对细胞增殖或凋亡的作用,然而,lnc46198对生殖细胞分化、精子发生及减数分裂的作用机制尚不清楚,后续将分析lnc46198表达水平与半番鸭胚胎性腺发育异常之间的效应关系,以进一步确定lnc46198靶向17β-HSD3在鸭生殖细胞增殖或分化过程中的功能,可为lncRNA对半番鸭生殖细胞分化发育作用机制提供参考依据,对半番鸭不育研究具有重要意义。
4 结 论
lnc46198在不同胚龄半番鸭性腺中动态表达,在半番鸭12.5 d胚龄性腺中特异性高表达,并抑制鸭胚成纤维细胞增殖,促进鸭胚成纤维细胞凋亡,可为lncRNA对鸭生殖细胞分化发育作用机制提供科学参考依据。
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21
昆明医科大学学报(2021年8期)2021-08-13
云南医药(2021年3期)2021-07-21
食品科学(2018年10期)2018-05-23
中国现代医学杂志(2015年26期)2015-12-23
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年33期)2015-03-01
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
食品工业科技(2014年15期)2014-03-11
