
A novel vanadium complex VO(p-dmada) inhibits neuroinflammation induced by lipopolysaccharide
2023-11-18 09:51ZhijunXioqinLiHujiZhngXinLiuShungxuHnAnwrAbdurhmnLimingShnXiuboDuNnLiXiodYngQiongLiu
Chinese Chemical Letters 2023年10期
Zhijun H, Xioqin Li, Huji Zhng, Xin Liu, Shungxu Hn,Anwr Abdurhmn, Liming Shn, Xiubo Du,c, Nn Li,f,*, Xiod Yng, Qiong Liu,c,*
a Shenzhen Key Laboratory of Marine Biotechnology and Ecology, College of Life Sciences and Oceanography, Shenzhen University, Shenzhen 518055, China
b National R&D Center for Se-rich Agricultural Products Processing, Hubei Engineering Research Center for Deep Processing of Green Se-rich Agricultural Products, School of Modern Industry for Selenium Science and Engineering, Wuhan Polytechnic University, Wuhan 430023, China
c Shenzhen Bay Laboratory, Shenzhen 518055, China
d State Key Laboratories of Natural and Biomimetic Drugs, Department of Chemical Biology, School of Pharmaceutical Sciences, Peking University Health Science Center, Beijing 100191, China
e Department of Pharmacy, Shenzhen People’s Hospital (The Second Clinical Medical College, Jinan University; The First Affiliated Hospital, Southern University of Science and Technology), Shenzhen 518020, China
f Shenzhen-Hong Kong Institute of Brain Science-Shenzhen Fundamental Research Institutions, Shenzhen 518055, China
Keywords:Vanadium Vanadyl N-(p-N,Nd(V im O(ept-hdyml aamdian)o)phenylcarbamoylmethyl)iminodiacetate NMei c urroogi n li fa lammation Proteomics Peroxisome proliferator activated receptor gamma (PPARγ)Uncontrolled microglial activation is decisively involved in the neuroinflammatory pathogenesis of brain diseases.Consequently, suppression of microglial overactivation appears to be a strategy for the prevention of nerve injury.In this paper, a novel vanadium complex, vanadyl N-(p-N,Ndimethylaminophenylcarbamoylmethyl)iminodiacetate (VO(p-dmada)), was synthesized from vanadyl sulfate and N,N-dimethyl-p-phenylenediamine, which was structurally characterized by Fourier transform infrared spectrum and ESI-MS analysis.The effect of VO(p-dmada) on neuroinflammation was investigated by using the models of lipopolysaccharide (LPS)-induced BV2 microglial cells and BALB/c mice.Our data demonstrated that VO(p-dmada) significantly suppressed microglial activation by downregulating inflammatory mediators and associated proteins, and inactivating nuclear factor-κB (NF-κB) signaling pathway.VO(p-dmada) also upregulated peroxisome proliferator activated receptor gamma (PPARγ) by reducing transglutaminase 2 and heat shock protein 60 expression.Co-treatment with PPARγ antagonist GW9662 significantly impeded the inhibitory effect of VO(p-dmada) on LPS-induced neuroinflammation.These cumulative findings demonstrated that VO(p-dmada) is a potential new drug for the treatment of neuroinflammation-related neurodegenerative diseases.
ABSTRACT
Neuroinflammation mediated by overactivated glial cells is a common pathological feature of neurodegenerative diseases [1,2].Microglia, acting as immune cells in the central nervous system,play a critical role in regulating the neurotoxicity caused by inflammatory response [3–5].Under pathological conditions, microglia can be overactivated and in turn result in overproduction of many inflammatory mediators including nitric oxide (NO), prostaglandin E2(PGE2), and pro-inflammatory cytokines such as tumor necrosis factor-α(TNF-α), interleukin-6 (IL-6) and interleukin-1β(IL-1β)[6].These inflammatory cytokines can collectively lead to eventful neurotoxicity and aggravated progress of neurodegenerative diseases [4].Accordingly, the control of aberrant activation of microglia is regarded as a potentially powerful strategy for preventing neurodegenerative diseases.
Neuroinflammation can be induced by a range of pathogens,such asβ-amyloid (Aβ), viruses and lipopolysaccharide (LPS) [7–9].These pathogenic stimuli could activate nuclear factor-kappa B (NF-κB) signaling pathway bothin vivoandin vitro[7–9].NFκB, a vital transcription factor that regulates inflammatory response, is sequestered in the cytoplasmviacombining with inhibitory kappa B (IκB) in the resting microglia.LPS can activate the IκB kinase (IKK) which in turn phosphorylates IκB and leads to its ubiquitination and further degradation by the proteasome,and eventually results in translocation of NF-κB from cytoplasm to the nucleus.Once in the nucleus, NF-κB acts as a nuclear transcription factor to regulate the expression of inflammatory mediators including TNF-α, IL-6, IL-1β, inducible nitric oxide synthase(iNOS) and cyclooxygenase-2 (COX-2) [7,10].Therefore, inactivation of NF-κB signaling is a promising strategy for treatment of neuroinflammation-based diseases.
Peroxisome proliferator activated receptor gamma (PPARγ), a vital ligand-activated nuclear transcription factor, plays a critical role in regulating energy balance, insulin sensitization, neuronal differentiation, and inflammation [11–14].When activated,PPARγimmediately binds to the PPAR-response element and directly induces or represses transcription of downstream target genes [15].Decreased expression of PPARγhas been observed in LPS-stimulated microglial cells and mice [16,17].PPARγactivators have been found to regulate inflammatory response in several models of central nervous system (CNS) disorders [7,18].Accumulated evidences showed that PPARγactivation can suppress the release of proinflammatory cytokines through the inhibition of NF-κB signaling [19].Some vanadium compounds, such as bis(ethylmaltolato)oxidovanadium(IV) (BEOV), have been reported as PPARγmodulators and thus as potential agents for the treatment of diabetes and cerebral ischemia [20–22].
Metal ions are indispensable for living organism.Metal-based coordination compounds have attracted extensive attention for diagnosis and treatment of various diseases [22–25].Vanadium, a biological trace element with some unique features, has also received a great deal of attention from the researchers in various areas including chemistry, biology, toxicology and pharmacology [26].VanadylN-(p-N,N-dimethylaminophenylcarbamoylmethyl) iminodiacetate (VO(p-dmada)), a novel vanadium complex with low toxicity and high bioavailability, is synthesized from vanadyl sulfate andN,N-dimethyl-p-phenylenediamine in our laboratory (Fig.S1A in Supporting information).Thep-H2dmada ligand and VO(p-dmada)complex were structurally characterized using Fourier transform infrared (FT-IR) and nuclear magnetic resonance (NMR) spectroscopies and mass spectrum (MS) (Figs.S1B-F in Supporting information).In a preliminary study, VO(p-dmada) showed better anti-diabetic effect than other vanadium compounds [27].In this study, we endeavored to explore the effect of VO(p-dmada)on neuroinflammation.Thus, BV2 microglial cells were initially treated with VO(p-dmada) to find out its optimum concentration by CCK-8 assay.As shown in Fig.S2 (Supporting information), VO(p-dmada) did not display any cytotoxic effect on BV2 cells and LPS-induced BV2 cells at doses ranging from 0 to 10 μmol/L.
LPS is a common toxicant for microglial activation [9,28,29].During neuroinflammatory processes, NO and PGE2, two critical proinflammatory mediators, are produced by activated microglia[30].Thus, the suppression of NO and PGE2emancipation in LPSstimulated BV2 microglia were measured to evaluate the antiinflammatory effects of VO(p-dmada).As shown in Fig.S3A (Supporting information), the levels of released NO and PGE2in the culture medium were dramatically elevated after stimulation with LPS.Treatment with VO(p-dmada) significantly inhibited the excessive secretion of NO and PGE2by LPS-stimulated BV2 microglia.Since the production of NO and PGE2is regulated by iNOS and COX-2, respectively, inhibition of iNOS and COX-2 expression could be a primary strategy for decreasing inflammatory response [31].Thus, the expression of iNOS and COX-2 were examined by Western blot and immunofluorescence analysis.The data in Figs.S3BE and Fig.S4 (Supporting information) indicated that iNOS and COX-2 protein levels were dramatically decreased by VO(p-dmada)treatment.The mRNA level of iNOS was also decreased significantly by VO(p-dmada) treatment (Fig.S5A in Supporting information).Further investigation showed that VO(p-dmada) reduced the levels of NO, iNOS and COX-2 proteins in a time-dependent manner, when it was incubated with the cells for 4, 8, 16 and 24 h,respectively (Fig.S6 in Supporting information).
TNF-α, IL-12 and IL-6 were important pro-inflammatory cytokines released by activated immune cells [3,4,32,33].Vanadium compounds have been reported to regulate the production of proinflammatory cytokines [7,21].To ascertain whether VO(pdmada) can affect the production of inflammatory cytokines, the levels of TNF-α, IL-12 and IL-6 were analyzed by ELISA.As shown in Fig.S3F (Supporting information), TNF-α, IL-6 and IL-12 levels were increased dramatically in the culture media of LPS-stimulated BV2 microglia when compared with unstimulated control, while VO(p-dmada) treatment obviously inhibited the secretion of inflammatory cytokines.These data suggest that VO(p-dmada) suppresses LPS-induced inflammatory responsesviadownregulating the expression of iNOS and COX-2 and inflammatory cytokines.
Overproduction of proinflammatory mediators in LPSstimulated BV2 microglia was primarily regulated by NF-κB signaling pathway [34].To further evaluate whether the antiinflammatory effect of VO(p-dmada) was associated with NF-κB signaling pathway, the expression levels of IκB-α,p-IκB-α, p65 andp-p65 were analyzed by Western-blot in LPS-stimulated BV2 microglia.As shown in Figs.S3G and H (Supporting information), the phosphorylation levels of IκB-αand p65 were distinctly augmented in LPS-stimulated BV2 microglia compared with unstimulated control.Interestingly, VO(p-dmada) treatment significantly reduced the phosphorylation levels of these proteins.Immunofluorescent staining was used to reveal the cellular distribution of p65.The fluorescent images showed that VO(p-dmada)treatment markedly blocked NF-κB/p65 nuclear translocation in LPS-stimulated BV2 microglia (Figs.S3I and J in Supporting information).The mRNA level of p65 was also decreased significantly by VO(p-dmada) treatment (Fig.S5B in Supporting information).These results indicated that the inhibitory effects of VO(p-dmada)on LPS-induced TNF-α, IL-6 and IL-12 secretion came from the suppression of NF-κB signaling pathway.
We further investigated the effects of VO(p-dmada) on LPSinduced inflammatory responses in mice and the experimental procedures for mice treatment are illustrated in Fig.S7A (Supporting information).Whether the novel compound can pass through the blood-brain barrier (BBB) is a checkpoint for developing drug to treat brain diseases.In a previous study, we confirmed that BEOV was able to cross the BBB in APPswe/PS1E9 transgenic mice [35].Similarly, we verified in the current study that VO(pdmada) could also pass through the BBB and reach into the brain parenchyma (Figs.S7B and C in Supporting information).Ionized calcium binding adaptor molecule-1 (Iba-1) and glial fibrillary acidic protein (GFAP) are specifically expressed in the microglia and astroglia, respectively, and up-regulated upon stimulation of toxicants [36].The expression levels of Iba-1 and GFAP were measured in the hippocampus by Western blot and immunofluorescence staining.We found that the protein levels of Iba-1 and GFAP were significantly increased in the LPS-stimulated group in hippocampal tissue when compared with those in control group.However, treatment with VO(p-dmada) significantly suppressed the levels of Iba-1 and GFAP (Figs.1A-C and Fig.S8 in Supporting information).Furthermore, VO(p-dmada) significantly reduced the production of pro-inflammatory enzymes and cytokines including iNOS, COX-2, TNF-αand IL-6 in the brain and serum of LPS-treated mice (Figs.1D-G).These findings support the speculation that VO(p-dmada) can pass through the BBB to exert antineuroinflammatory effects in brain.

Fig.1.VO(p-dmada) suppressed the activation of microglia and astrocytes, the release of pro-inflammatory factors, and the expression of iNOS and COX-2 in LPS-stimulated mice.(A, B) The protein expression levels of GFAP and Iba1 in the hippocampus tissues detected by Western-blot analysis.(C) Representative immunofluorescent images of astrocytes (GFAP, green) in CA1, CA3 and DG region of the hippocampus.(D, E) The protein expression levels of iNOS and COX-2 detected by Western-blot analysis.(F, G)Levels of TNF-α and IL-6 in the serum and hippocampus measured by ELISA kit.#: LPS group vs.untreated group; *: LPS group vs.LPS+VO(p-dmada) group.#P <0.05, ##P<0.01, ###P <0.001, *P <0.05 and **P <0.01, respectively.Scale bar: 100 μm.
To systematically investigate the proteins affected by VO(pdmada) treatment, a quantitative proteomic analysis based on SWATH methodology was used to identify differentially expressed proteins (DEPs) in hippocampus.SWATH-MS analysis identified a total of 4006 quantitative proteins with 2825 quantitative proteins meeting the quality control requirements after QC correction.Among them, 76, 201, 44 and 31 proteins were identified as DEPs in LPS-inducedvs.control groups (LPS/Ctrl), LPS+0.1 mmol/kg VO(p-dmada) treatedvs.LPS groups (Low/LPS), LPS+0.2 mmol/kg VO(p-dmada) treatedvs.LPS groups (High/LPS), and High/Low groups, respectively (Tables S2-S5 in Supporting information, Figs.2A-D).Among the DEPs in LPS/Ctrl group, 66 proteins were upregulated, and 10 proteins were down-regulated (Fig.2A).Gene ontology (GO) analysis and KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway enrichment were then performed to those DEPs (Fig.2 and Fig.S9 in Supporting information).GO analysis showed that the biological processes (BPs) linked to DEPs of LPS/Ctrl groups were mainly enriched in the response and regulation to stress, vesicle-mediated transports, cell-cell signaling, secretion, neuron projection development and immune system process,etc.(Fig.2E).KEGG pathway analysis showed that the majorities of DEPs were involved in endocytosis, metabolism, dopaminergic synapse and adrenergic signaling in cardiomyocytes,etc.(Fig.2F).Interestingly, the results of the cluster analysis showed that the protein expression pattern of the LPS group remarkably different from those of the other three groups, while the two VO(p-dmada)-treated groups and the Ctrl group had similar pattern.Interestingly,the majority of DEPs up-regulated in LPS/Ctrl groups were downregulated in two LPS+VO(p-dmada)/LPS groups (Fig.2G).Among the DEPs in Low/LPS groups, 4 proteins were up-regulated and 197 proteins were down-regulated (Table S3 and Fig.2B).Among the DEPs in High/LPS groups, 8 proteins were up-regulated and 36 proteins were down-regulated (Table S4 and Fig.2C).GO analysis of BP, cellular component (CC) and molecular function (MF)and KEGG pathways enrichment of these proteins were shown in Figs.S10A-D and S11 (Supporting information).In addition, the GO-BP enrichment results of DEPs in High/LPS groups contained a part of the enriched terms for DEPs in Low/LPS groups (Fig.S10E in Supporting information).The Pathway Activation Strength(PAS) value was calculated for DEPs byomicsbeanonline tool.In the LPS/Ctrl groups, DEPs-activated processes included the responses to stress, oxygen-containing compound and immune system.The changes of these processes were reversed in a certain degree after VO(p-dmada) treatment (Fig.S12 in Supporting information).The protein-protein interactions (PPI) network further confirmed that the majority of DEPs in LPS/Ctrl, Low/LPS and High/LPS groups were enriched in the responses to stress, oxygen-containing compound and inflammation (Fig.S13 in Supporting information).Among them, transglutaminase 2 (Tgm2), heat shock protein 60(Hspd1), Parkinson disease protein 7 homolog (Park7), 14-3-3 protein zeta/delta (Ywhaz), and peroxiredoxin 2 (Prdx2) were proteins closely associated with inflammatory response, which were identified as the key proteins of VO(p-dmada) treatment in LPS-induced mice (Fig.S13D).Those proteomic results further verified the beneficial effects of VO(p-dmada) on modulating neuroinflammatory reactions.
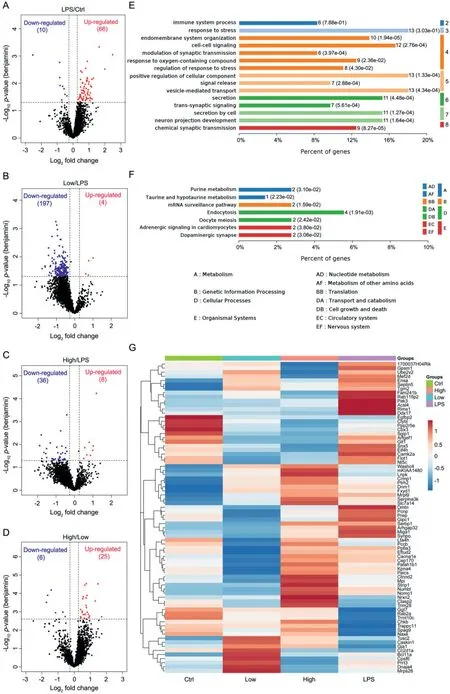
Fig.2.Proteomic identification and bioinformatic analysis of differentially expressed proteins (DEPs) in different groups.(A-D) Volcano plots depicted the distribution of DEPs in the LPS-treated group vs.control group (LPS/Ctrl), LPS+0.1 mmol/kg VO(p-dmada) treated group vs.LPS group (Low/LPS), LPS+0.2 mmol/kg VO(p-dmada) treated group vs.LPS group (High/LPS), and High/Low groups.(E) GO analysis results of the top 15 biological processes (BP) enriched from the DEPs of LPS/Ctrl.(F) KEGG pathways related to the DEPs of LPS/Ctrl.(G) Cluster map comparing the DEPs identified in LPS/Ctrl, Low/LPS, High/LPS, and High/Low groups.

Fig.3.Docking configurations of VO(p-dmada) with PPARγ.(A) The docking mode of VO(p-dmada) with PPARγ protein.(B, C) VO(p-dmada) interacts with the ligand-binding domain of PPARγ and forms three hydrogen bonds at the residues Ser 342, Gly284 and Arg288, shown by dashed yellow and green line.

Fig.4.Down-regulation of iNOS and COX-2 expression and inactivation of NF-κB signaling by VO(p-dmada) was impeded under the co-treatment of PPARγ inhibitor GW9662.(A, B) LPS-induced BV2 microglia were treated with VO(p-dmada) (10 μmol/L) or both 10 μmol/L VO(p-dmada) and 20 μmol/L GW9662 for 24 h.The protein expression levels of iNOS and COX-2 were detected by Western-blot analysis.(C, D) The levels of p-IκB-α, IκB-α, p-p65 and p65 were measured by Western-blot analysis.n=6 wells per group, # : LPS group vs.untreated group or LPS+VO(p-dmada) group; *: LPS+VO(p-dmada)+GW9662 group vs.LPS+VO(p-dmada) group. #P <0.05, ##P <0.01, ###P <0.001, *P <0.05 and **P <0.01, respectively.
Transglutaminase 2 (TG2 or TGase 2, gene name: Tgm2) and heat shock protein 60 (Hsp60, gene name: Hspd1) are two proteins that play essential roles in regulating inflammatory response[37,38].LPS could induce up-regulation of TG2 and Hsp60, leading to inflammatory cascade reactions [39,40].To verify the results of proteomic study, Western blot analysis was performed for those two proteins.As shown in Figs.S14A and B (Supporting information), the levels of TG2 and Hsp60 were remarkably increased in the hippocampus of LPS-induced mice compared with the vehicletreated control.Treatment with VO(p-dmada) significantly reduced TG2 and Hsp60 protein levels.
PPARγ, an important ligand-activated transcription factor, plays a critical role in regulating gene expression during several physiologic and pathological processes [41,42].TG2 could control inflammation through PPARγin cystic fibrosis epithelial cells [43].Previous study reported that overexpression of Hsp60 significantly decreased PPARγprotein level and vanadium compounds were able to elevate PPARγprotein level in mouse insulinoma NIT-1 cells through regulating Hsp60 [20].Here, we examined the effect of VO(p-dmada) on PPARγexpression in LPS-stimulated BV2 cells and BALB/c mice.As shown in Figs.S14C-H (Supporting information),LPS treatment significantly reduced PPARγexpression both in BV2 microglia and mice, while VO(p-dmada) effectively upregulated the protein level of PPARγ.These findings suggest that VO(p-dmada) is possible to indirectly increase the level of PPARγthrough downregulation of TG2 and Hsp60 expression.
Then, is it possible for VO(p-dmada) to act directly on PPARγ?To answer this question, we applied the softwares Autodock vina 1.1.2 and Free of Meastro 10.1 to mimic the binding of VO(pdmada) and PPARγmolecules.The binding energy of VO(p-dmada)with PPARγwas calculated to be -8.2 kcal/mol, indicating a superior binding affinity between VO(p-dmada) (as a ligand) and PPARγ(as a receptor).As shown in Fig.3, three hydrogen bonds formed between VO(p-dmada) and PPARγat residues Arg288, Gly284 and Ser342, demonstrating that VO(p-dmada) can interact directly with PPARγat high binding affinity and thus functions as a PPARγagonist to regulate its activity.
Cumulative evidences have shown that activated PPARγcould inhibit NF-κB signaling pathway and in turn mitigated neuroinflammation bothin vivoandin vitro[7,44].Bisperoxovanadium and BEOV have been reported to increase PPARγactivity and inhibit inflammation [7,21].In order to investigate the mechanism of VO(p-dmada) in regulating LPS-evoked neuroinflammation, BV2 microglia were treated with LPS, VO(p-dmada) and GW9662 (a general PPARγinhibitor), simultaneously.As expected, the results showed that GW9662 remarkably impeded the effect of VO(pdmada) on increasing the protein levels of PPARγ(Fig.S15 in Supporting information).The levels of released inflammatory mediators including TNFα, IL-6, IL-12, PGE2and NO were also meausured using ELISA kit and Griess reagent.As shown in Fig.S16 (Supporting information), the inhibitory effect of VO(p-dmada) on inflammatory mediators was eliminated after adding GW9662.Moreover,the inhibition of PPARγby GW9662 markedly restored the levels of iNOS and COX-2 that were reduced by VO(p-dmada) treatment(Figs.4A and B, Fig.S17 in Supporting information).Further study on VO(p-dmada) in regulating NF-κB signaling pathway, the levels of IκB-αand p65 phosphorylation were detected.As shown in Figs.4C and D, GW9662 also inhibited the down-regulating effect of VO(p-dmada) on these proteins.Taken together, these findings indicated that VO(p-dmada) treatment attenuated the LPS-stimulated neuroinflammation by inactivating NF-κB signalingviaa PPARγdependent mechanism.
In summary, we demonstrated, for the first time, that VO(pdmada) can attenuate proinflammatory mediator production and NF-κB signaling pathway activation in LPS-stimulated mice and BV2 microglia.The underlying mechanism of VO(p-dmada) in inhibiting inflammatory response is concluded and shown in Fig.S18(Supporting information).VO(p-dmada) thus might be a promising candidate for treating neuroinflammation-based brain diseases.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This study was financially supported by grants from the National Natural Science Foundation of China (No.21877081), the China Postdoctoral Science Foundation (No.2021M692210), Guangdong Provincial Key S&T Program (No.2018B030336001), the Shenzhen Science and Technology Innovation Commission (No.JCYJ20200109110001818), and the Shenzhen-Hong Kong Institute of brain Science-Shenzhen Fundamental Research institutions (No.2022SHIBS0003).We would like to thank the Instrumental Analysis Center of Shenzhen University and the Central Research Facilities of College of Life Sciences and Oceanography of Shenzhen University for the use of laser scanning confocal microscope.
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2023.108236.
 Chinese Chemical Letters2023年10期
Chinese Chemical Letters2023年10期
- Chinese Chemical Letters的其它文章
- Tribute text in memoriam of James N.Seiber (1940–2023)
- Recent advances in MXenes-based glucose biosensors
- Oxidative cyclopalladation triggers the hydroalkylation of alkynes✩
- An integrated supramolecular fungicide nanoplatform based on pH-sensitive metal–organic frameworks
- Probing the effect of nitrate anion in CAN: An additional opportunity to reduce the catalyst loading for aerobic oxidations✩
- Nickel-catalyzed reductive coupling reaction of monofluoroalkyl triflates with alkyl carboxylic acids toward the synthesis of α-alkyl-α-fluoro-alkylketones✩