
秸秆焚烧对白叶枯病稻田中致病菌和抗生素抗性基因的影响
2023-12-29 08:20王彬浩陈彦博刘荣杰王冬群董作珍狄蕊王笑吴愉萍
浙江大学学报(农业与生命科学版) 2023年6期
王彬浩,陈彦博,刘荣杰,王冬群,董作珍,狄蕊,王笑,吴愉萍
(1.浙江大学环境与资源学院,浙江 杭州 310058;2.浙江大学杭州国际科创中心,浙江 杭州 311200;3.宁海县农业技术推广站,浙江 宁波315600;4.慈溪市农业监测中心,浙江 宁波 315300;5.宁波市海曙区农业技术管理服务站,浙江 宁波 315012;6.宁波市镇海区农业技术推广总站,浙江 宁波 315200;7.宁波市江北区农业技术推广服务站,浙江 宁波 315033;8.宁波市农业农村绿色发展中心,浙江 宁波315012)
作物秸秆是在农业生产过程中产生的一种生物质资源。作为农业大国,我国2021年农作物秸秆总量为8.65亿t,其中秸秆直接还田量为4.02亿t,有力推动了秸秆的绿色利用[1]。然而,由于在实际生产过程中,秸秆直接还田在还田量、还田方式、还田配套农艺措施等方面仍存在许多问题,对后茬作物的产量和病害暴发存在负面影响[2],因此,部分农户仍然采用焚烧方式处理秸秆,以期避免后茬作物发病。
水稻白叶枯病是由水稻黄单胞菌(Xanthomonas oryzae)侵染引起的一种细菌性病害,是水稻三大病害之一[3]。水稻感染白叶枯病后,通常减产10%~30%,严重时减产50%以上[4]。此外,由于稻田通常是常年连作,这种方式不仅会加剧致病菌富集,而且有利于抗生素抗性基因(antibiotic resistance genes,ARGs)的富集,并通过移动基因元件(mobile genetic elements, MGEs)加速病菌的水平传播,最终导致耐药性致病菌(生物复合污染)的比例不断升高。例如,随着单一作物的连续种植,土传植物致病菌会不断积累,从而导致枯萎病、青枯病等土传病害的暴发[5]。棉花、黄瓜、马铃薯及香草等作物的农业系统均易受连作效应的影响,随连作时间的延长,其土壤中土传植物致病菌丰度增高[6-8]。已有的研究表明,秸秆焚烧会造成土壤微生物数量和活性的显著下降[9-10],这对于揭示田间秸秆焚烧抑制白叶枯病的潜在机制提供了一定的依据。然而,目前关于秸秆焚烧的影响研究多集中在大气和土壤理化环境变化方面,针对秸秆焚烧前后稻田土壤中致病菌或ARGs的变化情况少见报道。本试验以暴发白叶枯病的稻田土壤和稻茬为研究对象,对比焚烧前后土壤理化特性、微生物致病菌、ARGs、MGEs的变化情况,探讨稻田致病菌群落和ARGs 组成与土壤含水量、有机质含量、速效养分含量的相关性,以期为综合解析和客观评价秸秆焚烧对土壤生物复合污染的影响提供一定的参考。
1 材料与方法
1.1 样品采集
取样调查地点位于浙江省宁波市,分别为慈溪市龙山镇海甸戎村、海曙区洞桥镇宣裴村和集士港镇朱浪漕村、宁海县茶院乡下徐塘村、镇海区蟹浦镇觉渡村和骆驼街道清湖村,包括了该市主要的水稻种植区域(表1)。于2021 年11—12 月在暴发白叶枯病的稻田中对秸秆焚烧前后的土壤和稻茬分别取样,各采集16 个混合样品。土壤样品:除去土壤表面的灰分和秸秆,用内径为5 cm的土钻在各小区取0~5 cm表层的土样,去除石砾和根系,每个样品均由3 点土样混合而成(约0.5 kg),并分成两份,一份保存在-20 ℃冰箱中,用于微生物的测定,另一份于室内风干,用于土壤含水量、有机质含量、速效养分含量等理化因子的测定。稻茬样品:收集留茬秸秆样品(取3茬,每茬取2根),保存在-20 ℃冰箱中,用于微生物的测定。
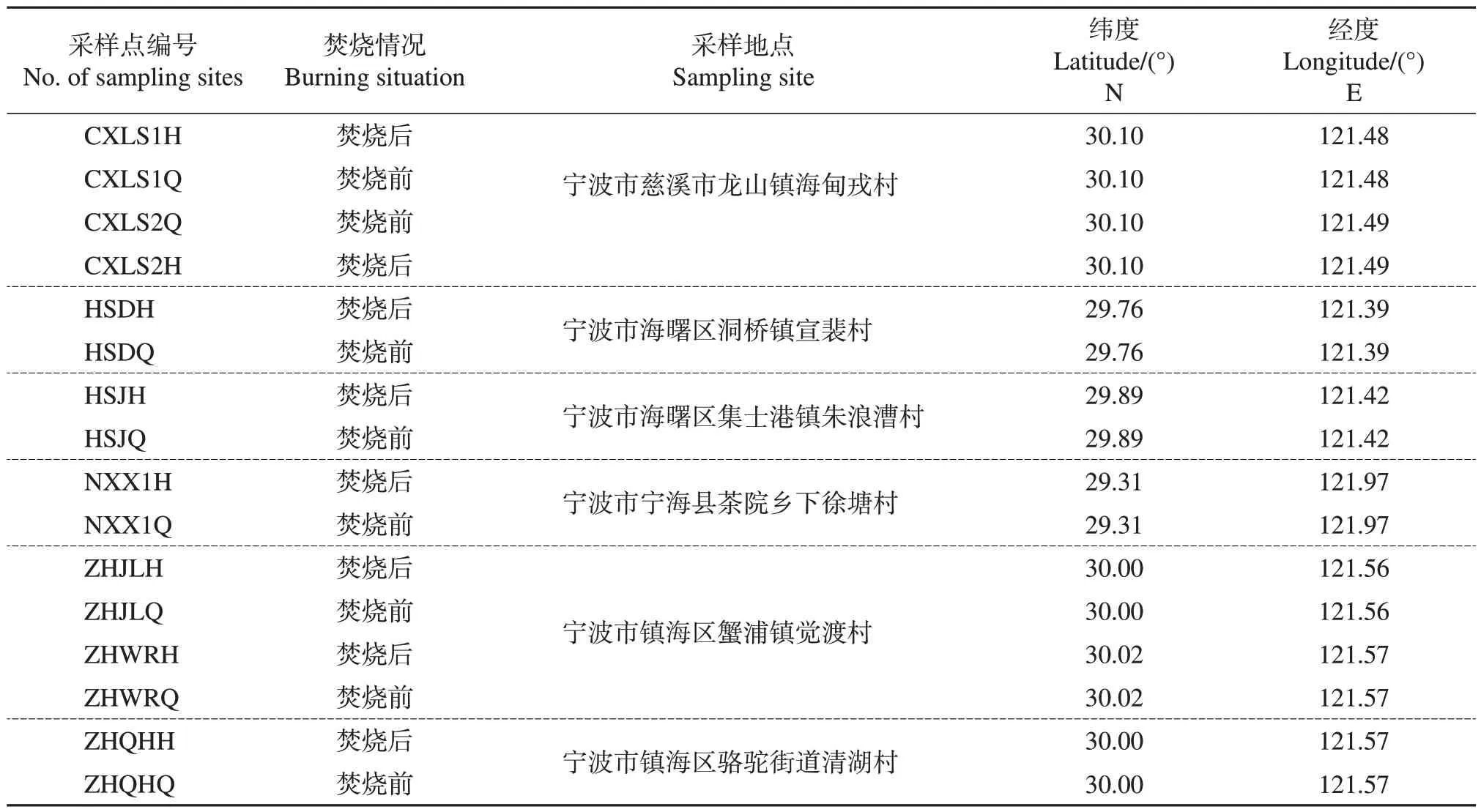
表1 研究区域和样品采集Table 1 Study area and sample collection
1.2 土壤理化因子测定
将土壤在105~110 ℃烘箱中烘干,称量其烘干前后的质量,获得土壤含水量。称量5.0 g 土壤于50 mL 离心管中,加入20.0 mL 去离子水,振荡30 min,离心后用pH 计测定上清液的pH 值。利用元素分析仪测定土壤中的总有机碳(total organic carbon, TOC)和总氮(total nitrogen, TN)含量。在过2 mm筛的土壤风干样中加入去离子水,水土体积质量比为5∶1,连续振荡4 h 后过滤,测定土壤溶解有机碳(dissolved organic carbon, DOC)含量[10]。用1 mol/L 中性乙酸铵溶液浸提土壤,采用原子吸收分光光度计测定土壤浸出液中的速效钾(available potassium, AK)含量[11]。基于《土壤农化分析》[12]中的方法,分别测定土壤有效磷(available phosphorus,AP)、硝态氮(NO-3-N)和铵态氮(NH+4-N)含量。
1.3 土壤和稻茬样品的DNA提取及宏基因组测序
DNA 提取和宏基因组测序由上海凌恩生物科技有限公司完成。具体方法为:利用土壤DNA提取试剂盒(美国Omega Bio-Tek公司)进行土壤和稻茬样品的DNA提取和纯化,然后用NanoDrop 2000超微量分光光度计(美国Thermo Fisher Scientific公司)进行定量。每个样品取1 μg DNA,用Covaris S220超声波破碎仪(美国Woburn公司)随机打断成长度约为450 bp 的片段,依次通过修复DNA 片段末端、添加多(poly)A尾、添加测序接头、纯化DNA片段、扩增聚合酶链反应(polymerase chain reaction, PCR)片段等步骤来完成整个文库制备。使用Qubit 3.0荧光定量仪对文库进行初步定量,然后稀释文库至2 ng/μL,并用定量聚合酶链反应(quantitative PCR, qPCR)对文库的有效浓度进行准确定量,以保证文库质量。所有样品采用Illumina NovaSeq 2000系统进行测序,读长模式为双端配对150 bp。每个样品的原始下机数据不小于20 Gb,并已上传至国家生物信息中心组学原始数据归档库(Genome Sequence Archive,https://bigd.big.ac.cn/gsa/),项目号为PRJCA015029。
1.4 致病菌、ARGs 和MGEs 注释
从病原体-寄主互作数据库[Pathogen-Host Interaction Database(PHI-base),http://www.phi-base.org]下载原核和真核致病菌物种基因数据,构建本地致病菌基因参考数据库(总共270种);此外,本研究还下载了ARGs-OAP v2.0 数据集(https://smile.hku.hk/SARGs)和MGEs数据集(https://github.com/KatariinaParnanen/MobileGeneticElementDatabase),分别构建本地ARGs 和MGEs参考数据库。针对土壤样品,先利用Trimmomatic软件对原始测序数据进行质控,获得高质量读长(clean reads)[13],然后用CLARK 软件[14](默认参数)将高质量读长比对到参考序列上,注释得到致病菌类群,最后用DIAMOND软件对ARGs 类型进行注释[e值<1×10-10,一致性(identity)值>0.8][15],用BLAST软件对MGEs进行注释(e值<1×10-8)[16]。针对稻茬样品,在对原始测序数据进行常规质控后,还使用Bowtie2 软件去除水稻寄主基因组序列,最终获得有效微生物测序数据,其致病菌、ARGs和MGEs的注释方法同土壤样品。
1.5 数据统计与分析
所有参数在秸秆焚烧前后样品间的差异通过双样本t检验进行统计学分析。通过R语言的picante包得到香农(Shannon)指数,以表征致病菌群落的α多样性。基于组间群落的Bray-Curtis距离,采用主坐标分析(principal coordinate analysis, PCoA)计算致病菌群落的β多样性,并用置换多元方差分析(ADONIS)检验其显著性。筛选超过70%的样品都存在的致病菌和ARGs组成信息,利用SparCC法构建致病菌和ARGs 的共存网络,网络可视化通过Gephi v0.9.2 软件完成。分别用R 语言的linkET 和ecodist包进行曼特尔(Mantel)检验和多元线性回归分析,以确定环境参数对致病菌群落结构和ARGs组成的影响程度。除了共存网络,其他图像的可视化均在R 4.1.3软件中进行。
2 结果与分析
2.1 秸秆焚烧对土壤理化性质的影响
土壤含水量、有机质含量和微量元素含量是土壤重要理化特性之一,直接或间接地与土壤肥力/质量相关。由图1可知,秸秆焚烧后,稻田表土的铵态氮、硝态氮含量及含水量有所下降,而有效磷、溶解有机碳、速效钾、总氮和总有机碳含量上升,其中秸秆焚烧前后速效钾含量变化显著。
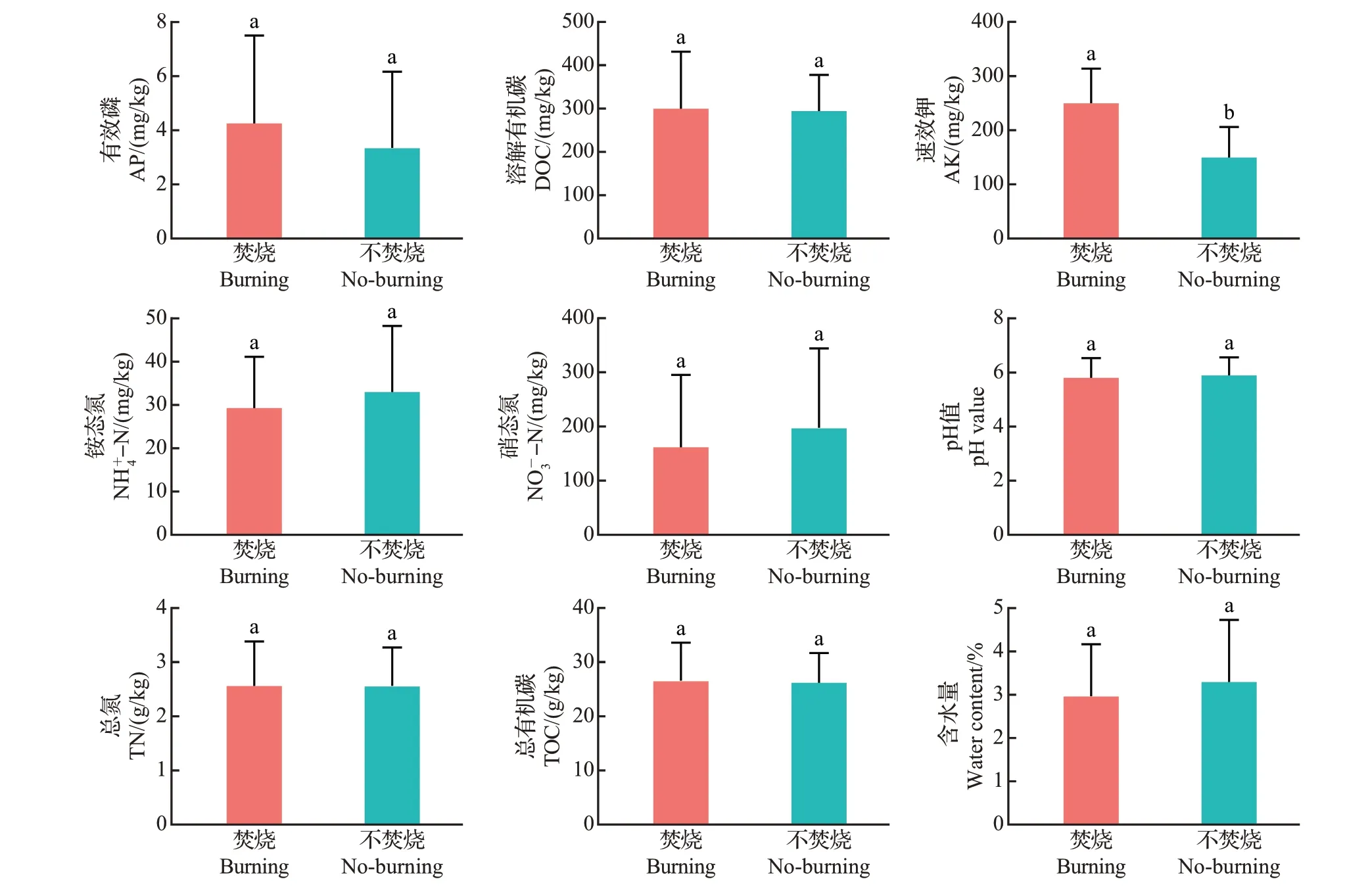
图1 秸秆焚烧前后土壤理化性质的变化Fig.1 Changes in soil physicochemical properties before and after straw burning
2.2 秸秆焚烧对土壤和稻茬中致病菌的影响
通过对浙江省宁波地区出现水稻白叶枯病的稻田土壤和稻茬样品进行生物信息学分析,得到土壤和稻茬中致病菌(包括原核和真核致病菌)的总相对丰度。如图2 所示:土壤样品中致病菌丰度占总微生物丰度的1%;然而,稻茬样品中致病菌丰度占总微生物丰度的18%,远高于土壤样品。其中,黄单胞菌属(Xanthomonas)、泛菌属(Pantoea)、假单胞菌属(Pseudomonas)是稻茬内主要的细菌致病菌属,镰刀菌属(Fusarium)是稻茬内主要的真菌致病菌属。针对出现白叶枯病的稻茬样品进一步进行检测,发现黄单胞菌占据主导地位,且其相对丰度在秸秆焚烧后显著降低(图3)。
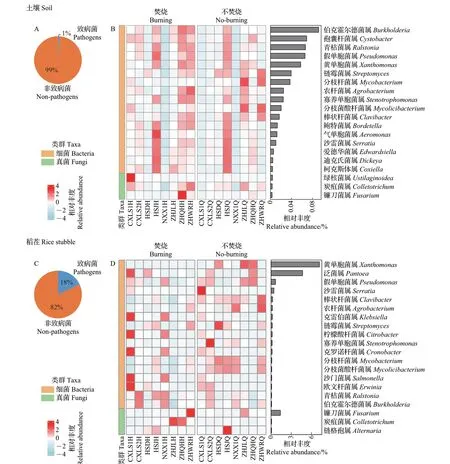
图2 秸秆焚烧前后土壤(A、B)和稻茬(C、D)致病菌丰度占比及主要致病菌属的相对丰度Fig.2 Proportions of pathogen abundances and the relative abundances of major pathogen genera in the soil (A, B) and the rice stubble (C, D) before and after straw burning

图3 秸秆焚烧前后黄单胞菌的相对丰度变化Fig.3 Changes in relative abundances of Xanthomonas and X.oryzae before and after straw burning
稻田致病菌多样性变化结果如图4 所示。由图4A、C可知,秸秆焚烧后土壤样品中香农指数有所下降,而稻茬样品中香农指数有所上升,但差异均不显著(P>0.05)。由图4B可知,主坐标1(PCoA 1)的贡献率为74.88%,主坐标2(PCoA 2)的贡献率为15.01%,秸秆焚烧前后土壤中致病菌群落组成未发生明显分离(R2=0.05,p=0.5)。由图4D可知,主坐标1(PCoA 1)的贡献率为31.25%,主坐标2(PCoA 2)的贡献率为22.88%,秸秆焚烧前后稻茬内致病菌群落组成存在差异但不显著(R2=0.12,p=0.09)。
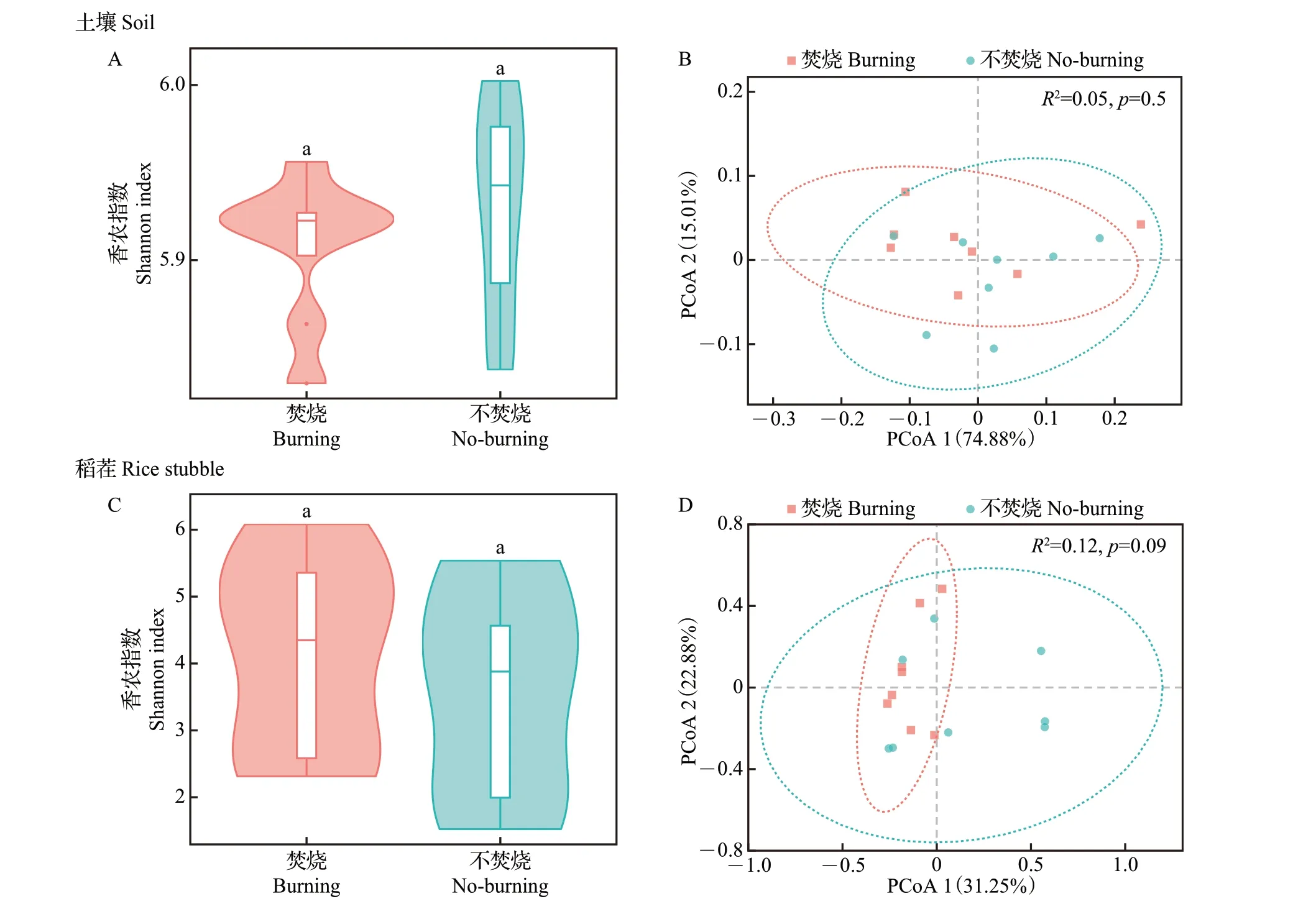
图4 秸秆焚烧前后致病菌群落组成差异Fig.4 Differences of pathogen community compositions before and after straw burning
2.3 秸秆焚烧对ARGs 和MGEs 丰度的影响
图5A~B、D~E 展示了土壤和稻茬内ARGs 种类和相对丰度的总体特征。ARGs在土壤和稻茬内的分布情况与致病菌的分布特征相似,即稻茬样品中ARGs的相对丰度高于土壤样品。秸秆焚烧后土壤中ARGs 丰度(单位细胞内ARGs 的拷贝数)为0.15,而稻茬内为2.7,主要包括多药、杆菌肽、大环内酯类-林可酰胺类-链阳性菌素类等ARGs 类型。同样地,ARGs 的相对丰度(每百万序列中的读长数)也得到类似的结果:秸秆焚烧后土壤中ARGs相对丰度为86,而稻茬内为702。此外,MGEs 在稻茬内富集,秸秆焚烧后土壤中MGEs相对丰度(每百万序列中的读长数)为34(图5C),而稻茬内为436(图5F)。在秸秆焚烧前后,土壤和稻茬中的ARGs相对丰度变化不显著,而秸秆焚烧后MGEs 的相对丰度在稻茬内显著降低。相关性分析结果(图6)表明,致病菌相对丰度与ARGs、MGEs 相对丰度呈正相关,且在土壤中致病菌和ARGs 的相对丰度表现出极显著正相关关系(r=0.65,p<0.01),而在稻茬内致病菌和MGEs的相对丰度表现出高度显著的正相关关系(r=0.95,p<0.001)。

图5 秸秆焚烧前后土壤(A~C)和稻茬(D~F)内抗生素抗性基因总体特征及其相对丰度变化和移动基因元件相对丰度变化Fig.5 Profiles of antibiotic resistance genes (ARGs), changes in relative abundances of ARGs and mobile genetic elements(MGEs) in the soil (A-C) and the rice stubble (D-F) before and after straw burning

图6 秸秆焚烧前后ARGs、MGEs相对丰度与致病菌相对丰度的相关性分析Fig.6 Correlation analysis on the relative abundances of ARGs and MGEs with the relative abundances of pathogens before and after straw burning
2.4 秸秆焚烧对致病菌和ARGs 共存的影响
进一步通过生态网络分析秸秆焚烧对致病菌和ARGs 共存特征的影响,结果如图7 所示。稻田土壤和稻茬内高频共存着上百种致病菌和ARGs(图7A~B、D~E)。基于网络结构和拓扑特征发现,秸秆焚烧前后土壤中致病菌和ARGs 的共存网络分别具有1 012个和1 454个连接,而稻茬中分别具有4 155个和7 029个连接。进一步研究发现,秸秆焚烧前后稻茬内致病菌和ARGs 的正相关(正向)连接数占比分别为41.8%和41.5%。这些结果表明,致病菌和ARGs 虽然存在紧密关联,但主要以负相关为主,且秸秆焚烧减弱了致病菌与ARGs的正相关关系(如共生关系)。此外,秸秆焚烧后水稻黄单胞菌(X.oryzae)所在网络模块的拓扑特征也发生了变化(正相关连接数占比减少),表明秸秆焚烧会对水稻白叶枯病病原菌和ARGs的共存网络产生影响。进一步研究发现,与致病菌共存的主要类型是耐多药ARGs。通过曼特尔(Mantel)分析发现,对土壤中致病菌和ARGs 共存影响较大的理化参数是含水量、pH值和总氮含量(图7C),而对稻茬内致病菌和ARGs共存影响较大的是速效钾和有效磷含量(图7F)。
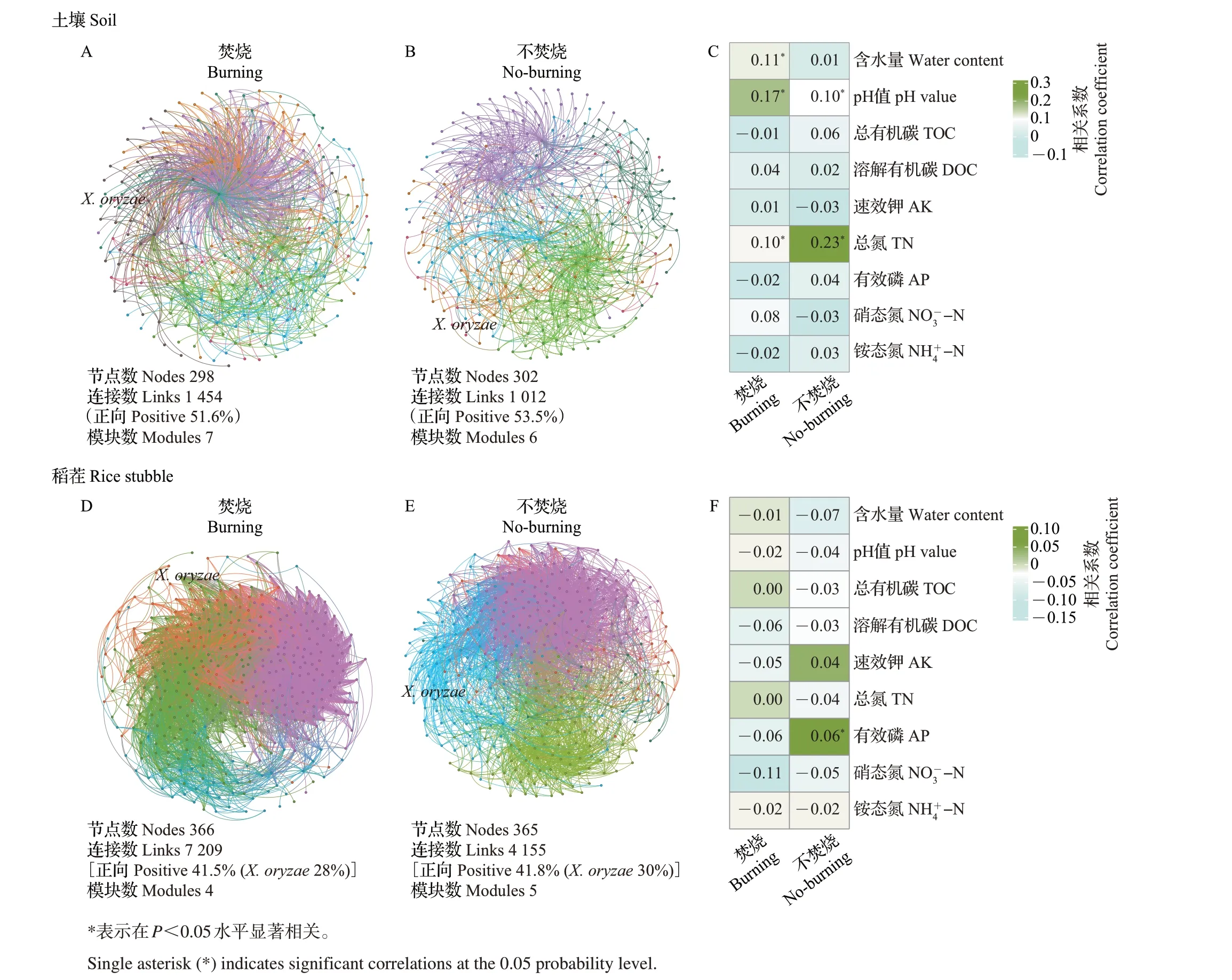
图7 秸秆焚烧前后土壤(A~C)和稻茬(D~F)内致病菌和ARGs共存分析及其共存网络拓扑特征与土壤理化因子的相关性分析Fig.7 Coexistence analysis of pathogens-ARGs and correlation analysis between co-occurrence network topology features and environmental factors in the soil (A-C) and the rice stubble (D-F) before and after straw burning
2.5 稻田生物复合污染的重要驱动因子
曼特尔(Mantel)检验结果揭示了土壤理化因子与致病菌群落结构和ARGs 组成的关系(图8)。在土壤中,土壤理化因子对致病菌群落结构变化的影响较小,而对ARGs组成的影响较大。其中,含水量(r=0.642,p<0.01)、pH 值(r=0.582,p<0.05)、总有机碳含量(r=0.325,p<0.05)、总氮含量(r=0.570,p<0.01)和有效磷含量(r=0.311,p<0.05)与ARGs组成密切相关。在稻茬中,溶解有机碳含量(r=0.201,p<0.05)显著影响致病菌群落结构,且溶解有机碳含量(r=0.275,p<0.05)和速效钾含量(r=0.312,p<0.05)与ARGs 组成密切相关。多元线性回归模型分析结果总体上与曼特尔检验结果一致,土壤理化因子对土壤中致病菌群落结构和ARGs组成具有显著贡献(r2=0.451,p=0.006;r2=0.527,p=0.002),而对稻茬内致病菌群落结构和ARGs组成的影响不显著(r2=0.090,p=0.61;r2=0.189,p=0.14)。
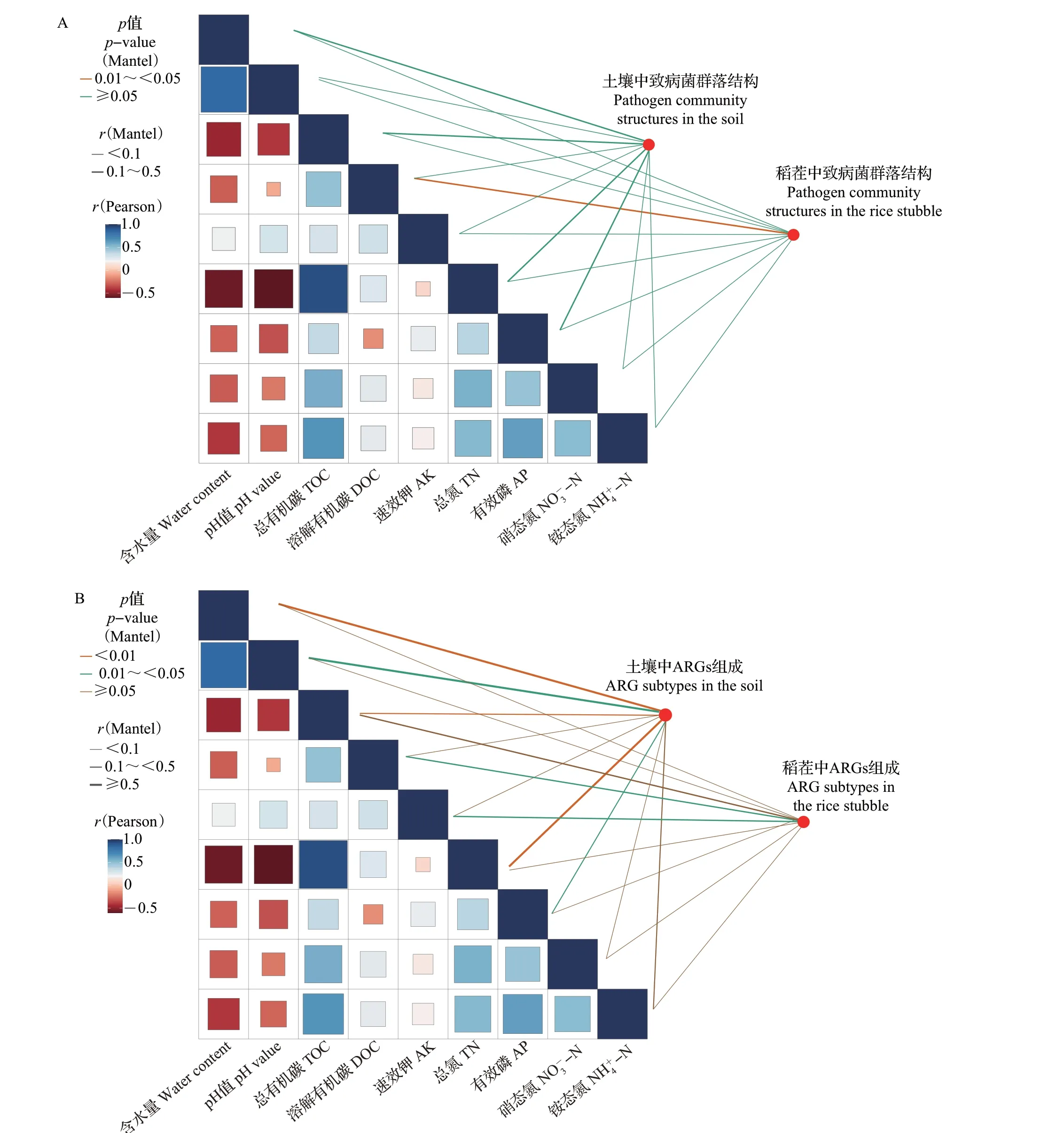
图8 土壤和稻茬内致病菌群落结构(A)和ARGs组成(B)与土壤理化因子的相关性分析Fig.8 Correlation analysis of pathogen community structures (A) and ARG subtypes (B) in the soil and rice stubble with the soil physicochemical factors
3 讨论
从2010年全面禁止焚烧秸秆以来,秸秆还田成为秸秆资源的主要利用模式。尽管这种模式对环境保护起到了重要作用,但不利于作物病害的控制,因为焚烧秸秆能降低致病菌在田间的接种量,使农作物保持低感染率[17]。近年来,浙江省宁波地区水稻种植区出现连片的白叶枯病,且不断扩散蔓延,呈逐年加重态势[4]。因此,亟须就不同秸秆管理方式对作物的影响进行重新评估。本研究对比了秸秆焚烧前后稻田致病菌、ARGs 和MGEs 的变化情况,结果表明,秸秆焚烧对稻茬内致病菌群落结构、ARGs组成及其生态网络产生了重要影响,这为客观评价农田秸秆焚烧提供了一定的参考。
土壤是多种致病菌的储存库。尽管致病菌在绝大部分时期具有较低的丰度和活性,但当其在土壤中的累积量达到植物病害发生的条件时,则能实现指数增长。例如,一旦黄单胞菌成功侵染水稻,该菌种便会在秸秆内大量富集[4]。这也提示收割自白叶枯病暴发的秸秆需要经过有效的杀菌处理才能进行还田利用,否则粗放式的秸秆还田势必加剧下茬作物的染病风险。尽管秸秆焚烧能够显著降低稻田中黄单胞菌的丰度,但其遏制下茬作物发病的程度还需要长期定位试验来加以验证。由于焚烧效果具有不可预测性,焚烧处理可能以休眠形式降低了土壤中病原菌的接种量,然而并未完全根除[18]。同时,焚烧效果取决于焚烧强度、秸秆数量和质量以及焚烧持续时间[19-20]。除了黄单胞菌,菠萝泛菌(Pantoea ananatis)也能引起类似白叶枯病症状的水稻病害[21-22]。在本研究中,稻茬内菠萝泛菌的丰度仅次于黄单胞菌,表明近些年国内暴发的白叶枯病可能同时受到黄单胞菌和菠萝泛菌的侵染,从而导致传统防治药剂的防控效果较差。因此,开展水稻白叶枯病病原菌的鉴定和防治药剂的筛选等技术研究刻不容缓。
有效防治植物病害还与致病菌体内是否携带抗生素抗性基因(ARGs)密切相关。本研究发现,在水稻白叶枯病暴发时,稻茬内ARGs 相对丰度较低,而当秸秆焚烧后,稻茬内ARGs相对丰度有所升高,进而提升了致病菌和ARGs 共存网络密度。这与其他秸秆处理方式下ARGs的相对丰度变化有相似之处。魏华炜[23]指出,在施用秸秆处理的肥料后,小葱地上部的ARGs丰度上升,进一步研究发现,内生细菌是促进植物体内ARGs 传播的主要驱动因子。有研究表明,秸秆还田能够为土传植物致病菌的生长、繁殖和积累提供适宜的环境[24],例如,小麦秸秆还田会提高下茬小麦赤霉病病穗率和病情指数,水稻秸秆还田会提高下茬水稻稻瘟病、鞘腐病和纹枯病的病情指数,同时显著影响作物体内的ARGs 丰度[25]。尽管本研究中稻茬内ARGs 与致病菌寄主共存关系在秸秆焚烧后有所加强,但致病菌和ARGs 共存网络的正相关连接数占比有所降低,并且与致病菌相对丰度呈正相关的移动基因元件(MGEs)相对丰度显著下降,表明秸秆焚烧将减小基因水平转移的性能,有可能遏制耐药致病菌的形成,并降低通过致病菌传播抗性基因的风险。为了探究土壤和稻茬中可能形成的致病菌和ARGs的生物复合污染风险,需要分离鉴定潜在的耐药致病菌,进一步验证生物信息学分析的结果。
土壤理化性质(含水量、pH 值、温度、有机质含量等)可以显著影响土壤中微生物群落结构的多样性,从而诱导ARGs的传播和富集[26]。在本研究中,相关性分析结果表明,土壤中致病菌的多样性与土壤理化性质关系密切。秸秆焚烧对环境因子的改变在一定程度上驱动了土壤中致病菌群落的生态演替,降低了黄单胞菌丰度,从而降低下茬病菌的侵染率。秸秆焚烧后土壤中致病菌的多样性有所下降,与李明等[9]的研究结果一致。值得注意的是,多个理化参数(含水量、pH值及总有机碳、溶解有机碳、速效钾、总氮、有效磷含量)对ARGs组成具有显著影响,与已有的相关研究结果一致。例如,tet基因的绝对丰度与土壤有机质含量具有显著相关性[27];土壤pH值和含水量通过影响养分有效性和微生物活性干扰ARGs 的传播、富集、去除和持久性[28-29]。这也给未来绿色农业发展提出了新的挑战:在农业实践中(如秸秆还田、有机肥施用等),不仅要防控病害的暴发,而且要密切关注抗性基因与致病菌互作的危害,避免耐药致病菌的形成和富集。
4 结论
本研究采用宏基因组学技术对浙江省宁波地区白叶枯病暴发稻田的致病菌群落、ARGs和MGEs在秸秆焚烧前后的组成、丰度、生态网络特征变化进行了研究。在染病稻田中稻茬内致病菌相对丰度远高于土壤,其中黄单胞菌、菠萝泛菌为优势菌种。秸秆焚烧导致稻茬内黄单胞菌的相对丰度急剧降低。在秸秆焚烧后,尽管土壤和稻茬内的ARGs相对丰度有所提升,但致病菌和ARGs的正相关共存关系占比减小,而且MGEs 相对丰度显著降低,从而可能降低通过致病菌传播抗性基因的风险。土壤中ARGs组成变化与含水量、pH值及总有机碳、总氮、有效磷含量相关,而稻茬内的致病菌群落结构和ARGs组成变化与溶解有机碳含量密切相关。本研究揭示了秸秆焚烧对白叶枯病病原菌黄单胞菌的抑制作用,为进一步研究不同秸秆管理模式对稻田生物复合污染的影响提供了参考,对在实践中加强对水稻白叶枯病的防控具有指导意义。本研究结果也提示,未来针对作物病原菌的研究不仅要关注其组成和丰度的变化,而且要进一步聚焦于菌种的绝对丰度、活性及耐药性的变化。
猜你喜欢
现代农业科技(2020年17期)2020-09-17
现代农业科技(2018年19期)2018-12-21
安徽农学通报(2018年11期)2018-07-09
上海农业学报(2017年4期)2017-04-10
现代食品(2016年24期)2016-04-28
中华老年口腔医学杂志(2016年5期)2016-03-01
上海蔬菜(2016年5期)2016-02-28
华南农业大学学报(2015年5期)2015-12-04
果树资源学报(2015年3期)2015-09-26
中国质量与标准导报(2014年4期)2014-03-11
