
思茅松半同胞和全同胞子代测定林生长性状遗传分析与选择
2023-12-25 12:41孙晓梅董雷鸣
中南林业科技大学学报 2023年11期
陈 伟,孙晓梅,董雷鸣,李 江,付 强
(1. 中国林业科学研究院林业研究所 林木遗传育种国家重点实验室,北京 100091;2. 云南省林业和草原科学院,云南 昆明 650201;3. 国家植物园,北京 100093;4. 普洱市林业和草原种苗工作站,云南 普洱 665000)
遗传测定是林木育种的核心。基于表型性状选出的优树或是通过交配设计获得的子代,其遗传品质的优劣程度需要通过合理的试验设计,经田间对比试验和遗传分析后才能进行评定[1]。遗传测定对于遗传参数的准确估计、遗传增益的预测、选择年龄的确定和改良策略的制定都具有重要的指导意义[1-2]。子代测定作为遗传测定的主体部分,估算亲本无性系和子代单株的育种值,用于后向选择,可作为亲本无性系留优去劣的依据,用于前向选择,在全同胞优良家系中优选个体为高世代遗传改良提供育种材料[3]。基于前期良好的遗传测定,我国许多重要造林树种已完成了第1 代改良进入高世代育种阶段,如杉木Cunninghamialanceolata已进入第三代遗传改良阶段[4-5],马尾松Pinusmassoniana[6-7]、日本落叶松Larixkaempferi[8]、油松Pinustabuliformis[9]等全面进入第二代改良,并开始创制第3 代种质[10]。
思茅松Pinuskesiyavar.langbianensis是我国特有的针叶树种,主要分布于云南南部哀牢山以西的南亚热带山地,具有速生、适应性强、材质优良和松脂产量高等特点,为云南林产工业的重要树种。围绕着产业需求,前期研究主要聚焦于育苗技术[11]、空间配置模式[12]、病虫害防治[13]和施肥[14]等人工林培育技术的研究,同时在群落结构[15]、生物量模型[16]、碳汇计量[17]和土壤有机碳[18]等生态学研究方面也开展了大量的工作,但遗传改良起步较晚,在前期优树选择与资源收集工作的基础上[19-20],20 世纪90 年代建成了第一个思茅松无性系种子园,基于初级种子园开展了花期物候观测[21]、花粉萌发与保存技术[22]、控制授粉[23]和子代测定[24]等基础工作的探索,与其他主要针叶造林树种相比差距较大,仍停滞于一代改良阶段[25],不能有效支撑思茅松下游产业的快速发展。与此同时,在思茅松一代改良材料广泛应用的基础上,积累了大量的表型数据。如何利用现有数据进行科学分析,对前期遗传改良的成效进行评估,从而更好地制定出针对思茅松树种特性和现有基础的长期育种策略,是当前思茅松遗传改良工作中急需解决的科学问题。基于此,本研究对2000 年定植的思茅松半同胞子代测定林和2013 年定植的全同胞子代测定林数据进行分析,科学评估参试材料及其亲本的遗传特性,以便更好地服务于思茅松高世代遗传改良。
1 材料与方法
1.1 试验林概况
子测林1(半同胞子代测定林):位于云南省普洱市景谷县威远镇文郎村,地理坐标为(23°29′50″N,100°29′15″E),海拔1 550 m,属南亚热带季风气候区,造林前植被类型为思茅松为主的针阔混交林。参试家系29 个,来源于普文思茅松无性系种子园,以当地商品种子为对照,2000年3 月育苗,7 月造林,采用随机完全区组设计,4 次重复,9 株方形小区,株行距3 m×3 m,试验区的四周各设置1 行保护带,定植穴规格40 cm×40 cm×40 cm,每株施150 g 复合肥作底肥,于定植当年9 月和11 月各除草1 次,以后每年除草2 次,连续抚育3 年。
子测林2(全同胞子代测定林):位于云南省普洱市思茅区思茅镇莲花村,地理坐标为(22°54′11″N,100°54′53″E),海拔1 430 m,南亚热带季风气候区,造林前植被类型为思茅松人工林。共12 个全同胞家系,为2×6 的测交系子代,父本和母本均源于普文思茅松无性系种子园,2014年3 月育苗,7 月造林,采用随机完全区组设计,5 次重复,4 株方形小区,株行距2 m×3 m,试验区的四周各设置一行保护带,定植穴规格40 cm×40 cm×40 cm,抚育方式同上。
1.2 数据调查与分析方法
1.2.1 表型数据测定
子测林1 在林龄14 a 时、子测林2 在林龄6 a时进行生长性状调查。树高采用测高仪测量,胸径采用胸径尺测定,单株材积按原林业部颁布的标准公式[26]计算。
1.2.2 遗传参数估计
采用线性随机效应模型和限制性最大似然估计方法估算随机效应中的方差分量[27],半同胞子测林为模型(1),全同胞子测林为模型(2)。
式中,y为观测值向量,μ为观测性状的均值,b为固定的区组效应,f为随机的家系效应、fb为随机的家系区组互作效应向量,a为随机的加性遗传效应,d为随机的显性效应,e为随机残差,X、Z1、Z2、Z3、Z4分别为对应效应的关联矩阵。
利用似然比检验各方差组分的统计显著性,利用泰勒级数展开法计算遗传参数的标准误[27]。
遗传变异系数(CVG)和表型变异系数(CVp)分别按公式(3)~(4)计算:
式中,σg2σ2p、分别为遗传方差、表型方差、总体均值,其数值依据公式(1)~(2)由计算机通过R 软件求出。
半同胞子测林单株遗传力(hi2)和家系遗传力(h2f)分别按公式(5)~(6)计算:
式中,σ2f、σ2fb、σe2分别为家系方差,家系与区组互作效应方差和残差方差,其数值依据公式(1)(2)由计算机通过R 软件求出,n为小区株数,b为区组数。
全同胞子测林狭义单株遗传力(hi2)、广义单株遗传力(Hi2)和广义家系遗传力(H2f)参照文献[28]分别按公式(7)~(9)计算:
式中,σd2=4σs2,σa2、σd2、σs2、σb2、σe2分别为加性方差、显性方差、特殊配合力方差、区组方差和残差方差,其数值依据公式(1)~(2)由计算机通过R 软件求出,n为小区株数,b为区组数。
表型相关系数(rp),遗传相关系(rg)数按公式(10)~(11)计算:
σg(xy)、σp(xy)分别为性状x和性状y的遗传和表型协方差,分别为性状x和y的遗传方差和表型方差,其数值依据公式(1)~(2)由计算机R 软件求出。
采用BLUP 法计算不同组合一般配合力和特殊配合力并估算育种值,基于育种值和遗传值(育种值+特殊配合力)计算目标性状的遗传增益[29-30]。
1.2.3 优质遗传材料的选择
以材积性状开展后向选择,基于育种值在控制家系入选率和遗传增益的条件下开展优良家系的选择;通过混合方程组法(MME)求解获得参试半同胞家系子代个体的育种值,参照家系和单株配合选择二代优树的方法[31]前向选择二代优树。
1.2.4 数据处理
除子测林1 的19 号家系保存率仅30.6%未参与计算外,其余家系保存率均大于60%。采用Excel 2010 软件进行数据的录入和转换,方差分析、遗传参数和育种值的估算采用R 软件和ASReml-R包进行分析[32]。
2 结果与分析
2.1 生长性状的变异性分析
对子测林参试家系的生长性状进行统计(表1)。14 年生半同胞家系树高、胸径和材积的平均值分别为15.66 m、23.12 cm 和0.349 3 m3,材积的表型变异系数最大,达42.9%,胸径次之,树高相对较小,也超过10%。各性状遗传变异系数为8.95%~24.00%,材积的遗传变异最大,是胸径遗传变异系数的2.61 倍、树高遗传变异系数的2.68倍,具有较大的选择潜力。6 年生全同胞家系树高、胸径和材积均具有丰富的表型变异,变幅分别为3.20 ~7.00 m、3.50 ~12.60 cm 和0.001 8 ~0.047 3 m3,但遗传变异相对较小,材积遗传变异系数最大也仅达3.45%。
2.2 生长性状的遗传参数估算
利用限制性最大似然估计方法对2 个子测林生长性状的方差分量进行估算(表2 ~3)。14年生半同胞子代测定林树高、胸径和材积的家系遗传力分别为0.680、0.691 和0.748,受较强的遗传控制;树高性状的单株遗传力为0.656,属于高遗传力,其家系方差组分占表型方差组分的比例相对较高,胸径和材积性状的单株遗传力分别为0.234 和0.321,属于中等遗传力,胸径性状中家系×区组的方差组分占表型方差组分的比例相对较小,说明家系和区组互作效应对胸径生长性状的差异影响较小。
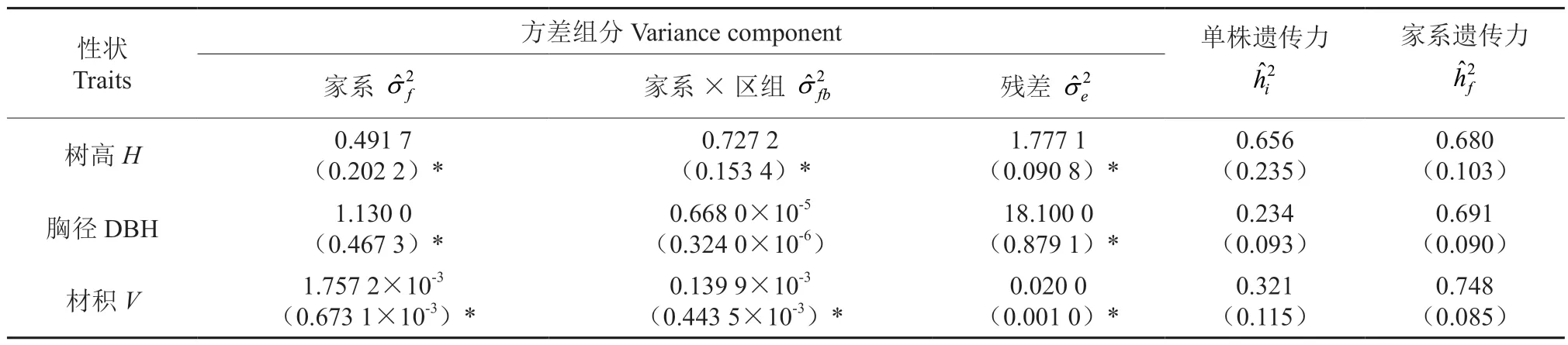
表2 半同胞家系生长性状遗传参数估算†Table 2 Estimation of genetic parameters of growth traits in half-sib families
6 年生全同胞子代测定林生长性状的方差组分见表3。3 个性状的狭义单株遗传力均小于0.01,表明其受加性效应的控制较弱。树高、胸径和材积3 个性状的广义单株遗传力分别为0.233、0.247和0.191,广义家系遗传力分别为0.245、0.256 和0.204,表明3 个性状的表型变异不仅受环境影响,还受中等程度的显性效应影响,不同交配组合的特殊配合力在表型变异方面具有一定程度的贡献。
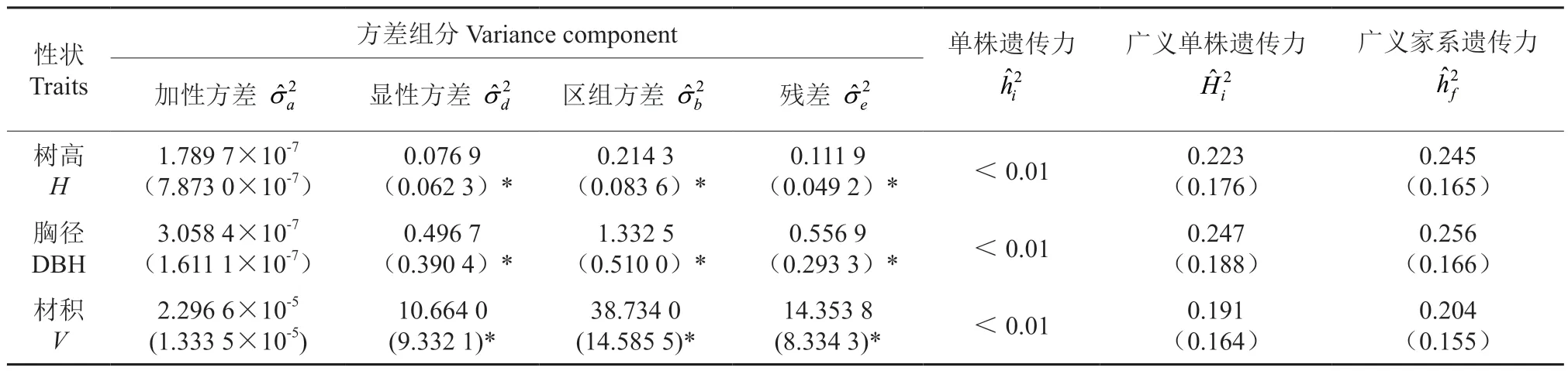
表3 全同胞家系生长性状遗传参数估算Table 3 Estimation of genetic parameters of growth traits in full-sib families
基于生长性状的加性遗传方差、两两性状的协方差和误差方差计算出了树高、胸径和材积性状间的遗传相关和表型相关关系(表4)。无论是半同胞子测林还是全同胞子测林,3 个性状间均存在极显著的遗传和表型的正相关关系(P<0.01),以材积与胸径间遗传相关系数最大,表型相关系数均低于对应的遗传相关系数。
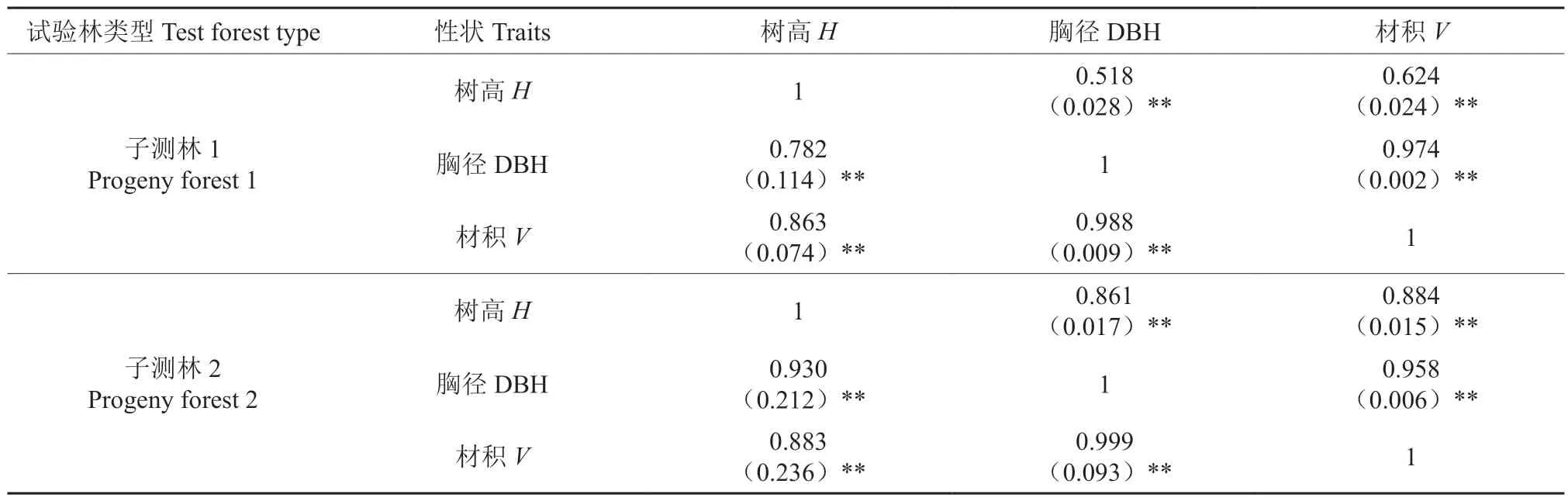
表4 不同生长性状相关分析†Table 4 Correlation analysis of different growth traits
2.3 优良遗传材料的选择
14 年生半同胞子测林各参试家系材积性状均值、变异系数和育种值估算见表5,参试半同胞家系子代个体的育种值见表6。14 年生家系间单株材积均值的变幅为0.192 4 ~0.422 0 m3,变异系数在26.78%~55.19%,家系间和家系内部均具有较大的变异,其中有25 个家系的材积均值超过对照(0.285 2 m3)。

表5 半同胞家系材积变异与育种值估计Table 5 Volume variation and breeding value estimation of half-sib families
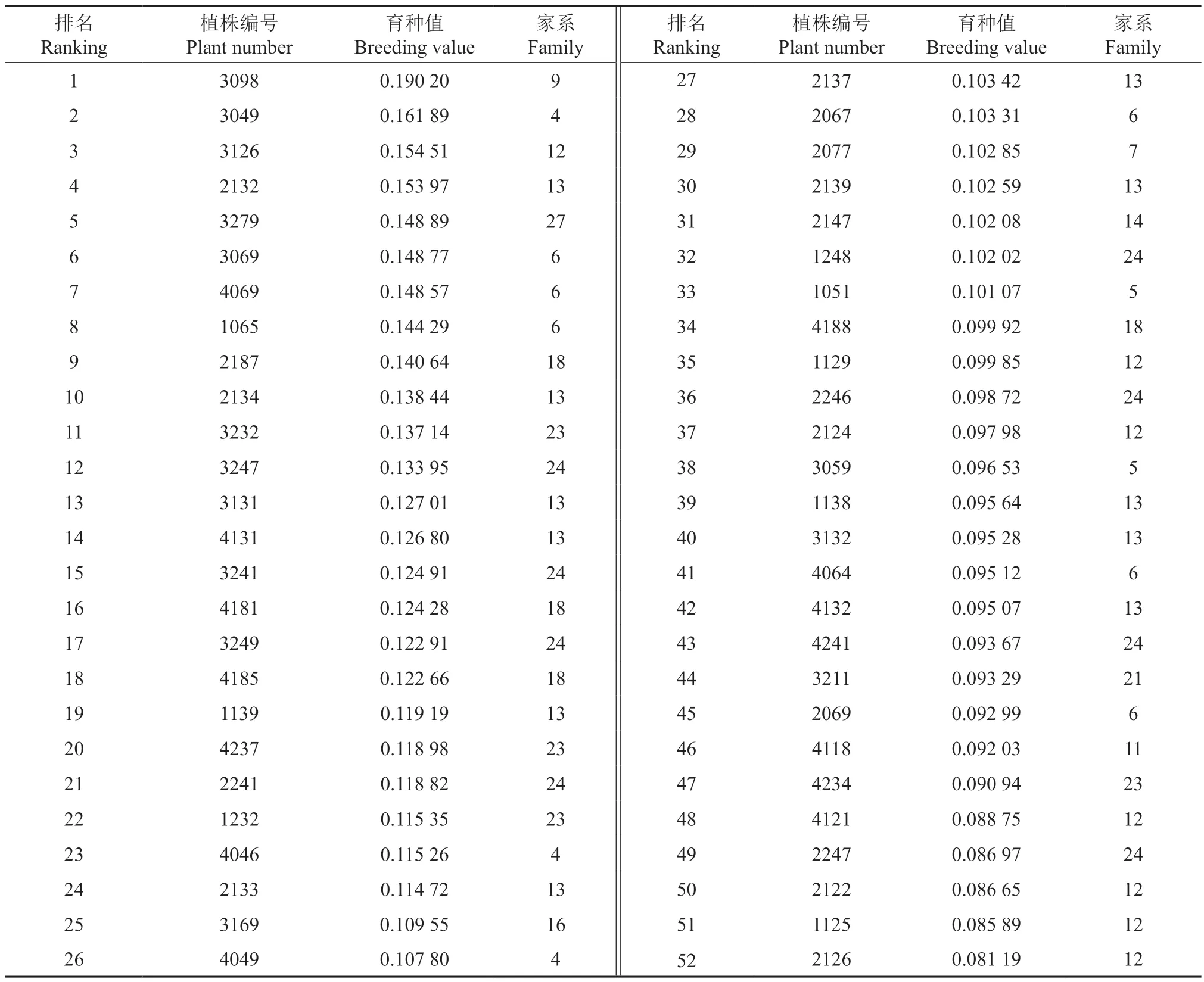
表6 部分半同胞家系子代个体材积育种值相对排名Table 6 Relative ranking of individual volume breeding value of partial half-sib families
在半同胞子测林中以材积性状开展优良家系的后向选择,以材积育种值为指标,对参试家系的育种值进行排序,按入选家系总遗传增益不低于10%且入选率不高于20%的原则,选出6 个优良家系,分别为12、13、18、24、6 和4 号,6 个入选家系的材积较对照提高37.9%~48.0%,材积均值为0.403 7 m3,材积遗传增益达11.47%。根据参试半同胞家系子代个体材积育种值相对排名进行优良单株的前向选择,按参试子代个体数量的5%的入选率,共选出植株编号为3098、3049 等52 株优良单株,入选的优良单株分属于15 个家系,其中6 个入选优良家系子代的比例为78%,入选优良单株材积均值为0.684 1 m3,材积遗传增益达30.64%。鉴于全同胞子测林3 个生长性状具有丰富的表型变异、加性遗传效应低而显性遗传效应中等的现状,以材积为选择目标,以遗传值为选择指标,开展最优交配组合的选择。按20%的比例进行最优交配组合的选择,选出P016-P019、P020-P008 等2 个最优组合,2 个组合材积均值为0.023 2 m3,其材积遗传增益为15.53%(表7)。
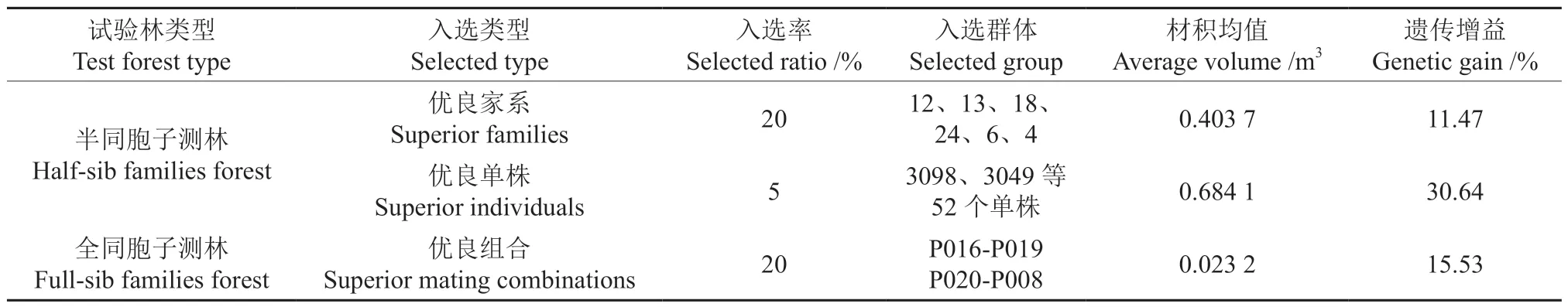
表7 子测林不同选择的遗传增益Table 7 Genetic gain of different selection in progeny test forest
3 讨论与结论
3.1 讨 论
遗传力反映了遗传变量占表型变量的比率[1],表征性状受遗传效应控制的强弱[33]。不同物种不同性状受遗传控制的强度不一致,如日本落叶松树高主要受显性效应控制,胸径和材积主要受加性效应控制,材性的化学性状除了受加性效应控制外,也受显性效应控制[34]。遗传力因特定群体、特定时间、特定环境而变化[1],也与性状的变异特性、试验设计、计算方法等有关系[33]。本研究中,14 年生思茅松半同胞子测林的树高、胸径和材积家系遗传力受高等强度的遗传控制,单株遗传力也属于中等以上遗传控制。因试验地点、试验设计、测定年龄以及计算方法的不同,本次估算的遗传力与赵文书等[24]估算的遗传力存在差异。6 年生全同胞子代测定林3 个生长性状的狭义遗传力较低,可能与分析数据为单地点单次测定值有关,其遗传力是否也会随着林龄的增长而变化,有待进行连续观测。显性效应只有当特定基因组合在一起时才能表现出来[1],在火炬松和辐射松的遗传改良过程中就发现某些生长性状受显性效应(特殊配合力)控制较为突出[35-36]。本研究中,6年生全同胞家系子代测定林狭义单株遗传力较低,但中等强度的广义遗传力反映了参试材料受较大程度的显性效应影响,说明特殊配合力在全同胞测定林生长性状的变异中发挥着重要作用。特殊配合力一般通过两种方式进行利用,一是通过无性繁殖利用,二是通过建立特殊组合的杂交种子园进行利用[1]。通过遗传值选出的2 个优良组合可通过成熟的思茅松扦插繁殖技术充分利用特殊配合力效应,最大化地获得遗传增益服务于生产需求。
半同胞家系是自由授粉产生的子代,因其具有易得、成本低、可估算遗传参数的特点,其测定林在林木遗传改良早期阶段发挥着重要的作用,但由于父本未知,能估算的遗传参数有限。全同胞家系是完全谱系设计的子代,因其父本和母本清楚,在遗传分析时能得到更多的遗传信息,不仅能估算特殊配合力、父本一般配合力等半同胞子测林无法估算的遗传参数,而且预估的精度也更高。本研究中,思茅松半同胞子代测定林估算出的遗传力较全同胞子代测定林高,除测定年龄、地点等因素外,全同胞家系中区组方差组分在方差组分中的比例相对较大,而半同胞家系受计算模型的限制,其区组方差组分被忽略,计算遗传力时加性效应中可能包含了非加性效应等,导致估算的遗传力偏高。因此,对于半同胞子代测定林可以通过系谱重建的方式,获得完整的系谱信息,以获得更为丰富的遗传信息、更为准确的遗传参数和育种值,更有助于提高半同胞子代测定效率[34]。
基于育种值的选择以基因型代替表型值进行性状的选择,能够极大地提高选择的效率和精度[37]。林木遗传育种有别于作物育种,周期长,测定林参试材料的保存率差异大,存在试验数据的不平衡现象,适宜的遗传评价和选择方法显得尤为重要。育种值估算的方法有很多,在林木育种的实践中,对于有亲缘关系或具有不同遗传固定效应的候选材料,BLUP 法更为适用,尤其在群体规模大,群体结构复杂,观测数据不平衡的情况下,仍可获得较为准确的估计育种值,并在林木遗传改良中得到广泛应用[37]。半同胞子代测定林基于亲本育种值的后向选择,可以指导种子园去劣疏伐和1.5 代种子园建园材料的选择[3],也可以为创制二代思茅松育种材料的选择亲本和交配设计提供依据。基于个体育种值的前向选择可提高遗传改良的选择效率[37-38]。在半同胞测定林中,研究基于个体的育种值进行优良单株选择显示出了较高的选择效率,本研究采用家系和单株配合选择方法选出的优良单株78%的比例来源于选出的优良家系,22%的比例来源于其他家系,在保证优良家系优良单株入选的情况下,又最大限度地选择了子代育种值相对较好的非入选优良家系的优良单株,兼顾遗传增益最大化及入选群体的遗传多样性水平。此外,本研究未对入选的优良家系的优良单株进行株数限制,基于两方面的原因,一方面配合选择时,性状遗传力高时,应给予个体较高的权重[1],可以适当扩大选择的数量;另一方面,思茅松遗传改良基础薄弱,基础的遗传材料相对较少,适当扩大优良家系优良单株入选数量,可以增加思茅松育种材料的选择面,符合思茅松遗传改良的现状,而且随着分子生物学技术的发展,可以应用系谱重建技术控制新入选子代间的亲缘关系,为科学的育种策略的制定提供理论依据[34]。
在研究结果的应用上,半同胞子代测定林中选出的优良单株可入选思茅松二代育种群体,用作下一轮遗传改良的材料,但在思茅松系谱重建技术成熟以前,鉴于半同胞子代谱系不清,按照传统的遗传改良策略,思茅松二代育种群体中仍然应以谱系清楚的全同胞子代中的优良单株为主,半同胞子代优良单株的数量应有所控制。本研究虽然开展了全同胞子代测定,但受限于交配设计简单,杂交组合数量少,且测定结果生长性状的非加性效应较大等因素的影响,在全同胞子测林中未能选出适宜进入下一轮遗传改良的材料,其应用上有一定的局限性。下一步将加大全同胞子代种质的创制工作,通过构建思茅松一代核心育种群体,基于科学的交配设计在核心育种群体间开展一定规模的控制授粉工作,创制出一大批全同胞子代,经过子代测定前向筛选出符合要求的遗传材料,更好地服务于思茅松高世代遗传改良工作。同时,为了高效利用杂交组合的特殊配合力,可以在思茅松优质种质资源的无性系化利用方向开展新的攻关,可借鉴相近物种马尾松成熟胚高质量芽诱导的组培快繁植株再生体系的方法[39]开展思茅松组培技术研究,高效利用优质的思茅松种质资源。
3.2 结 论
思茅松子测林材积性状的表型和遗传变异系数相对较高,适宜作为遗传选择的主要生长性状,14 年生思茅松半同胞家系子代测定林,树高、胸径和材积3 个生长性状受中等以上的遗传控制,基于材积育种值,后向选择出的优良家系和前向选择出的优良单株均具有较好的遗传增益。6 年生思茅松全同胞子代测定林,树高、胸径和材积3个生长性状受中等程度的显性效应影响,可利用遗传值选出优良交配组合,通过无性系化利用可获得最大化的遗传增益。基于子测林的遗传分析,可为思茅松的持续遗传改良选出较好的遗传材料。
猜你喜欢
养猪(2022年4期)2022-08-17
东北林业大学学报(2021年1期)2021-12-03
东北林业大学学报(2021年1期)2021-01-14
山东林业科技(2017年1期)2017-06-29
西南农业学报(2016年6期)2016-04-16
上海农业学报(2016年5期)2016-02-10
湖北畜牧兽医(2015年11期)2016-01-11
武夷学院学报(2015年3期)2015-07-18
山东农业科学(2014年1期)2015-03-09
长春工程学院学报(自然科学版)(2013年4期)2013-12-06
