
丝状真菌分生孢子及其核型相关基因的研究进展
2024-01-11 07:07陆娇娇郑珊珊王思琦施碧红
福建农业科技 2023年10期
吴 君,陆娇娇,郑珊珊,王思琦,施碧红,2*
(1.福建师范大学生命科学学院, 福建 福州 350117; 2.福建师范大学工业微生物与发酵技术国家联合工程研究中心, 福建 福州 350117)
丝状真菌种类繁多,包括曲霉、木霉、毛霉和青霉等,它们与人类活动息息相关,一些被广泛用于工业生产发酵食品、工业酶和次级代谢产物,如青霉素、灰黄霉素生产者产黄青霉Penicilliumchrysogenum[1]、灰黄青霉Penicilliumgriseofulvum[2];各种酶制剂、有机酸等的生产菌米曲霉Aspergillusoryzae[3]、黑曲霉Aspergillusniger[4]等。同时还有不少种类是重要的植物病原菌,少数可侵染动物或人类,引起真菌感染。由于丝状真菌基因组中含有大量与生物降解、初级和次级代谢、转录调控和细胞信号转导有关的基因,且具有强大的合成和分泌大量蛋白质的能力,因此是生产异源蛋白的理想宿主[5]。
曲霉属和青霉属是已报道的产生外代谢产物最丰富的两类丝状真菌,曲霉属的每个物种平均产生外代谢物5.77种,青霉为3.77种,且该数据仍有可能被低估[6]。因此曲霉和青霉中的一些物种是作为细胞工厂或工业微生物育种的重要材料。曲霉和青霉菌的营养体具有发达的菌丝,有隔膜和分枝。其生活史以无性繁殖为主,产生具有独特形态结构的分生孢子梗和大量的无性孢子-分生孢子。其有性孢子为子囊孢子,但在曲霉和青霉这两个重要的属中,有些种很少产生或从不产生子囊果,而以分生孢子进行繁殖。然而分生孢子核型具有多样性,少数为单核、多数为多核孢子。尽管真菌多核性的生物学优势是众所周知的,多核的分生孢子不仅具有较高的发芽效率外,还有较强的生存能力和对紫外线辐射、冻融处理的抗性[7],因此可以有效提高丝状真菌在压力环境条件下的生存效率。多核特性对真菌进化亦至关重要,通过异核体导致不同遗传性的核融合而产生新的基因型,使得真菌种群不断进化得以适应复杂的环境。但是,多核特性亦会削弱遗传操作产生的表型效应,如异核转化子导致蛋白表达水平的下降等。开展真菌分生孢子核型研究对解析丝状真菌细胞生理生化、遗传变异机制以及对真菌遗传育种和病原真菌防治都具有重要意义,因此本文就曲霉等丝状真菌分生孢子的发育及其核型变化调控相关基因的功能进行归纳,为更好地开展相关物种的遗传操作提供理论基础。
1 丝状真菌分生孢子发育过程的基因调控
分生孢子是绝大多数曲霉、青霉属丝状真菌的无性生殖细胞。分生孢子产生于由菌丝分化而形成的分生孢子梗上,分生孢子梗始于菌丝的厚壁足细胞生出并向空中伸长,产生气生茎。茎停止延伸后开始膨胀,形成囊泡,在囊泡膨胀阶段,囊泡内进行大量核分裂复制,接着出芽产生瓶梗,囊泡内细胞核迁移到瓶梗,瓶梗继续进行出芽生殖,通过重复不对称分裂产生分生孢子链,并且在出芽点与孢子连接处形成隔膜[8]。当囊泡膨胀和分生孢子伸长时,细胞核会同时经历几次有丝分裂,并逐步从菌丝体迁移到孢子中。总之,在分生孢子形成过程中,菌丝中的细胞核经过一系列核分裂、核迁移到形成隔膜,最后形成菌丝分支。分生孢子的形成过程受到多种基因调控(表1)。
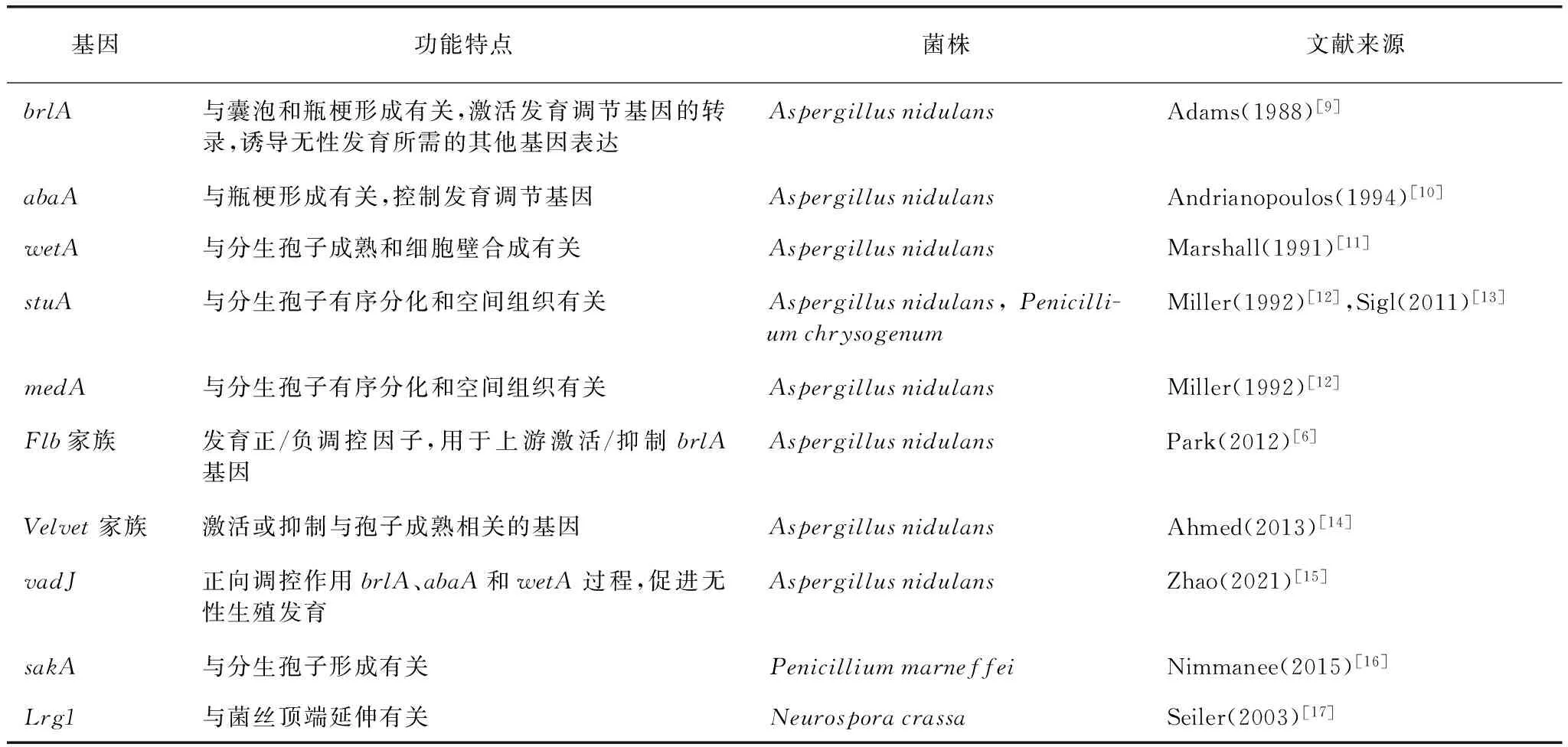
表1 常见的控制分生孢子形成的基因

表2 常见的控制核分裂及核迁移的基因
分生孢子发育通常受brlA、abaA、wetA、stuA和medA等基因的协同调控[6,18],它们的表达产物直接促进分生孢子梗的形成(表1)。分生孢子发育的关键步骤是激活编码C2H2锌指转录因子的基因brlA,该基因还诱导无性发育所需的其他基因abaA和wetA的表达。abaA是分生孢子瓶梗正常形成所必需的,abaA突变体导致珠状孢子的生成,abaA过表达导致菌丝生长终止和细胞空泡化。wetA是分生孢子特异性基因的(转录)调控因子,由brlA和abaA顺序表达激活,wetA突变体无法积累分生孢子特异性基因的mRNA。brlA、abaA和wetA这三个基因被定义为一个中央调控通路,通过brlA、abaA和wetA基因的顺次激活决定分生孢子形成和成熟的特异性基因表达[9]。stuA和medA是调控分生孢子有序分化和空间组织所必需的,这两个基因中的任一突变都会导致空间紊乱的分生孢子梗。
2 丝状真菌分生孢子核型变化过程的基因调控
丝状真菌具有多种生殖方式,一些丝状真菌在其生活史中不发生有性生殖过程,其细胞核在有丝分裂过程中会经历准性重组事件,造成异核体、同核体、重组单倍体或二倍体的存在,故其菌丝和分生孢子中可以保持单核或多核状态。大多数丝状真菌的分生孢子具有核异质性,如Aspergillusflavus、Aspergillusoryzae、Neurosporacrass、Peniclliumchrysogenum及Microsporumgypseum等,且在其整个生命周期中具有单核、多核或两者共存状态[19]。多核特性使丝状真菌适应更广泛环境,但同时也增加了对其进行遗传分析及菌种改良的难度,如隐性突变体的获得等。真菌细胞核动力学是多核形成的重要基础,也是维持分生孢子正常发育的关键,涉及微管、肌动蛋白、动力蛋白、驱动蛋白和调节因子组成的调控网络。有关分生孢子核型变化过程基因调控的机制研究在不断进行中。
2.1 核分裂过程的基因调控
丝状真菌分生孢子形态与生长和核分裂密切相关。分生孢子发育与细胞有丝分裂周期相偶联,在丝状真菌形成的分生孢子中还含有一些停滞在有丝分裂G1期的单核孢子,这些孢子经历新一轮萌发,顶端开始极化生长,同时细胞核再次进入有丝分裂周期。核分裂在A.nidulans由两种不同的蛋白激酶的活性控制,分别是nimXcdc2编码的周期蛋白依赖性激酶p34nimX和nimA编码的NIMA蛋白激酶。p34nimX的激活需要与细胞周期蛋白NIMEcyclinB相结合,其活性在核分裂时显著提高[20]。NIMA促进有丝分裂,同时抑制孢子极性生长,但是在最佳生长条件时孢子进行极化生长的前提是核完成第一轮有丝分裂[21]。Morris[22]发现细胞周期缺陷突变体在43.5℃限制性温度下,其细胞核虽然不能完成第一次有丝分裂,但孢子依旧能进行极化生长。因此,蛋白激酶和环境生长信号共同组成复杂的调控网络,协调孢子极化形态发生与核分裂周期和细胞生长的作用。
丝状真菌通过形成隔膜完成胞质分裂,其中心孔是细胞核等细胞器和细胞质流动的通道[23]。隔膜的形成也依赖于核分裂,且需要将肌动蛋白聚集到隔膜形成的部位。在一个腔室中所有核开始有丝分裂后约10分钟会出现隔膜,随着菌丝的生长,继续形成新的隔膜和菌丝细胞[24]。在A.nidulans野生株中,胞质分离发生在核分裂三轮之后,该过程由Sep家族基因参与协调[25]。sep突变体有两种类型,其中sepB、sepE、sepI或sepJ突变株能够在37℃限制温度下完成三到四轮核分裂,但由于无法形成隔膜所以停止生长。sepA、sepD、sepG和sepH突变株在限制温度下形成隔膜原基,但在隔膜组装后期被阻断。
2.2 核迁移过程的基因调控
丝状真菌多核分生孢子的形成与核迁移有关。核迁移通常发生在营养菌丝生长过程中,多核的产生不是由新形成的分生孢子继续有丝分裂形成的,而是通过细胞核向子代迁移形成[26]。核迁移方式的多样性导致核型的多样,这主要与微管组织中心(MTOC)驱动的微管(MT)蛋白系统相关,包括细胞质微管(CMT)、纺锤体微管(SPB)和星状微管,细胞核的位置取决于MT的位置。微管在有丝分裂时在细胞核内形成纺锤体,在间期形成胞质轨道[27],可直接对细胞核施加拉力和推力或通过与MTOC的连接参与核定位。在多核丝状真菌中,CMT细胞骨架具有控制同一细胞内多个细胞核迁移和分布的能力。此外,核迁移还可以依赖其他细胞器沿MT运动,这些细胞器不是从头合成的,而是在细胞分裂时传给子细胞,必须在胞质分离前被精确定位。在S.cerevisiae中,核定位通过源自纺锤体的细胞质微管进行。由于纺锤体在核膜内组装,细胞核在有丝分裂前必须精确地定位在母芽颈部,以便为母细胞和芽提供遗传物质,然后核内SPB的伸长使得核从母细胞移动到子细胞[28-29]。肌动蛋白在核迁移过程中与微管协同作用,主要通过两种机制在核定位中起作用:肌动球蛋白收缩,或肌动蛋白偶联跨膜肌动蛋白逆行流动。胞质动力蛋白和驱动蛋白是MT的主要马达,可以直接连接MT的核膜或控制MT张力,其功能需要动力蛋白激活蛋白的辅助[30]。Maruyama,J[31]等克隆并鉴定了米曲霉细胞质动力蛋白重链编码基因(dhcA),证实了胞质动力蛋白还在维持米曲霉分生孢子核数目方面起作用。除此之外,my01、act1、c1n、ypt1、spaI、tcp1、dyn1、jnm1、nup1等基因的功能也是调控细胞核运动所必需的。这些基因对应的蛋白质与微管相互作用,有些参与微管的稳定[32]。由此可得,控制细胞核的位置普遍通过主动迁移机制和细胞核正确锚定机制来实现,这对于真菌细胞生长和发育至关重要。
丝状真菌的菌丝通过顶端极化生长形成相互联系的细胞网络,菌丝体中的多个细胞核会彼此保持适当的距离,这需要胞质动力蛋白和动力蛋白复合物以及其他几种蛋白质共同作用,涉及五个相关基因,分别是nudA、nudC、nudE、nudF和nudG[27]。nud突变体的细胞核在42℃限制温度下不能从萌发孢子体移到分生孢子梗中,导致细胞核分布异常,多数聚集在菌丝隔膜附近[33]。nudA编码细胞质动力蛋白的重链,nudC参与核迁移和顶端极性生长,但nudC突变不影响微管结构的完整,也不参与核分裂,它与微管蛋白、肌动蛋白和驱动蛋白之一相互作用或是调节其活性[34]。A.nidulans的nudE编码产物NUDE是N.crassa的核分布蛋白RO11的同源物,在细胞质动力蛋白通路中起作用。nudF[35]基因编码一个包含β-转导蛋白重复序列的保守蛋白,因其与异源三聚体G蛋白亚基序列相似,表明该蛋白与细胞内信号转导有关[26]。另外还在S.cerevisiae和哺乳动物中分别发现了NUDE的同源物PAC1和LIS1参与胞质动力蛋白功能,并且哺乳动物的NUDC同源物也与LIS1蛋白相互作用[36]。这表明nud家族基因共同组成胞质动力蛋白调节通路,并且调节机制是保守的。
核迁移是菌丝顶端发生极化生长的基础,由多种蛋白共同调控。其中一种核定位蛋白APS(包括APSA和APSB)定位于菌丝的细胞质膜[37],Suelmann[38]在A.nidulans构建stu-GFP融合系统,利用荧光显微镜和延时成像追踪细胞核迁移,观察到多核菌丝细胞的一个腔室中,整体上所有核都向菌丝顶端迁移,但每个核的迁移速率不同。这表明存在控制同一细胞质内各个核运动的调节因子,其通过与微管、马达蛋白连接或触发马达蛋白活性来调控核运动,该调节因子即Aps。在Aps缺失株中,分生孢子在初生孢子梗形成之前发育正常。在形成初生孢子梗的过程中,细胞核无法从囊泡迁移到瓶梗,并且不能激活发育特异基因。但细胞核偶尔也会进入初生孢子梗,这些孢子梗就会继续形成产孢瓶梗细胞,从而完成生命周期。这表明细胞核进入初生孢子梗是一个发育节点,确保菌丝的形成。发育缺陷型Aps突变体细胞核未能进入初生孢子梗,发育停止在瓶梗起始阶段。除发育停滞外,许多分生孢子特异性基因在ApsA突变体中没有活性,这表明ApsA基因产物可能直接激活它们,或者ApsA是指导这些基因转录的调控因子。在酿酒酵母中发现了ApsA蛋白的同源物NUM1p,该蛋白在细胞周期的G2阶段定位于酵母母细胞的皮层,并与星状微管相互作用[39]。
在A.nidulans中除了产生单核分生孢子外,在相同的环境条件下还以恒定的比例产生双核和三核分生孢子,该性状由bncA单基因控制[40]。它在异核体中非自主表达且受温度影响,参与细胞核从囊泡向初生孢子梗的迁移过程[41-42]。可见bncA基因与Aps基因调控方式相反。在Aps突变体中,细胞核移动到囊泡后不能继续向初生孢子梗移动,因此发育在瓶梗阶段停滞。而在bncA基因存在下,两个核可以向初生孢子梗移动,这表明双核分生孢子的产生可能是由于两个核结合到初生孢子梗中形成的。在bncA突变体中细胞核进入初生孢子梗就会形成隔膜,因此不会形成1个以上的核。本实验室也通过蛋白质组学等技术在A.oryzae中发现了两个与分生孢子核型相关的候选基因,有关此两基因的功能验证尚在进行中。
3 展望
丝状真菌种类繁多、分布广泛,在基因组、蛋白质组、转录组等各种组学基础上对其功能基因的深入发掘,促进我们了解无性孢子形态结构差异和繁殖方式等各种复杂行为形成的机理,为更好地利用丝状真菌提供理论指导。丝状真菌作为细胞工厂,广泛用于生产各种抗生素、有机酸、酶和维生素等,新技术诸如CRISPR等在丝状真菌基因组中的应用可改善其多核特性带来的不良效应,加快丝状真菌遗传改良效率[47-48],构建高效遗传操作体系,更好地为人类服务。同时,CRISPR技术在丝状真菌中的应用仍需进一步完善,如其适用性、可扩展性和靶向效率方面仍有待改进。
在利用丝状真菌为人类创造财富的同时,也不应忽视一些丝状真菌物种对动植物的潜在威胁。如A.fumigatus,A.flavus和Talaromycesmarneffei等已经被世界卫生组织列为重要的病原微生物[49-50]。了解分生孢子繁殖方式的机制有助于制定出有效的病害诊断措施和特异性抗真菌药物。CRISPR-Cas9的应用成功鉴别T.marneffei致病性的关键基因[51],从遗传水平阐明致病真菌的发病机制,并为分子抗真菌治疗提供靶点。未来,在深入挖掘丝状真菌各类组学信息和充分利用分子生物学技术基础上,改善真菌疾病的检测、预防和治疗方法,并为开发利用丝状真菌资源发挥积极作用。
猜你喜欢
首都食品与医药(2023年14期)2023-07-17
黑龙江大学自然科学学报(2022年4期)2022-11-17
生物化工(2022年4期)2022-09-20
——水芹主要害虫识别与为害症状
长江蔬菜(2022年13期)2022-07-29
医药前沿(2018年13期)2018-04-20
中外医疗(2016年15期)2016-12-01
中外医疗(2015年5期)2016-01-04
化工进展(2015年6期)2015-11-13
恋爱婚姻家庭·养生版(2015年10期)2015-05-14
化工管理(2015年6期)2015-03-23
