
利用基因编辑技术创制福香占糯稻新种质
2024-01-11 07:06魏林艳樊佳星王进兰谢鸿光魏毅东张建福
福建农业科技 2023年10期
魏林艳,樊佳星,张 敏,王进兰,谢鸿光,魏毅东,张建福*
(1.福建省农业科学院水稻研究所/水稻国家工程实验室, 福建 福州 350003;2.福建农林大学农学院, 福建 福州 350002)
糯稻(glutinous rice)作为中国重要的农作物之一,因其米粒含有糯性淀粉而备受青睐。糯稻不仅是许多地区的传统主食,还可以制作出粽子、酒酿、汤圆等多种传统小吃。目前,抗瘟糯稻新品种相对匮乏,据2022、2023年福建省初审通过主要农作物品种目录可知,糯稻新品种的仅2个、且都不抗稻瘟病。因此,需要创制抗瘟糯稻新种质来应对不断变化的生态环境和生产环境。糯稻一般指胚乳直链淀粉含量低于2%的水稻[1],而颗粒结合淀粉合酶(Granule-Bound Starch Synthase,GBSS)基因Waxy(Wx)是控制直链淀粉含量的主效基因[2]。因此,Wx基因的多态性在很大程度上决定了直链淀粉含量的多样性[3]。过去的研究发现存在多种Wx自然等位基因,这些基因控制着水稻直链淀粉含量从低于2%到高于25%的范围[3-5]。其中,Wx基因的缺失突变体Wx导致直链淀粉下降到2%以上,是稻米糯性的主要效应基因[6-7]。
CIRPSR/Cas9系统是近年来迅速发展的高效、准确、便捷的基因编辑技术。与转基因不同,基因编辑技术可以在不引入外源基因的情况下对目标基因进行敲除突变,可用于水稻品种的精确改良。福香占(闽审稻20200011)是福建省农科院水稻研究所于2020年新育成的优质、抗瘟、长粒籼型常规稻品种,具有较好的推广潜力。本研究的目标是利用基因编辑技术快速创制福香占糯稻材料,为抗瘟病糯稻育种提供新的种质资源。
1 材料与方法
1.1 试验材料
CRISPR/Cas9敲除载体PHUE411由中国农业大学生物学院陈其军教授惠赠;农杆菌菌株EHA105由福建省农业科学院水稻国家工程实验室保存;受体材料籼稻福香占由福建省农业科学院水稻国家工程实验室提供。所有转基因材料种植于福建省农业科学院转基因试验基地,采用常规种植与水肥管理。
1.2 载体构建
将Wx基因的登录号LOC_Os06g04200在CRISPR-PLANT(https://www.genome.arizona.edu/crispr/CRISPRsearch.html)进行靶点筛选,选择内含子与外显子交界处的靶点作为候选位点,设计该靶点的CRISPR/Cas9敲除引物(Wx-sgRNA-F:AATAATGGTCTCAGGCGAGCCTCGAGTGCTGC-CTGCGTTTTAGAGCTAGAAATAGC,Wx-sgRNA-R:ATTATTGGTCTCTAAACGCAGGCAGCACTCGAGGCTCGCTTCTTGGTGCC)随后以sgRNA质粒元件pCBC-MT1T2为模板PCR扩增目的片段,采用KOD-FX高保真酶(东洋纺生物科技有限公司)进行扩增,扩增体系见表1。

表1 PCR扩增体系及流程
随后将产物进行凝胶电泳检测并回收,将产物与敲除载体PHUE411通过金门法进行连接后转化大肠杆菌感受态,所用T4连接酶以及BsaI内切酶购自New England Biolabs(NEB),连接步骤见表2。获得最终有正确敲除片段的重组质粒后,利用冻融法转入农杆菌转化后用于水稻转化,农杆菌介导的水稻转化法参照本实验室建立体系进行[8]。
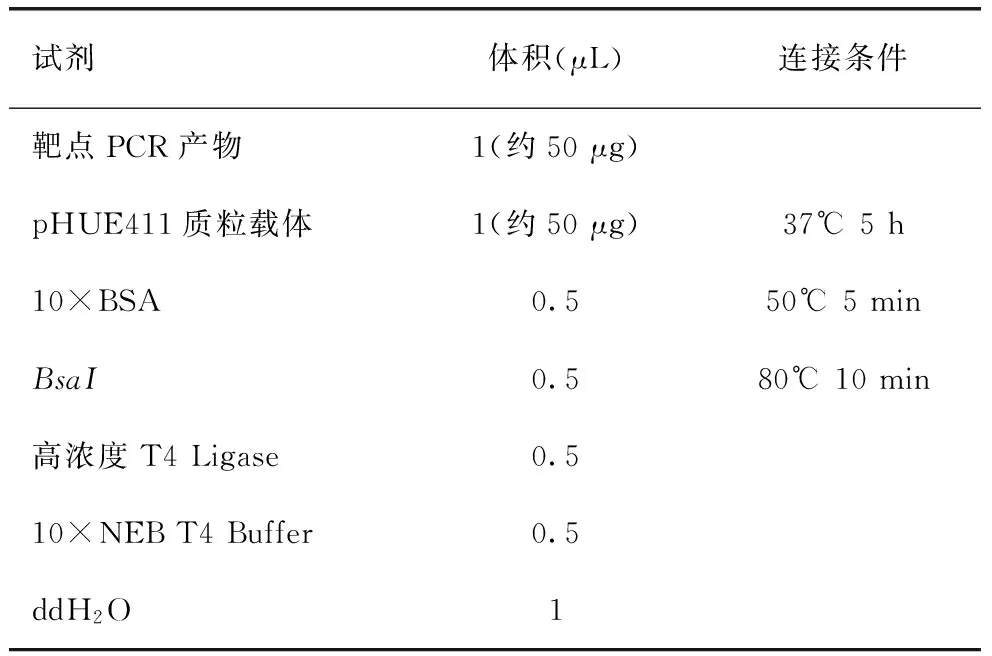
表2 金门连接反应体系
1.3 基因编辑载体的遗传转化及转基因植株分子检测
基因编辑载体的遗传转化采用的是农杆菌介导的方法,具体参考文献[9]。剪取样品幼嫩叶片,用CTAB法提取DNA。针对Wx基因突变型分析,采用PCR扩增子测序的方法,引物采用Wx基因靶点侧翼引物Wx-seq-F:CCTGGTAGGAGATGTTGTGGATG和Wx-seq-R:AGGTTTTTCCATTGCTACAAGCGT,所得产物大小735 bp。PCR扩增反应体系 :DNA模板50 ng,2×TaqPCR Mix 12.5 μL,10 mmol·L-1引物各 1 μL ,Nuclease-Free Water 定容至 25 μL。PCR扩增条件:预变性94℃ 5 min,扩增35个循环(94℃ 15 s,55℃ 30 s,72℃ 30 s)。扩增产物在1%琼脂糖凝胶电泳后送测序公司(铂尚生物技术有限公司)进行测序。测序结果利用SnapGene软件进行初步比对,杂合体测序双峰采用DSDecodeM[10]进行解析。
1.4 突变体植株田间农艺性状调查
田间材料按照《国家水稻品种试验观察记载项目、方法标准》进行株高、分蘖数、株高、千粒重等农艺性状调查进行考查,每个株系重复5~8个单株,获得数据通过Graphpad进行统计分析。
1.5 突变体株系稻米淀粉含量测定
总淀粉含量按国标 GB5006-1985《谷物籽粒粗淀粉测定法》所述方法进行,直链淀粉含量测定依据国标GB7648-87《水稻、玉米、谷子籽粒直链淀粉测定法》所述方法进行,所有样品测定3个重复,数据采用Graphpad进行统计分析。
1.6 突变体株系稻瘟病抗性鉴定
稻瘟病抗性鉴定选择福建省上杭县稻瘟病高压病圃中进行,每个品种按小区播种,设置3个重复,四周设置感病诱发行。在苗期观察并记录苗瘟发病情况,将存活植株移栽到大病圃种植至成熟,其间观察叶瘟和穗瘟的发生情况。苗瘟、叶瘟和穗颈瘟分级按国际水稻所0~9级标准进行判定。
2 结果与分析
2.1 基因编辑载体的构建与转化
通过靶点在线评估以及突变类型分析,选取Wx编码区第5外显子上游边界区域作为CRISPR靶点(图1A),设计通过金门连接法将Wx的基因编辑靶点串联重组至CRISPR/Cas9载体PHUE411上(图1B)。通过农杆菌的水稻转化法将Wx基因编辑载体转入受体福香占,共获得T0代独立克隆12个(图2)。

注:A为Wx基因结构图,白色部分为非翻译区,绿色部分为编码区,折线部分为内含子区,红色三角形标注为本研究设计的Wx靶点。B为Wx敲除载体结构图,LB/RB为T-DNA的左/右臂,OsU3p/TaU3p为U3启动子区,蓝色区域为含有Wx靶点的sgRNA序列,U3t为终止子区,Cas9/HygR分别表示Cas9蛋白/潮霉素抗性蛋白表达盒图1 Wx基因基因编辑载体的构建Fig.1 Construction of Wx Genome Editing Vector

注:M1~12为12个独立转化克隆图2 Wx基因编辑T0代材料Fig.2 T0 Generation Materials of Wx Genome Editin
2.2 利用基因编辑获得Wx基因突变植株
提取T0植株DNA进行靶点突变分析,结果表明12个克隆均检测到编辑事件(图3),其中双等位突变10株,单等位突变2株。详细的碱基变异见(表3),由于所选靶点位于外显子和内含子交界处,根据突变类型不同,可能会形成Wx基因内含子滞留或者编码区移码两种类型。通过突变类型分析,结合田间农艺性状观察,确定了2个克隆M1和M9为后续研究材料。M1和M9在外显子区分别插入和删除了一个碱基“G”,属于移码突变,会形成无功能(Loss-of-function)的蛋白变体,可作为糯稻候选材料进行种植。

注:参考序列为福香占Wx基因靶点周边序列,蓝色标识为靶点PAM序列,红色标识为靶点区域,小写字符为内含子序列,大写字符为外显子序列,三角形标注Cas9蛋白的切点(PAM序列上游-3/-4之间);M1~3为3个独立转化克隆测序峰图,红色箭头标注出现突变的峰图起始位点图3 Wx基因编辑材料突变检测Fig.3 Mutation Detection of Wx Genome Editing Materials

表3 T0代植株突变基因型
2.3 Wx基因编辑植株的表型鉴定
由于T0代植株为双等位突变,为了获得纯合突变体植株,对T0代收获的种子进行了突变型分析,M1和M9移码突变体都筛选到纯合的无功能突变型种子(图4)。将这些种子扩繁提纯后获得两个株系G-1(M1子代)和G-2(M9子代),对突变体株系进行农艺性状考查和糙米外观品质检测,结果表明突变体株的抽穗期、株高、穗长与福香占相似,而产量性状略有不同,其中G-1和G-2千粒重略有下降,G-1差异达到显著水平,但两个株系的总体产量与对照相比,无显著差异(表4)。此外,通过外观品质可以看出福香占透明度较高,两个基因编辑的株系的透明度较差,呈糊化状态。可初步判断两个株系均为糯稻材料(图5)。
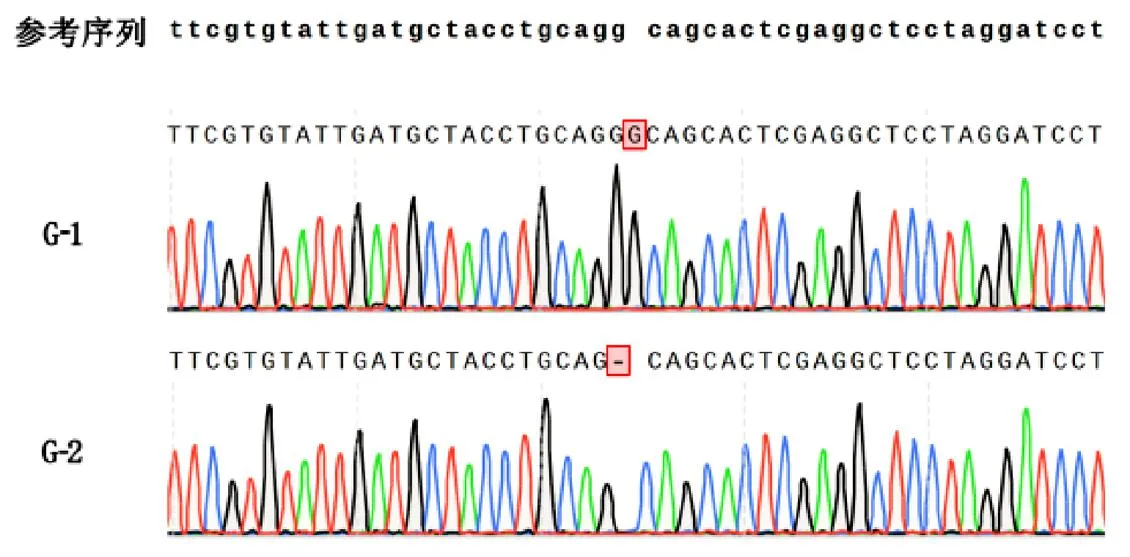
注:参考序列为福香占Wx基因靶点周边序列,G-1和G-2纯合突变体序列(上)和测序峰图(下)。图4 Wx纯合缺失突变体基因型鉴定Fig 4 Genotype Identification of Wx Homozygous Deletion Mutants
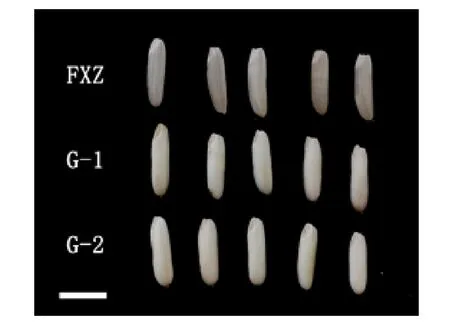
注:FXZ为亲本福香占,G-1和G-2为2个缺失突变株系,标尺为1 cm。图5 Wx缺失突变体糙米外观形态Fig.5 Appearance of Wx knockout Mutant Brown Rice
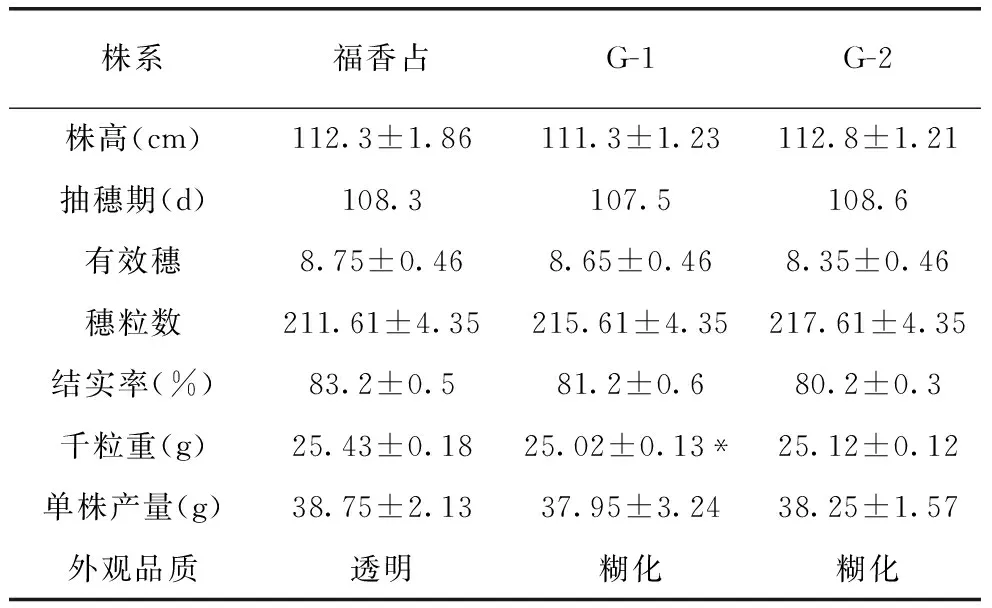
表4 突变体株系农艺性状调查
2.4 Wx基因编辑植株的直链淀粉含量测定
为了进一步验证突变体植株的糯稻表型,测定了福香占和突变系G-1和G-2的稻米直链淀粉和总淀粉含量。由表5可知,亲本福香占的直链淀粉含量为16.3%,2个纯合株系的直链淀粉含量分别为1.64% 和1.81%,符合糯米国家标准的2%范围内。综上所述,本试验通过基因编辑技术成功获得福香占糯稻新材料。

表5 突变体株系淀粉含量测定
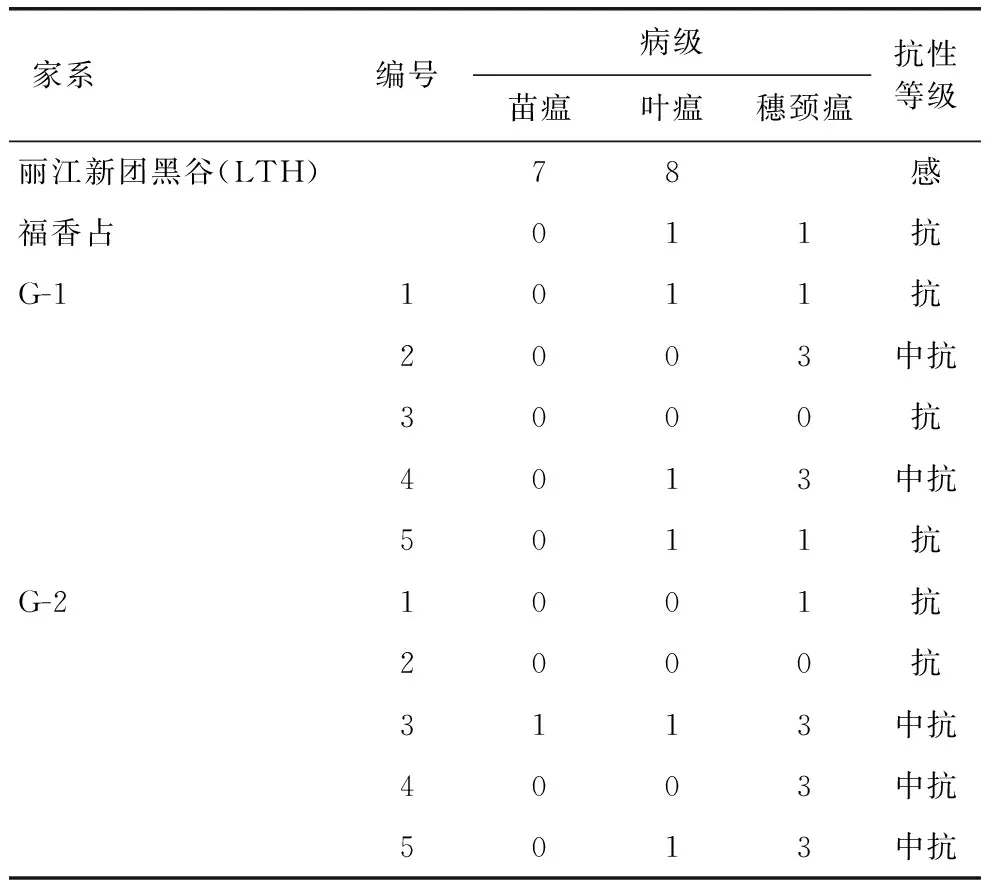
表6 福香占糯稻家系稻瘟病抗性鉴定
2.5 福香占糯稻材料的稻瘟病抗性鉴定
从G-1和G-2株系中各选了5个综合性状好的单株进行扩繁。福建稻瘟病高致病菌种对10个家系材料进行稻瘟病接种鉴定。结果表明,10个株系均有稻瘟病抗性,其苗瘟和叶瘟均为 0~1级,株系G1-1/3/5、G2-1/2穗颈瘟 0~1 级,综合抗性评价为抗稻瘟病,株系G1-2/4、G2-3/4/5穗颈瘟3级,综合抗性评价为中抗稻瘟病(表 6)。以上结果表明,利用基因编辑改良福香占的糯性,能基本保持其稻瘟病抗性。
3 讨论与结论
随着近年来功能基因组学和基因编辑技术的迅猛发展,基因编辑已经广泛应用于作物性状改良和种质创制,在品质、株型、抗性等多个方面取得了显著进展[11-15]。基因编辑作为一种不引入外源片段的靶向改良方法,具有后代遗传稳定和多性状快速改良的优势[16]。在本研究中,我们选取了优质抗瘟常规稻福香占进行基因编辑,通过田间的选择,突变株系在株高、穗数、生育期等农艺性状方面没有显著差异。同时,这些突变体还保持了对稻瘟病的抗性特性,适合用于改良现有的潜力品种,避免了烦琐的回交导入选育过程。
根据本研究的结果,Wx基因的敲除对性状产生的负面效应并不显著。然而,突变体中的千粒重大约降低了1.4%。这可能是由于Wx基因参与胚乳淀粉合成途径的重要性,其突变会引起籽粒灌浆的生理和分子变化。此前的研究也观察到敲除Wx基因后千粒重略微下降的现象[7]。然而,其他类似的Wx基因编辑研究中,大部分材料的千粒重没有发生下降,甚至有些突变体略微升高[6,17-19],这可能是由于不同遗传背景产生的效应差异。此外,福香占的突变系穗粒数略有升高,因此总体产量并没有显著变化。然而,要实现福香占糯稻种质的真正育种应用,还需要进行一系列的工作:除获得基因编辑的安全证书外,还需要对其环境适应性、抗逆性和其他生产特性进行全面评估,以更全面地评估Wx基因敲除的影响。
值得注意的是,随着基因编辑技术的进一步发展,已经开发出更先进的工具,可以精确地降低或激活特定基因的表达,从而创制出目标性状梯度突变的材料。以Wx基因为例,通过对启动子和5′UTR区的定点突变,可以产生直链淀粉梯度变化的种质,如高直链淀粉的米粉稻和松软口感的软米品种等,极大地扩展了基因编辑在育种上的应用[20-22]。这些编辑位点同样可以进行编辑,创制出更丰富多样的福香占抗瘟衍生系,以满足不同市场需求。在未来的研究中,我们将继续探索基因编辑技术在水稻种质创新中的应用,推动水稻生物育种的创新发展。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
中国粮油学报(2019年4期)2019-07-12
四川农业科技(2019年5期)2019-07-01
中央民族大学学报(自然科学版)(2017年4期)2017-10-22
中国塑料(2016年2期)2016-06-15
浙江农业科学(2016年11期)2016-05-04
浙江柑橘(2016年1期)2016-03-11
福建稻麦科技(2015年1期)2016-01-06
农产品市场周刊(2014年39期)2014-12-19
食品科学(2013年22期)2013-03-11
